
Peptidomics Study of Plant-Based Meat Analogs as a Source of Bioactive Peptides

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Beef and PBMA Gastrointestinal Digests
2.3. Molecular Weight Distribution
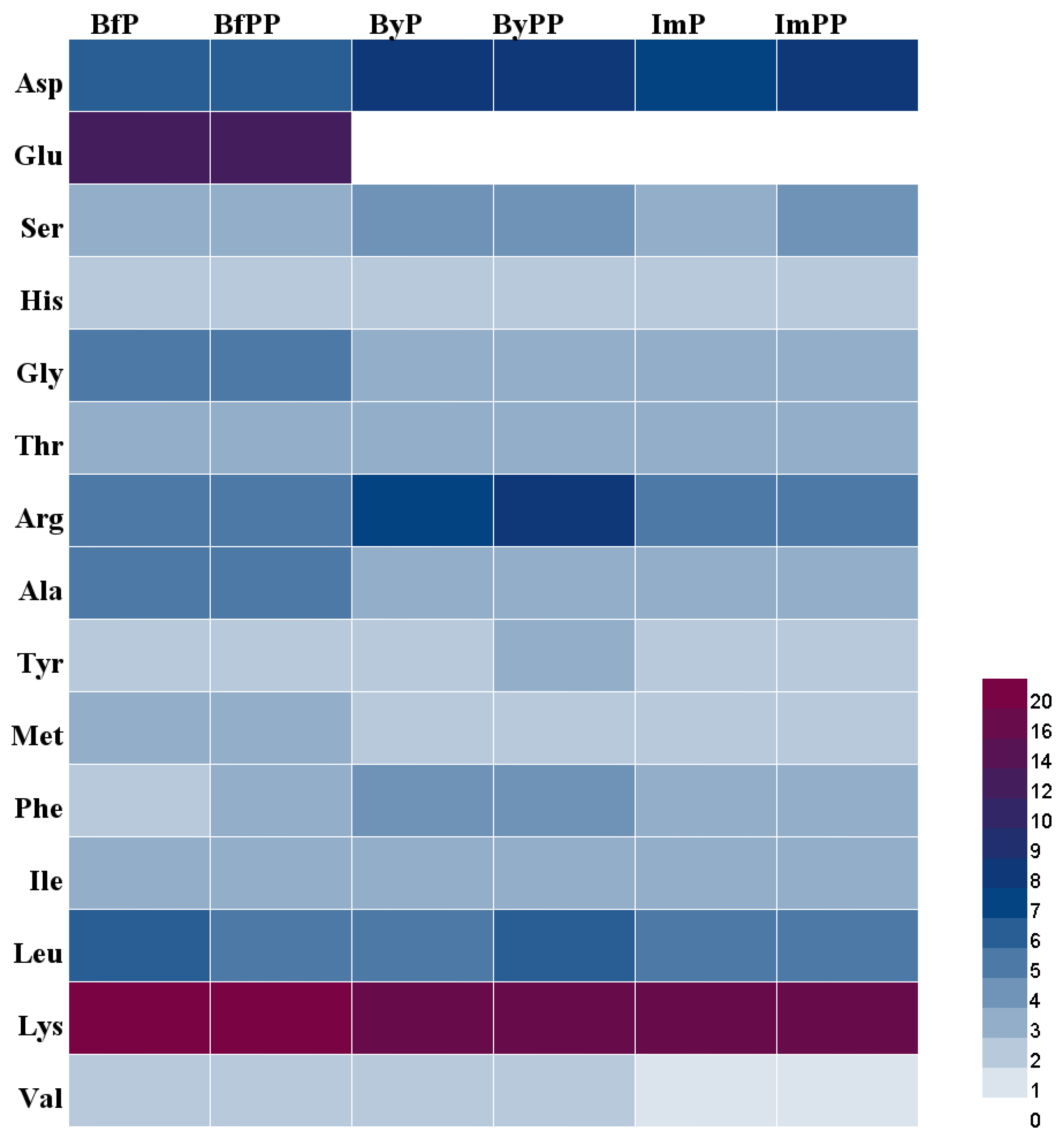
2.4. Degree of Hydrolysis (DH) and Amino Acid Compositions
2.5. Identification of Peptides by LC-MS/MS
2.6. ACE Inhibition Assay
2.7. Desalting Protocol, Cell Culture and Cytotoxicity
2.8. Superoxide Detection
2.9. Western Blotting
2.10. Statistical Analysis
3. Results and Discussion
3.1. Molecular Weight Distribution, DH and Amino Acid Compositions of Beef and PBMA Digests
3.2. Effects of Gastrointestinal Digestion on Peptide Profiles of Beef and PBMA Hydrolysates
3.3. ACE Inhibition, Antioxidant and Anti-Inflammation of Beef and PBMA Hydrolysates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, S.; Zhao, M.; Fan, H.; Wu, J. Emerging proteins as precursors of bioactive peptides/hydrolysates with health benefits. Curr. Opin. Food Sci. 2022, 48, 100914. [Google Scholar] [CrossRef]
- Hadi, J.; Brightwell, G. The safety of alternative proteins: Technological, environmental and regulatory aspects of cultured meat, plant-based meat, insect protein and single-cell protein. Foods 2021, 10, 1226. [Google Scholar] [CrossRef]
- Yang, H.; Shen, Y.; Li, Y. Physicochemical and functional properties of texturized vegetable proteins and cooked patty textures: Comprehensive characterization and correlation analysis. Foods 2022, 11, 2619. [Google Scholar]
- Onwezen, M.C.; Bouwman, E.P.; Reinders, M.J.; Dagevos, H. A systematic review on consumer acceptance of alternative proteins: Pulses, algae, insects, plant-based meat alternatives, and cultured meat. Appetite 2021, 159, 105058. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; You, J.; Moon, J.; Jeong, J. Mechanisms factors affecting consumers’ alternative meats buying intentions: Plant-based meat alternative and cultured meat. Sustainability 2020, 12, 5662. [Google Scholar] [CrossRef]
- Bohrer, B.M. An investigation of the formulation and nutritional composition of modern meat analogue products. Food Sci. Hum. Wellness 2019, 8, 320–329. [Google Scholar] [CrossRef]
- Singh, M.; Trivedi, N.; Enamala, M.K. Plant-based meat analogue (PBMA) as a sustainable food: A concise review. Eur. Food Res. Technol. 2021, 247, 2499–2526. [Google Scholar] [CrossRef]
- Zhou, H.; Hu, Y.; Tan, Y. Digestibility and gastrointestinal fate of meat versus plant-based meat analogs: An in vitro comparison. Food Chem. 2021, 364, 130439. [Google Scholar] [CrossRef]
- McClements, D. Future foods: Is it possible to design a healthier and more sustainable food supply. Nutr. Bull. 2020, 45, 341–354. [Google Scholar] [CrossRef]
- Sridhar, K.; Bouhallab, S.; Croguennec, T.; Renard, D.; Lechevalier, V. Recent trends in design of healthier plant-based alternatives: Nutritional profile, gastrointestinal digestion, and consumer perception. Crit. Rev. Food Sci. 2022, 2081666. [Google Scholar] [CrossRef]
- Zhang, T.; Dou, W.; Zhang, X. The development history and recent updates on soy protein-based meat alternatives. Trends Food Sci. Technol. 2021, 109, 702–710. [Google Scholar] [CrossRef]
- Khalesi, M.; FitzGerald, R.J. In vitro digestibility and antioxidant activity of plant protein isolate and milk protein concentrate blends. Catalysts 2021, 11, 787. [Google Scholar] [CrossRef]
- Xie, Y.; Cai, L.; Zhao, D. Real meat and plant-based meat analogues have different in vitro protein digestibility properties. Food Chem. 2022, 387, 132917. [Google Scholar] [CrossRef]
- Chen, D.; Rocha-Mendoza, D.; Shan, S. Characterization and cellular uptake of peptides derived from in vitro digestion of meat analogues produced by a sustainable extrusion process. J. Agric. Food Chem. 2022, 70, 8124–8133. [Google Scholar] [CrossRef] [PubMed]
- Mora, L.; Escudero, E.; Toldra, F. Characterization of the peptide profile in Spanish Teruel, Italian Parma and Belgian dry-cured hams and its potential bioactivity. Food Res. Int. 2016, 89, 638–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.; Wang, J.; Liao, W.; Wu, J. Identification and characterization of gastrointestinal-resistant angiotensin-converting enzyme inhibitory peptides from egg white proteins. J. Agric. Food Chem. 2019, 67, 7147–7156. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Fan, H.; Yu, W.; Liao, W.; Wu, J. Spent hen protein hydrolysate with good gastrointestinal stability and permeability in Caco-2 cells shows antihypertensive activity in SHR. Foods 2020, 9, 1384. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Suarez, L.M. Optimization of enzymatic hydrolysis for preparing cassava leaf hydrolysate with antioxidant activity. Food Bioprocess. Tech. 2021, 14, 2181–2194. [Google Scholar] [CrossRef]
- Universal Protein Knowledgebase. 2022. Available online: https://www.uniprot.org/downloads (accessed on 1 May 2022).
- Wu, J.; Aluko, R.; Muir, A. Improved method for direct high-performance liquid chromatography assay of angiotensin-converting enzyme-catalyzed reactions. J. Chromatogr. A 2002, 950, 125–130. [Google Scholar] [CrossRef]
- Wang, X.; Bhullar, K.S.; Fan, H.; Wu, J. Regulatory effects of a pea-derived peptide Leu-Arg-Trp (LRW) on dysfunction of rat aortic vascular smooth muscle cells against angiotensin II stimulation. J. Agric. Food Chem. 2020, 68, 3947–3953. [Google Scholar] [CrossRef]
- Bakhsh, A. A novel approach for tuning the physicochemical, textural, and sensory characteristics of plant-based meat analogs with different levels of methylcellulose concentration. Foods 2021, 10, 560. [Google Scholar] [CrossRef] [PubMed]
- De Marchi, M. Detailed characterization of plant-based burgers. Sci. Rep. 2021, 11, 2049. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Jones, O.G.; Campanella, O.H. Plant protein-based fibers: Fabrication, characterization and potential food applications. Crit. Rev. Food Sci. 2021, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Mulla, M.Z.; Subramanian, P.; Dar, B.N. Functionalization of legume proteins using high pressure processing: Effect on technofunctional properties and digestibility of legume proteins. LWT-Food Sci. Technol. 2022, 158, 113106. [Google Scholar] [CrossRef]
- Ozorio, L.; Mellinger-Silva, C.; Cabral, L.M.C. The influence of peptidases in intestinal brush border membranes on the absorption of oligopeptides from whey protein hydrolysate: An ex vivo study using an ussing chamber. Foods 2020, 9, 1415. [Google Scholar] [CrossRef]
- Pekala, J.; Patkowska-Sokola, B.; Bodkowski, R. L-carnitine-metabolic functions and meaning in humans’ life. Curr. Drug Metab. 2011, 12, 667–678. [Google Scholar] [CrossRef]
- Yoshizawa, F. Notable functions of branched chain amino acids as biological regulators. J. Pharmacol. Sci. 2011, 115, 39. [Google Scholar]
- Wu, G.; Wu, Z.; Dai, Z. Dietary requirements of “nutritionally non-essential amino acids” by animals and humans. Amino Acids. 2013, 44, 1107–1113. [Google Scholar] [CrossRef]
- Gallego, M.; Mauri, L.; Aristoy, M.C. Antioxidant peptides profile in dry-cured ham as affected by gastrointestinal digestion. J. Funct. Foods 2020, 69, 103956. [Google Scholar] [CrossRef]
- Fu, Y.; Young, J.; Lokke, M.; Lametsch, R.; Aluko, R.E.; Therkildsen, M. Revalorisation of bovine collagen as a potential precursor of angiotensin 1-converting enzyme (ACE) inhibitory peptides based on in silico and in vitro protein digestions. J. Funct. Foods 2016, 24, 196–206. [Google Scholar] [CrossRef]
- Wang, S.; Su, G.; Fan, J.; Zhao, M.; Wu, J. Arginine-containing peptides derived from walnut protein against cognitive and memory impairment in scopolamine-induced zebrafish: Design, release, and neuroprotection. J. Agric. Food Chem. 2022, 70, 11579–11590. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Mohan, A. Mechanisms of food protein-derived antihypertensive peptides other than ACE inhibition. J. Funct. Foods 2014, 8, 45–52. [Google Scholar] [CrossRef]
- Fleenor, B.S.; Seals, D.R.; Zigler, M.L. Superoxide-lowering therapy with TEMPOL reverses arterial dysfunction with aging in mice. Aging Cell 2012, 11, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Su, G.; Zhang, X. Characterization and exploration of potential neuroprotective peptides in walnut (Juglans regia) protein hydrolysate against cholinergic system damage and oxidative stress in scopolamine-induced cognitive and memory impairment mice and zebrafish. J. Agric. Food Chem. 2021, 69, 2773–2783. [Google Scholar] [CrossRef]
- Wang, S.; Sun-Waterhouse, D.; Waterhouse, G.I.N. Effects of food-derived bioactive peptides on cognitive deficits and memory decline in neurodegenerative diseases: A review. Trends Food Sci. Technol. 2021, 116, 712–732. [Google Scholar] [CrossRef]
- Fan, H.; Xu, Q.; Hong, H.; Wu, J. Stability and transport of spent hen-derived ACE-inhibitory peptides IWHHT, IWH, and IW in human intestinal Caco-2 cell monolayers. J. Agric. Food Chem. 2018, 66, 11347–11354. [Google Scholar] [CrossRef]
- Touyz, R.; Schiffrin, E. Reactive oxygen species in vascular biology: Implications in hypertension. Histochem. Cell Biol. 2004, 122, 339–352. [Google Scholar] [CrossRef]
- Odegaard, A.O.; Jacobs, D.R., Jr.; Sanchez, O.A. Oxidative stress, inflammation, endothelial dysfunction and incidence of type 2 diabetes. Cardiovasc. Diabetol. 2016, 15, 51. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Bhullar, K.S.; Wu, J. Spent hen muscle protein-derived RAS regulating peptides show antioxidant activity in vascular cells. Antioxidants 2021, 10, 290. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Fan, H.; Wu, J. Egg white-derived antihypertensive peptide IRW (Ile-Arg-Trp) inhibits angiotensin II-stimulated migration of vascular smooth muscle cells via angiotensin type I receptor. J. Agric. Food Chem. 2018, 66, 5133–5138. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Liao, W.; Davidge, S.T.; Wu, J. Chicken muscle-derived ACE2 upregulating peptide VVHPKESF inhibits angiotensin II-stimulated inflammation in vascular smooth muscle cells via the ACE2/Ang (1−7)/MasR axis. J. Agric. Food Chem. 2022, 70, 6397–6406. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BfP | BfPP | ByP | ByPP | ImP | ImPP | |
|---|---|---|---|---|---|---|
| DH (%) | 8.26 ± 0.67 | 10.66 ± 0.83 | 4.92 ± 0.11 | 7.94 ± 0.73 | 6.09 ± 0.59 | 7.48 ± 0.40 |
| Amino acids composition (g/100 g) | ||||||
| Total amino acids | 87.61 ± 1.51 a | 87.64 ± 1.43 a | 70.80 ± 1.26 b | 75.11 ± 1.73 c | 67.56 ± 1.20 d | 69.35 ± 0.54 d |
| EAA | 45.75 ± 0.84 a | 46.04 ± 1.13 a | 40.76 ± 0.53 b | 42.91 ± 1.28 c | 40.61 ± 0.71 b | 41.60 ± 1.34 b,c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Zhao, M.; Fan, H.; Wu, J. Peptidomics Study of Plant-Based Meat Analogs as a Source of Bioactive Peptides. Foods 2023, 12, 1061. https://doi.org/10.3390/foods12051061
Wang S, Zhao M, Fan H, Wu J. Peptidomics Study of Plant-Based Meat Analogs as a Source of Bioactive Peptides. Foods. 2023; 12(5):1061. https://doi.org/10.3390/foods12051061
Chicago/Turabian StyleWang, Shuguang, Mouming Zhao, Hongbing Fan, and Jianping Wu. 2023. "Peptidomics Study of Plant-Based Meat Analogs as a Source of Bioactive Peptides" Foods 12, no. 5: 1061. https://doi.org/10.3390/foods12051061