

Organic Acid Accumulation and Associated Dynamic Changes in Enzyme Activity and Gene Expression during Fruit Development and Ripening of Common Loquat and Its Interspecific Hybrid
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sampling
2.2. Chemicals and Reagents
2.3. Fruit Weight, Shape, and Color Measurement
2.4. Determination of Titratable Acidity (TA)
2.5. Determination of OAs Using HPLC Coupled with Ultraviolet (UV) Detection
2.6. Assay of OA Metabolism-Related Enzyme Activity
2.7. Quantification of Gene Expression Using Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.8. Statistical Analyses
3. Results
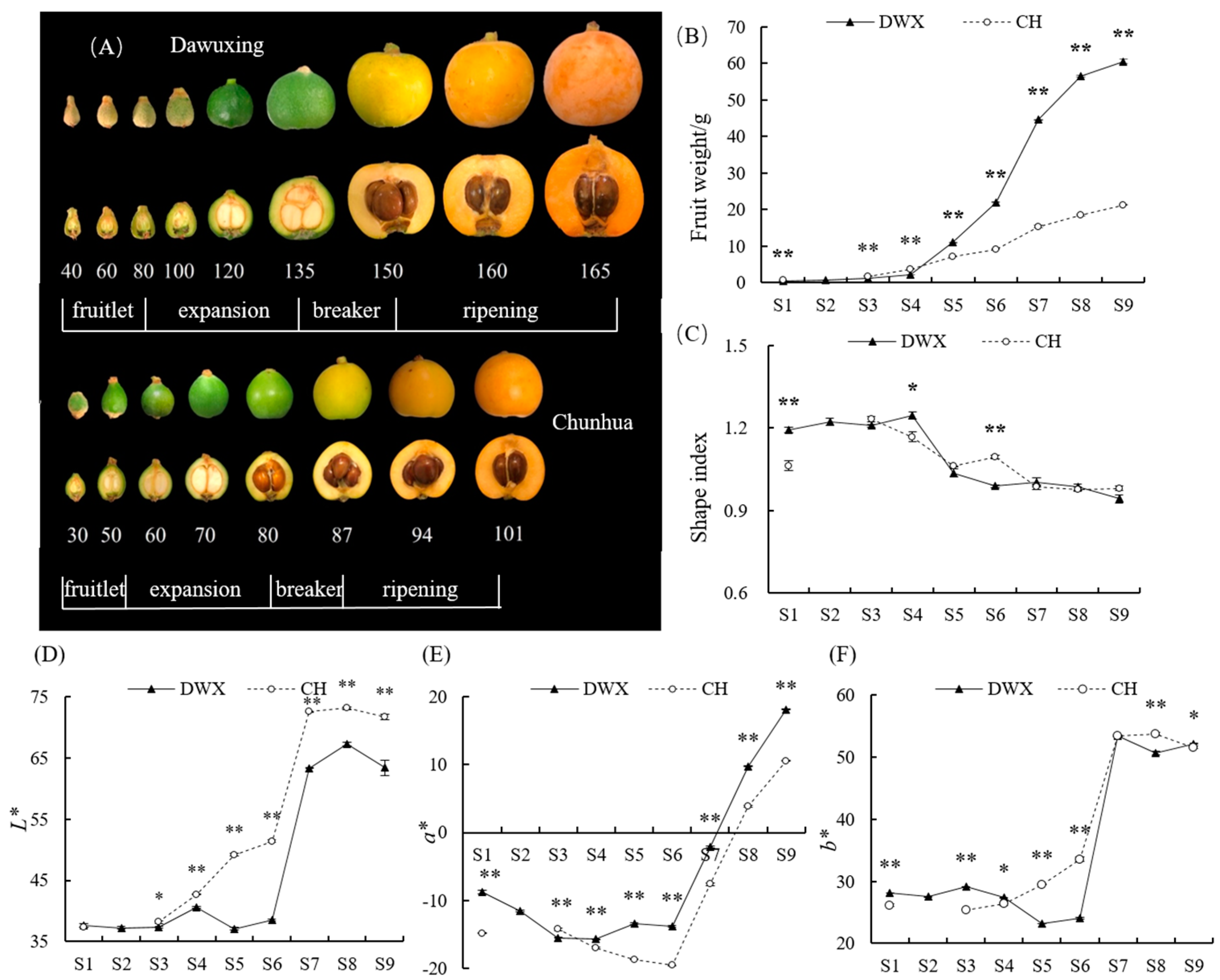
3.1. Comparative Analyses of Fruit Phenotype, Weight, Shape, and Color of Dawuxing Loquat and Its Interspecific Hybrid
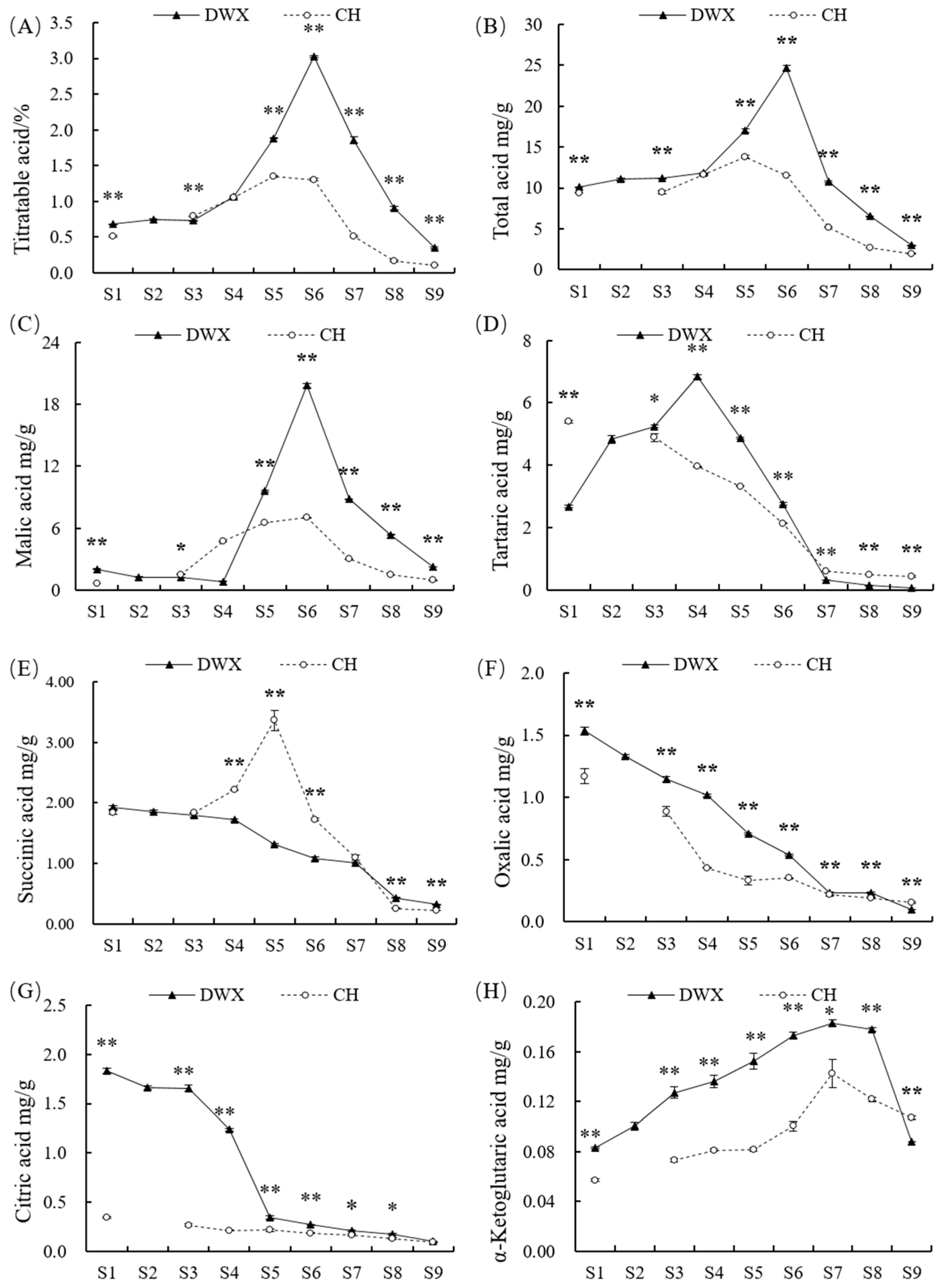
3.2. Comparative Analyses of Organic Acid Compositions and Contents of Dawuxing Loquat and Its Interspecific Hybrid
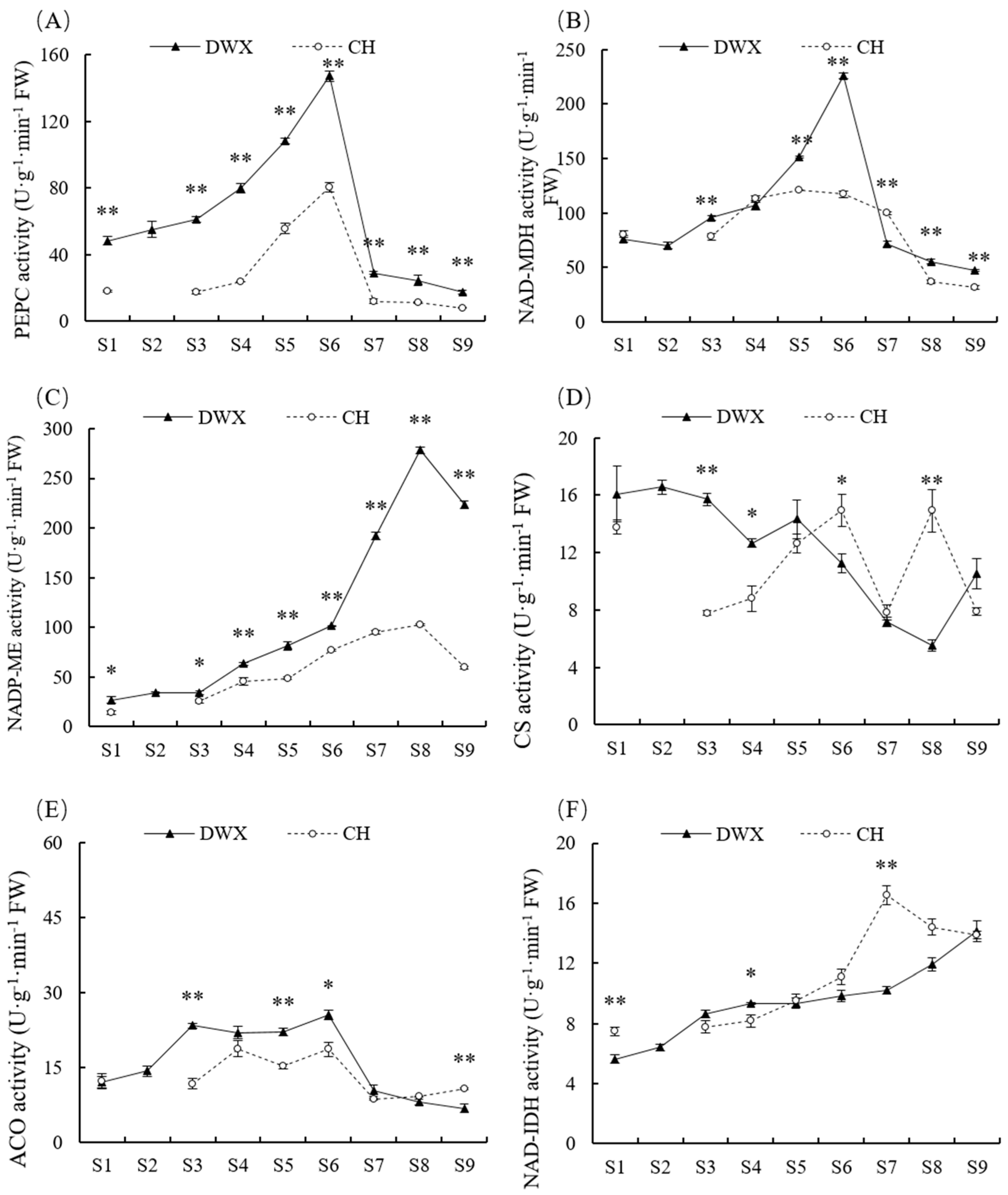
3.3. Comparative Analyses of Enzyme Activity for Organic Acid Metabolism of Dawuxing Loquat and Its Interspecific Hybrid
3.4. Comparative Analyses of Gene Expression for Organic Acid Metabolism of Dawuxing Loquat and Its Interspecific Hybrid
4. Discussion
4.1. Characteristics of Fruit Growth and Development in Dawuxing Loquat and Its Interspecific Hybrid
4.2. Organic Acid Accumulation in Dawuxing Loquat and Its Interspecific Hybrid
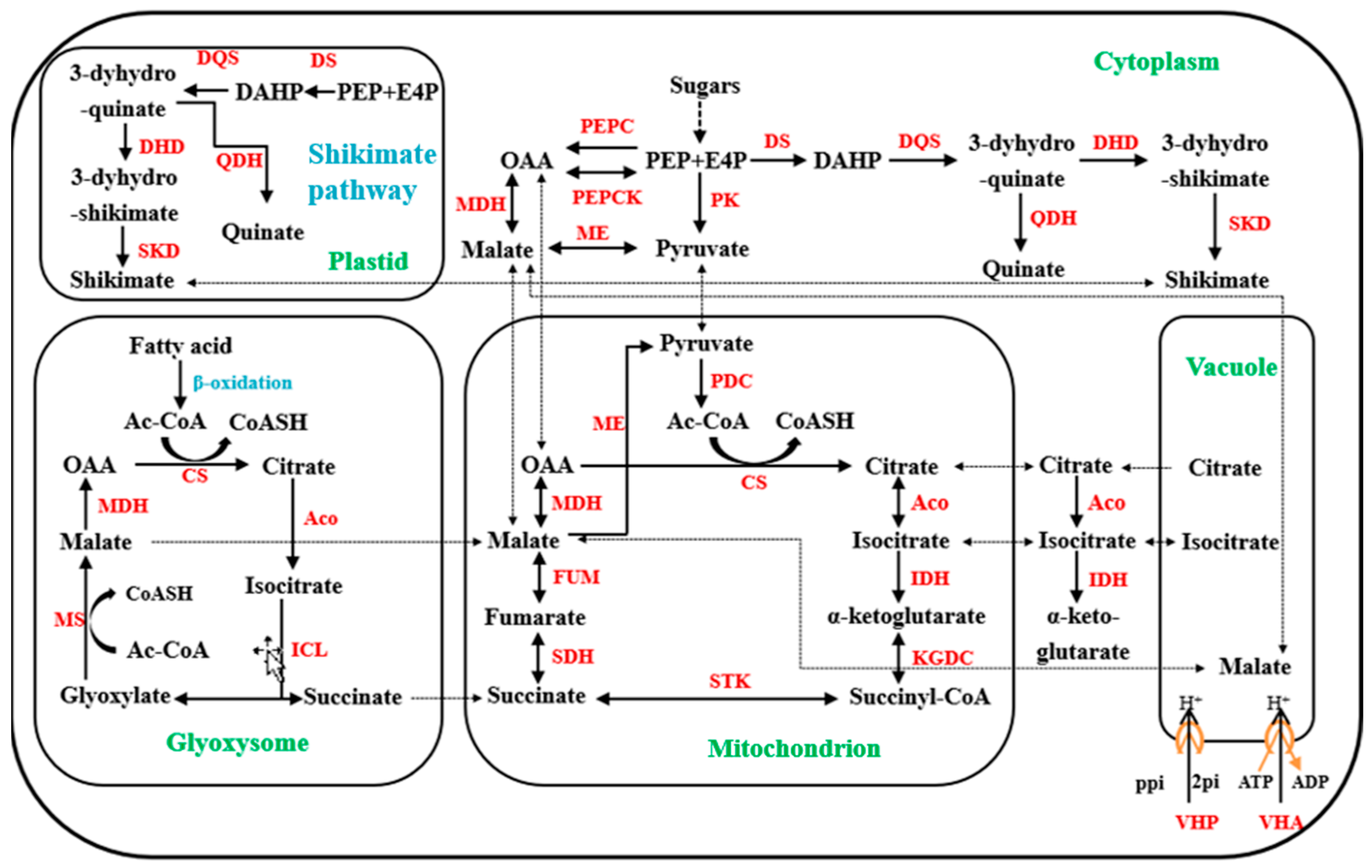
4.3. Organic Acid Synthesis, Degradation, and Transport-Related Enzyme Activity and Gene Expression in Dawuxing Loquat and Its Interspecific Hybrid
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Badenes, M.L.; Janick, J.; Zhang, Z.; Liang, G.; Wu, W. Breeding loquat. Plant Breed. Rev. 2013, 37, 259–296. [Google Scholar]
- Huang, X.; Wang, H.; Qu, S.; Luo, W.; Gao, Z. Using artificial neural network in predicting the key fruit quality of loquat. Food Sci. Nutr. 2021, 9, 1780–1791. [Google Scholar] [CrossRef]
- Zou, S.; Wu, J.; Shahid, M.Q.; He, Y.; Lin, S.; Liu, Z.; Yang, X. Identification of key taste components in loquat using widely targeted metabolomics. Food Chem. 2020, 323, 126822. [Google Scholar] [CrossRef]
- Costa, B.P.; Ikeda, M.; de Melo, A.M.; Bambirra Alves, F.E.S.; Carpiné, D.; Ribani, R.H. Eriobotrya japonica fruits and its by-products: A promising fruit with bioactive profile and trends in the food application—A bibliometric review. Food Biosci. 2022, 50, 102099. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, W.; Xu, C.; Li, X. Biological activities of extracts from loquat (Eriobotrya japonica Lindl.): A review. Int. J. Mol. Sci. 2016, 17, 1983. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.X.; Jian, T.Y.; Lv, H.; Ding, X.Q.; Zuo, Y.Y.; Ren, B.R.; Chen, J.; Li, W.L. Antitussive and expectorant properties of growing and fallen leaves of loquat (Eriobotrya japonica). Rev. Bras. Farmacogn. 2018, 28, 239–242. [Google Scholar] [CrossRef]
- Wang, Y. A draft genome, resequencing, and metabolomes reveal the genetic background and molecular basis of the nutritional and medicinal properties of loquat (Eriobotrya japonica (Thunb.) Lindl). Hortic. Res. 2021, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Yuan, Q.; Lin, S.; Liu, W.; Du, G.; Zhao, L.; Zhang, Q.; Lin, D.R.; Liu, Y.T.; Qin, W.; et al. Physicochemical characteristics and biological activities of polysaccharides from the leaves of different loquat (Eriobotrya japonica) cultivars. Int. J. Biol. Macromol. 2019, 135, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Du, K.; Deng, Q.; Zeng, J.; Kang, W.; Xu, Y.; Zhang, J. Identification of interspecific hybrids between loquat (Eriobotrya japonica Lindl.) and bengal loquat (E. bengalensis Hook.). Pakistan J. Bot. 2017, 49, 897–902. [Google Scholar]
- Chi, Z.H.; Wang, Y.Q.; Deng, Q.X.; Zhang, H.; Pan, C.P.; Yang, Z.W. Endogenous phytohormones and the expression of flowering genes synergistically induce flowering in loquat. J. Integr. Agric. 2020, 19, 2247–2256. [Google Scholar] [CrossRef]
- Yang, Z.; Deng, Q.; Wang, Y.; Du, K.; Pan, C.; Zhang, H.; Zhang, H.; Wang, Y. A new spring flowering loquat cultivar “Chunhua 1”. Acta Hortic. Sin. 2021, 48, 2819–2820. [Google Scholar]
- Batista-Silva, W.; Nascimento, V.L.; Medeiros, D.B.; Nunes-Nesi, A.; Ribeiro, D.M.; Zsögön, A.; Araújo, W.L. Modifications in organic acid profiles during fruit development and ripening: Correlation or causation? Front. Plant Sci. 2018, 9, 1689. [Google Scholar] [CrossRef] [PubMed]
- Etienne, A.; Génard, M.; Lobit, P.; Mbeguié-A-Mbéguié, D.; Bugaud, C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, R.P.; Famiani, F. Organic acids in fruits: Metabolism, functions and contents. Hortic. Rev. 2018, 45, 371–430. [Google Scholar]
- Huang, X.Y.; Wang, C.K.; Zhao, Y.W.; Sun, C.H.; Hu, D.G. Mechanisms and regulation of organic acid accumulation in plant vacuoles. Hortic. Res. 2021, 8, 227. [Google Scholar] [CrossRef]
- Emmerlich, V.; Linka, N.; Reinhold, T.; Hurth, M.A.; Traub, M.; Martinoia, E.; Neuhaus, H.E. The plant homolog to the human sodium/dicarboxylic cotransporter is the vacuolar malate carrier. Proc. Natl. Acad. Sci. USA 2003, 100, 11122–11126. [Google Scholar] [CrossRef] [Green Version]
- Kovermann, P.; Meyer, S.; Hörtensteiner, S.; Picco, C.; Scholz-Starke, J.; Ravera, S.; Lee, Y.; Martinoia, E. The Arabidopsis vacuolar malate channel is a member of the ALMT family. Plant J. 2007, 52, 1169–1180. [Google Scholar] [CrossRef]
- Martinoia, E. Vacuolar transporters—Companions on a longtime journey. Plant Physiol. 2018, 176, 1384–1407. [Google Scholar] [CrossRef]
- Fu, X.; Kong, W.; Peng, G.; Zhou, J.; Azam, M.; Xu, C.; Grierson, D.; Chen, K. Plastid structure and carotenogenic gene expression in red- and white-fleshed loquat (Eriobotrya japonica) fruits. J. Exp. Bot. 2012, 63, 341–354. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.M.; Anwar, R.; Naveed, R.; Rehman, U.; Ejaz, S.; Ali, S.; Yousef, A.F.; Ercisli, S.; Hu, X.; Hou, Y.; et al. Sugar and acid profile of loquat (Eriobotrya japonica Lindl.), enzymes assay and expression profiling of their metabolism- related genes as in fluenced by exogenously applied boron. Front. Plant Sci. 2022, 13, 1039360. [Google Scholar] [CrossRef]
- Chen, F.X.; Liu, X.H.; Chen, L.S. Developmental changes in pulp organic acid concentration and activities of acid-metabolising enzymes during the fruit development of two loquat (Eriobotrya japonica Lindl.) cultivars differing in fruit acidity. Food Chem. 2009, 114, 657–664. [Google Scholar] [CrossRef]
- Su, W.; Jing, Y.; Lin, S.; Yue, Z.; Yang, X.; Xu, J.; Wu, J.; Zhang, Z.; Xia, R.; Zhu, J.; et al. Polyploidy underlies co-option and diversification of biosynthetic triterpene pathways in the apple tribe. Proc. Natl. Acad. Sci. USA 2021, 118, e2101767118. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Niu, X.Q.; Zheng, X.L.; Chen, X.; Zheng, G.H.; Wu, J.C. Comparative transcriptome analysis reveals key genes potentially related to organic acid and sugar accumulation in loquat. PLoS ONE 2021, 16, e0238873. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Alam, S.M.; Anwar, R.; Ali, S.; Shi, M.; Liang, D.; Lin, Z.; Chen, F. Genome-wide identification, characterization and expression profiling of aluminum-activated malate transporters in Eriobotrya japonica Lindl. Horticulturae 2021, 7, 441. [Google Scholar] [CrossRef]
- Cuevas, J.; Salvador-Sola, F.J.; Gavilán, J.; Lorente, N.; Hueso, J.J.; González-Padierna, C.M. Loquat fruit sink strength and growth pattern. Sci. Hortic. 2003, 98, 131–137. [Google Scholar] [CrossRef]
- Hu, D.G.; Li, Y.Y.; Zhang, Q.Y.; Li, M.; Sun, C.H.; Yu, J.Q.; Hao, Y.J. The R2R3-MYB transcription factor MdMYB73 is involved in malate accumulation and vacuolar acidification in apple. Plant J. 2017, 91, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Jia, D.; Shen, F.; Wang, Y.; Wu, T.; Xu, X.; Zhang, X.; Han, Z. Apple fruit acidity is genetically diversified by natural variations in three hierarchical epistatic genes: Mdsaur37, Mdpp2ch and Mdalmtii. Plant J. 2018, 95, 427–443. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Zhao, L.; Jiang, X.; Cherono, S.; Liu, J.J.; Ogutu, C.; Ntini, C.; Zhang, X.; Han, Y. Assessment of organic acid accumulation and its related genes in peach. Food Chem. 2021, 334, 127567. [Google Scholar] [CrossRef]
- Hussain, S.B.; Shi, C.Y.; Guo, L.X.; Kamran, H.M.; Sadka, A.; Liu, Y.Z. Recent advances in the regulation of citric acid metabolism in citrus fruit. CRC Crit. Rev. Plant Sci. 2017, 36, 241–256. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, X.; Ali, M.M.; Rizwan, H.M.; Li, B.; Li, H.; Jia, K.; Yang, X.; Ma, S.; Li, S.; et al. Changes in the content of organic acids and expression analysis of citric acid accumulation-related genes during fruit development of yellow (Passiflora edulis f. flavicarpa) and purple (Passiflora edulis f. edulis) passion fruits. Int. J. Mol. Sci. 2021, 22, 5767. [Google Scholar] [CrossRef]
- Chen, M.; Jiang, Q.; Yin, X.R.; Lin, Q.; Chen, J.Y.; Allan, A.C.; Xu, C.J.; Chen, K.S. Effect of hot air treatment on organic acid- and sugar-metabolism in ponkan (Citrus reticulata) fruit. Sci. Hortic. 2012, 147, 118–125. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Sun, J.; Jackson, A. Dynamic changes of enzymes involved in sugar and organic acid level modification during blueberry fruit maturation. Food Chem. 2020, 309, 125617. [Google Scholar] [CrossRef]
- Putzeys, P.; Reijnaers, H. Haemoglobin association and the sigmoid oxygen equilibrium curve. Nature 1966, 212, 506. [Google Scholar] [CrossRef] [PubMed]
- Notton, B.A.; Blanke, M.M. Phosphoenolpyruvate carboxylase in avocado fruit: Purification and properties. Phytochemistry 1993, 33, 1333–1337. [Google Scholar] [CrossRef]
- Jiang, Z.; Huang, Q.; Jia, D.; Zhong, M.; Tao, J.; Liao, G.; Huang, C.; Xu, X. Characterization of organic acid metabolism and expression of related genes during fruit development of Actinidia eriantha ‘Ganmi 6’. Plants 2020, 9, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, M.; Bie, Z.L.; Wu, M.Z.; Yi, H.P.; Feng, J.X. Changes in organic acids and acid metabolism enzymes in melon fruit during development. Sci. Hortic. 2010, 123, 360–365. [Google Scholar] [CrossRef]
- Mindell, J.A. Lysosomal acidification mechanisms. Annu. Rev. Physiol. 2012, 74, 69–86. [Google Scholar] [CrossRef] [Green Version]
- Sharma, T.; Dreyer, I.; Kochian, L.; Piñeros, M.A. The ALMT family of organic acid transporters in plants and their involvement in detoxification and nutrient security. Front. Plant Sci. 2016, 7, 1488. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage a | Oxalic Acid | Tartaric Acid | Malic Acid | α-Ketoglutaric Acid | Citric Acid | Succinic Acid | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DWX | CH | DWX | CH | DWX | CH | DWX | CH | DWX | CH | DWX | CH | |
| S1 | 15.21 | 12.43 | 26.52 | 57.37 | 20.23 | 6.41 | 0.82 | 0.61 | 18.17 | 3.64 | 19.05 | 19.55 |
| S2 | 12.06 | 43.64 | 11.52 | 0.91 | 15.06 | 16.81 | ||||||
| S3 | 10.28 | 9.37 | 46.91 | 51.58 | 10.83 | 16.08 | 1.14 | 0.77 | 14.83 | 2.80 | 16.01 | 19.41 |
| S4 | 8.61 | 3.75 | 58.00 | 34.06 | 7.17 | 40.60 | 1.15 | 0.70 | 10.50 | 1.81 | 14.57 | 19.07 |
| S5 | 4.16 | 2.40 | 28.64 | 24.01 | 56.57 | 47.16 | 0.89 | 0.59 | 2.02 | 1.55 | 7.70 | 24.28 |
| S6 | 2.19 | 3.05 | 11.17 | 18.54 | 80.48 | 60.98 | 0.70 | 0.90 | 1.09 | 1.60 | 4.37 | 14.93 |
| S7 | 2.20 | 4.16 | 2.93 | 11.36 | 81.92 | 57.64 | 1.70 | 2.58 | 1.96 | 3.16 | 9.29 | 21.09 |
| S8 | 3.57 | 7.17 | 2.02 | 17.97 | 82.48 | 56.03 | 2.73 | 4.55 | 2.61 | 4.76 | 6.59 | 9.52 |
| S9 | 3.33 | 8.13 | 1.99 | 21.79 | 77.51 | 48.59 | 2.99 | 5.75 | 3.42 | 4.52 | 10.76 | 11.22 |
| Index | Titratable Acid | Total Acid | ||
|---|---|---|---|---|
| Dawuxing | Chunhua | Dawuxing | Chunhua | |
| titratable acid | 1 | 1 | 0.883 ** | 0.940 ** |
| oxalic acid | −0.269 | 0.093 | 0.160 | 0.383 |
| tartaric acid | 0.005 | 0.457 * | 0.390 * | 0.714 ** |
| malic acid | 0.948 ** | 0.886 ** | 0.789 ** | 0.704 ** |
| α-ketoglutaric acid | 0.703 ** | −0.366 | 0.437 * | −0.612 ** |
| critic acid | −0.456 * | 0.348 | −0.054 | 0.616 ** |
| succinic acid | −0.087 | 0.866 ** | 0.301 | 0.950 ** |
| Index | PEPC | NAD-MDH | NADP-ME | CS | ACO | NAD-IDH | |
|---|---|---|---|---|---|---|---|
| malic acid | DWX | 0.676 ** | 0.808 ** | 0.167 | −0.308 | 0.369 | 0.169 |
| CH | 0.570 ** | 0.756 ** | 0.251 | −0.247 | 0.224 | 0.793 ** | |
| α-ketoglutaric acid | DWX | 0.298 | 0.405 * | 0.423 * | −0.642 ** | 0.228 | 0.287 |
| CH | −0.177 | −0.225 | 0.908 ** | −0.105 | −0.420 * | 0.907 ** | |
| citric acid | DWX | −0.020 | −0.176 | −0.817 ** | 0.751 ** | 0.234 | −0.805 ** |
| CH | 0.098 | 0.414 * | −0.770 ** | 0.159 | 0.200 | −0.766 ** | |
| total acid | DWX | 0.947 ** | 0.960 ** | −0.435 * | 0.200 | 0.794 ** | −0.314 |
| CH | 0.693 ** | 0.872 ** | −0.505 ** | 0.202 | 0.730 ** | −0.731 ** | |
| Genes | Malic Acid | α-Ketoglutaric Acid | Citric Acid | Total Acid | ||||
|---|---|---|---|---|---|---|---|---|
| DWX | CH | DWX | CH | DWX | CH | DWX | CH | |
| PEPC | 0.865 ** | 0.753 ** | 0.532 ** | −0.019 | −0.567 ** | 0.178 | 0.647 ** | 0.657 ** |
| PEPC2 | 0.275 | 0.198 | −0.189 | −0.685 ** | 0.478 * | 0.624 ** | 0.726 ** | 0.692 ** |
| NAD-MDH | 0.792 ** | 0.578 ** | 0.545 ** | −0.453 * | −0.262 | 0.499 * | 0.707 ** | 0.820 ** |
| NADP-ME | 0.235 | −0.215 | 0.568 ** | −0.536 ** | −0.540 ** | 0.458 * | −0.002 | 0.283 |
| NADP-ME2 | −0.186 | −0.495 * | −0.303 | 0.664 ** | −0.486 * | −0.742 ** | −0.623 ** | −0.925 ** |
| NADP-ME4 | 0.376 | −0.176 | 0.765 ** | −0.205 | −0.637 ** | −0.046 | 0.109 | −0.051 |
| CS | −0.271 | 0.769 ** | −0.126 | −0.315 | 0.675 ** | 0.199 | 0.152 | 0.801 ** |
| ATP-CS α1 | 0.711 ** | 0.557 ** | 0.355 | −0.194 | −0.229 | 0.160 | 0.873 ** | 0.497 * |
| ATP-CS α2-1 | 0.477 * | −0.030 | 0.825 ** | 0.321 | −0.468 * | −0.189 | 0.187 | −0.219 |
| ATP-CS β2 | 0.388 * | 0.442 * | 0.322 | −0.208 | −0.055 | 0.199 | 0.591 ** | 0.402 |
| ACO | 0.300 | 0.457 * | 0.675 ** | −0.767 ** | −0.596 ** | 0.616 ** | −0.097 | 0.858 ** |
| NAD-IDH1 | 0.707 ** | −0.053 | 0.874 ** | −0.380 | −0.547 ** | 0.421 * | 0.490 ** | 0.299 |
| NAD-IDH5 | 0.436 * | 0.228 | 0.793 ** | 0.360 | −0.454 * | −0.027 | 0.124 | 0.070 |
| tDT2 | 0.014 | −0.383 | −0.259 | 0.616 ** | 0.394 * | −0.250 | 0.108 | −0.601 ** |
| VHA-A | 0.188 | 0.094 | 0.520 ** | 0.021 | 0.019 | 0.201 | 0.371 | 0.186 |
| VHA-A3 | 0.823 ** | −0.556 ** | 0.624 ** | −0.515 * | −0.380 | 0.628 ** | 0.681 ** | −0.012 |
| VHA-B2-1 | 0.501 ** | 0.226 | 0.843 ** | −0.080 | −0.510 ** | 0.171 | 0.240 | 0.338 |
| VHA-B2-2 | −0.420 * | 0.119 | −0.480 * | 0.301 | −0.035 | −0.224 | −0.556 ** | −0.119 |
| VHA-C | −0.073 | −0.104 | −0.138 | −0.630 ** | 0.633 ** | 0.766 ** | 0.456 * | 0.462 * |
| VHA-D2 | 0.491 ** | −0.242 | 0.766 ** | −0.364 | −0.435 * | −0.043 | 0.194 | −0.061 |
| VHA-E1 | −0.288 | −0.275 | 0.323 | −0.171 | 0.163 | 0.369 | −0.334 | 0.108 |
| VHA-F | −0.306 | −0.451 * | −0.103 | −0.498 * | 0.618 ** | 0.569 ** | 0.174 | 0.104 |
| VHP1 | 0.259 | −0.584 ** | 0.315 | 0.254 | −0.815 ** | −0.181 | −0.285 | −0.633 ** |
| ALMT1 | 0.159 | 0.027 | 0.175 | −0.824 ** | 0.148 | 0.778 ** | 0.571 ** | 0.673 ** |
| ALMT2 | −0.483 * | −0.347 | −0.097 | −0.286 | 0.574 ** | 0.123 | −0.137 | −0.051 |
| ALMT3 | −0.150 | −0.473 * | 0.295 | 0.472 * | −0.375 | −0.486 * | −0.567 ** | −0.735 ** |
| ALMT4 | −0.588 ** | −0.251 | −0.477 * | −0.348 | 0.748 ** | 0.367 | −0.140 | 0.165 |
| ALMT5 | 0.056 | −0.242 | −0.374 | −0.366 | 0.509 ** | 0.383 | 0.298 | 0.167 |
| ALMT6 | 0.183 | −0.800 ** | 0.081 | −0.127 | 0.011 | 0.261 | 0.019 | −0.460 * |
| ALMT7 | 0.444 * | 0.352 | 0.572 ** | −0.216 | −0.715 ** | 0.120 | 0.096 | 0.473 * |
| ALMT8 | 0.700 ** | −0.405 * | 0.855 ** | −0.001 | −0.652 ** | −0.466 * | 0.338 | −0.513 * |
| ALMT9 | −0.078 | −0.489 * | 0.283 | −0.683 ** | 0.233 | 0.818 ** | 0.121 | 0.208 |
| ALMT10 | 0.580 ** | −0.345 | 0.089 | −0.736 ** | 0.233 | 0.896 ** | 0.771 ** | 0.335 |
| ALMT11 | 0.003 | −0.469 * | 0.471 * | 0.745 ** | −0.533 ** | −0.775 ** | −0.474 * | −0.940 ** |
| ALMT12 | −0.070 | 0.072 | 0.117 | −0.830 ** | 0.605 ** | 0.786 ** | 0.416 * | 0.684 ** |
| ALMT13 | −0.193 | −0.220 | −0.032 | −0.805 ** | 0.573 ** | 0.827 ** | 0.223 | 0.481 * |
| ALMT14 | 0.405 * | −0.088 | 0.068 | −0.857 ** | 0.331 | 0.873 ** | 0.619 ** | 0.631 ** |
| ALMT15 | 0.401 * | −0.243 | 0.806 ** | −0.332 | −0.398 * | 0.322 | 0.138 | 0.130 |
| ALMT16 | 0.338 | −0.398 | −0.131 | −0.686 ** | 0.267 | 0.847 ** | 0.623 ** | 0.245 |
| ALMT17 | −0.351 | −0.579 ** | −0.654 ** | 0.429 * | 0.493 ** | −0.423 * | −0.233 | −0.775 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, H.; Li, X.; Wang, Y.; Ma, Q.; Zeng, Y.; Xiang, Y.; Chen, M.; Zhang, H.; Xia, H.; Liang, D.; et al. Organic Acid Accumulation and Associated Dynamic Changes in Enzyme Activity and Gene Expression during Fruit Development and Ripening of Common Loquat and Its Interspecific Hybrid. Foods 2023, 12, 911. https://doi.org/10.3390/foods12050911
Deng H, Li X, Wang Y, Ma Q, Zeng Y, Xiang Y, Chen M, Zhang H, Xia H, Liang D, et al. Organic Acid Accumulation and Associated Dynamic Changes in Enzyme Activity and Gene Expression during Fruit Development and Ripening of Common Loquat and Its Interspecific Hybrid. Foods. 2023; 12(5):911. https://doi.org/10.3390/foods12050911
Chicago/Turabian StyleDeng, Honghong, Xuelian Li, Yang Wang, Qiaoli Ma, Yuge Zeng, Yinchun Xiang, Mingmin Chen, Huifen Zhang, Hui Xia, Dong Liang, and et al. 2023. "Organic Acid Accumulation and Associated Dynamic Changes in Enzyme Activity and Gene Expression during Fruit Development and Ripening of Common Loquat and Its Interspecific Hybrid" Foods 12, no. 5: 911. https://doi.org/10.3390/foods12050911