
Metagenomic Insights into the Anti-Obesity Effect of a Polysaccharide from Saccharina japonica
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of SjC from S. japonica
2.2. Structural Analysis of SjC
2.3. Animal Experiment
2.4. Biochemical Measurements
2.5. Gut Microbiota Analysis by 16S rRNA Gene and Shotgun Metagenomic Sequencing
2.6. Linking 16S rRNA Gene Sequences with Metagenome-Assembled Genomes
2.7. Prediction of Bacteria with Fucoidan-Degrading Potential
2.8. Statistical Analysis
3. Results
3.1. Structure Analysis of SjC
3.2. SjC Supplementation Prevented HFD-Induced Obesity and Metabolic Disorders
3.3. SjC Supplementation Alleviated HFD-Induced Gut Microbiota Dysbiosis at Species Level
3.4. SjC Supplementation Alleviated HFD-Induced Gut Microbiota Dysbiosis at MAG Level
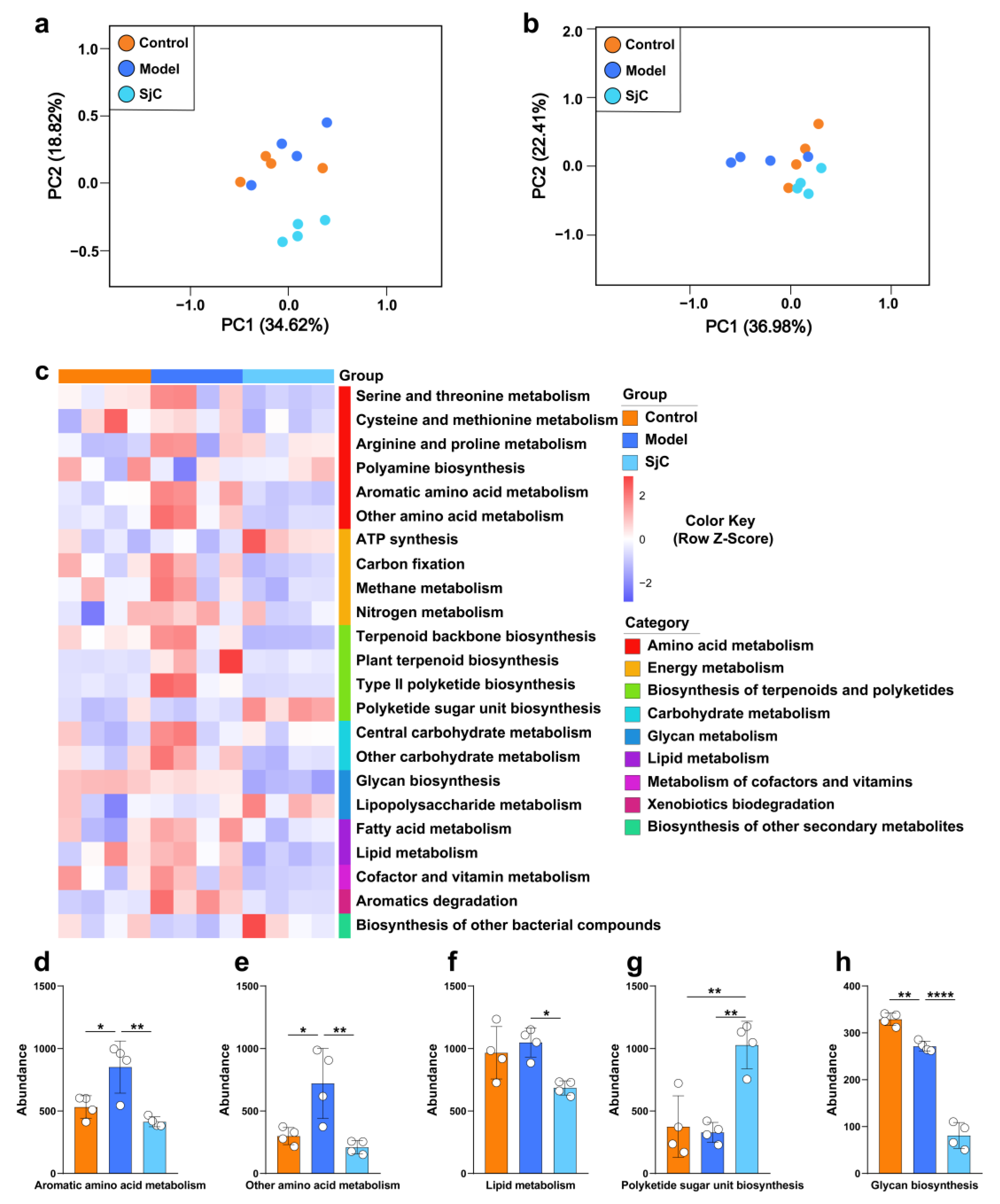
3.5. SjC Supplementation Suppressed Amino Acid Metabolism of Gut Microbiota
3.6. SjC Supplementation Enriched Fucoidan-Degrading Gut Bacteria That Correlated with Bodyweight
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pi-Sunyer, X. The Medical Risks of Obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Yazici, D.; Sezer, H. Insulin Resistance, Obesity and Lipotoxicity. Adv. Exp. Med. Biol. 2017, 960, 277–304. [Google Scholar] [PubMed]
- Zhang, Y.; Zuo, J.; Yan, L.; Cheng, Y.; Li, Q.; Wu, S.; Chen, L.; Thring, R.W.; Yang, Y.; Gao, Y.; et al. Sargassum fusiforme Fucoidan Alleviates High-Fat Diet-Induced Obesity and Insulin Resistance Associated with the Improvement of Hepatic Oxidative Stress and Gut Microbiota Profile. J. Agric. Food Chem. 2020, 68, 10626–10638. [Google Scholar] [CrossRef] [PubMed]
- Adam, C.L.; Gratz, S.W.; Peinado, D.I.; Thomson, L.M.; Garden, K.E.; Williams, P.A.; Richardson, A.J.; Ross, A.W. Effects of Dietary Fibre (Pectin) and/or Increased Protein (Casein or Pea) on Satiety, Body Weight, Adiposity and Caecal Fermentation in High Fat Diet-Induced Obese Rats. PLoS ONE 2016, 11, e0155871. [Google Scholar] [CrossRef] [PubMed]
- Aronsson, L.; Huang, Y.; Parini, P.; Korach-André, M.; Håkansson, J.; Gustafsson, J.Å.; Pettersson, S.; Arulampalam, V.; Rafter, J. Decreased Fat Storage by Lactobacillus paracasei Is Associated with Increased Levels of Angiopoietin-Like 4 Protein (ANGPTL4). PLoS ONE 2010, 5, e13087. [Google Scholar] [CrossRef]
- Nagpal, R.; Kumar, M.; Yadav, A.; Hemalatha, R.; Yadav, H.; Marotta, F.; Yamashiro, Y. Gut Microbiota in Health and Disease: An Overview Focused on Metabolic Inflammation. Benef. Microbes 2016, 7, 181–194. [Google Scholar] [CrossRef]
- Gao, J.; Lin, L.; Sun, B.; Zhao, M. Comparison Study on Polysaccharide Fractions from Laminaria japonica: Structural Characterization and Bile Acid Binding Capacity. J. Agric. Food Chem. 2017, 65, 9790–9798. [Google Scholar] [CrossRef]
- Swathi, N.; Kumar, A.G.; Parthasarathy, V.; Sankarganesh, P. Isolation of Enteromorpha Species and Analyzing Its Crude Extract for the Determination of in Vitro Antioxidant and Antibacterial Activities. Biomass Convers. Biorefinery 2022, 23, 1–10. [Google Scholar] [CrossRef]
- Wan, F.; Ma, F.; Wu, J.; Qiao, X.; Chen, M.; Li, W.; Ma, L. Effect of Lycium barbarum Polysaccharide on Decreasing Serum Amyloid A3 Expression through Inhibiting Nf-Kappab Activation in a Mouse Model of Diabetic Nephropathy. Anal. Cell. Pathol. Amst. 2022, 2022, 7847135. [Google Scholar]
- Zhang, Y.; Yang, L.; Zhao, N.; Hong, Z.; Cai, B.; Le, Q.; Yang, T.; Shi, L.; He, J. Soluble Polysaccharide Derived from Laminaria japonica Attenuates Obesity-Related Nonalcoholic Fatty Liver Disease Associated with Gut Microbiota Regulation. Mar. Drugs 2021, 19, 699. [Google Scholar] [CrossRef]
- Li, N.; Fu, X.; Xiao, M.; Wei, X.; Yang, M.; Liu, Z.; Mou, H. Enzymatic Preparation of a Low-Molecular-Weight Polysaccharide Rich in Uronic Acid from the Seaweed Laminaria japonica and Evaluation of Its Hypolipidemic Effect in Mice. Food Funct. 2020, 11, 2395–2405. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, F.; Gao, Y.; Xue, C.-H.; Li, R.W.; Tang, Q.-J. Transcriptome Analysis Revealed Anti-Obesity Effects of the Sodium Alginate in High-Fat Diet-Induced Obese Mice. Int. J. Biol. Macromol. 2018, 115, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, Y.; Chen, X.Q.; Aweya, J.J.; Cheong, K.L. Catabolism of Saccharina japonica Polysaccharides and Oligosaccharides by Human Fecal Microbiota. LWT Food Sci. Technol. 2020, 130, 109635. [Google Scholar] [CrossRef]
- Zhang, X.; Aweya, J.J.; Huang, Z.-X.; Kang, Z.-Y.; Bai, Z.-H.; Li, K.-H.; He, X.-T.; Liu, Y.; Chen, X.-Q.; Cheong, K.-L. In Vitro Fermentation of Gracilaria lemaneiformis Sulfated Polysaccharides and Its Agaro-Oligosaccharides by Human Fecal Inocula and Its Impact on Microbiota. Carbohydr. Polym. 2020, 234, 115894. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.C.; Hu, J.L.; Li, J.; Wang, J.; Zhang, X.Y.; Wu, X.Y.; Li, X.; Guo, Z.B.; Zou, L.; Wu, D.T. Physicochemical Characteristics and Biological Activities of Soluble Dietary Fibers Isolated from the Leaves of Different Quinoa Cultivars. Food Res. Int. 2023, 163, 112166. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Wang, Y.; Zeng, L.; Dong, J.; Bi, Q.; Yang, X.; Che, Y.; He, S.; Yu, J. Polysaccharides from Polygonatum kingianum Improve Glucose and Lipid Metabolism in Rats Fed a High Fat Diet. Biomed. Pharmacother. 2020, 125, 109910. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.T.; He, Y.; Yuan, Q.; Wang, S.; Gan, R.Y.; Hu, Y.C.; Zou, L. Effects of Molecular Weight and Degree of Branching on Microbial Fermentation Characteristics of Okra Pectic-polysaccharide and its Selective Impact on Gut Microbial Composition. Food Hydrocoll. 2020, 132, 107897. [Google Scholar] [CrossRef]
- Wei, B.; Zhang, B.; Du, A.-Q.; Zhou, Z.-Y.; Lu, D.-Z.; Zhu, Z.-H.; Ke, S.-Z.; Wang, S.-J.; Yu, Y.-L.; Chen, J.-W.; et al. Saccharina japonica Fucan Suppresses High Fat Diet-Induced Obesity and Enriches Fucoidan-Degrading Gut Bacteria. Carbohydr. Polym. 2022, 290, 119411. [Google Scholar] [CrossRef]
- Wu, D.-T.; An, L.-Y.; Liu, W.; Hu, Y.-C.; Wang, S.-P.; Zou, L. In Vitro Fecal Fermentation Properties of Polysaccharides from Tremella fuciformis and Related Modulation Effects on Gut Microbiota. Food Res. Int. 2022, 156, 111185. [Google Scholar] [CrossRef]
- Aleissa, M.S.; Alkahtani, S.; Abd Eldaim, M.A.; Ahmed, A.M.; Bungau, S.G.; Almutairi, B.; Bin-Jumah, M.; AlKahtane, A.A.; Alyousif, M.S.; Abdel-Daim, M.M. Fucoidan Ameliorates Oxidative Stress, Inflammation, DNA Damage, and Hepatorenal Injuries in Diabetic Rats Intoxicated with Aflatoxin B(1). Oxidative Med. Cell. Longev. 2020, 2020, 9316751. [Google Scholar] [CrossRef]
- Dinesh, S.; Menon, T.; Hanna, L.E.; Suresh, V.; Sathuvan, M.; Manikannan, M. In Vitro Anti-HIV-1 Activity of Fucoidan from Sargassum swartzii. Int. J. Biol. Macromol. 2016, 82, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Ke, S.; Yu, Y.; Xu, Q.; Zhang, B.; Wang, S.; Jin, W.; Wei, B.; Wang, H. Composition-Activity Relationships of Polysaccharides from Saccharina japonica in Regulating Gut Microbiota in Short-Term High-Fat Diet-Fed Mice. J. Agric. Food Chem. 2021, 69, 11121–11130. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.-W.; Zhou, T.-S.; Qiu, W.-H.; Wang, Y.-K.; Xu, Q.-L.; Ke, S.-Z.; Wang, S.-J.; Jin, W.-H.; Chen, J.-W.; Zhang, H.-W.; et al. Characterization and Hypoglycemic Effects of Sulfated Polysaccharides Derived from Brown Seaweed Undaria pinnatifida. Food Chem. 2020, 341, 128148. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using Qiime 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows—Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Lee, I.; Ouk Kim, Y.; Park, S.C.; Chun, J. Orthoani: An Improved Algorithm and Software for Calculating Average Nucleotide Identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. Ncbi Blast: A Better Web Interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Sichert, A.; Corzett, C.H.; Schechter, M.S.; Unfried, F.; Markert, S.; Becher, D.; Fernandez-Guerra, A.; Liebeke, M.; Schweder, T.; Polz, M.F.; et al. Verrucomicrobia Use Hundreds of Enzymes to Digest the Algal Polysaccharide Fucoidan. Nat. Microbiol. 2020, 5, 1026–1039. [Google Scholar] [CrossRef]
- Cheng, H.; Huang, G.; Huang, H. The Antioxidant Activities of Garlic Polysaccharide and Its Derivatives. Int. J. Biol. Macromol. 2020, 145, 819–826. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, G.; Yu, Z.; Song, X.; Li, X.; Yang, Y.; Wang, L.; Liu, L.; Dai, J. Purification, Characterization and Antiglycation Activity of a Novel Polysaccharide from Black Currant. Food Chem. 2016, 199, 694–701. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, T.; Jin, Z.-Y.; Xu, X.-M.; Wang, J.-H.; Zha, X.-Q.; Chen, H.-Q. Structural Characterisation, Physicochemical Properties and Antioxidant Activity of Polysaccharide from Lilium lancifolium Thunb. Food Chem. 2015, 169, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.; Bell, R.; Klag, K.A.; Lee, S.-H.; Soto, R.; Ghazaryan, A.; Buhrke, K.; Ekiz, H.A.; Ost, K.S.; Boudina, S.; et al. T Cell-Mediated Regulation of the Microbiota Protects against Obesity. Science 2019, 365, eaat9351. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yao, W.; Li, B.; Qian, S.; Wei, B.; Gong, S.; Wang, J.; Liu, M.; Wei, M. Nuciferine Modulates the Gut Microbiota and Prevents Obesity in High-Fat Diet-Fed Rats. Exp. Mol. Med. 2020, 52, 1959–1975. [Google Scholar] [CrossRef]
- Zhou, K. Strategies to Promote Abundance of Akkermansia muciniphila, an Emerging Probiotics in the Gut, Evidence from Dietary Intervention Studies. J. Funct. Foods 2017, 33, 194–201. [Google Scholar] [CrossRef]
- Corb Aron, R.A.; Abid, A.; Vesa, C.M.; Nechifor, A.C.; Behl, T.; Ghitea, T.C.; Munteanu, M.A.; Fratila, O.; An-dronie-Cioara, F.L.; Toma, M.M.; et al. Recognizing the Benefits of Pre-/Probiotics in Metabolic Syndrome and Type 2 Diabetes Mellitus Considering the Influence of Akkermansia muciniphila as a Key Gut Bacterium. Microorganisms 2021, 9, 618. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.L.; Chiang, E.; Plantinga, A.; Carey, H.V.; Suen, G.; Swoap, S.J. Effect of stevia on the gut microbiota and glucose tolerance in a murine model of diet-induced obesity. FEMS Microbiol. Ecol. 2020, 96, fiaa079. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.F.; Lai, S.S.; Chen, Y.H.; Liu, D.; Liu, B.; Ai, C.; Wan, X.Z.; Gao, L.Y.; Chen, X.H.; Zhao, C. Anti-Diabetic Effect of Oligosaccharides from Seaweed Sargassum confusum via Jnk-Irs1/Pi3k Signalling Pathways and Regulation of Gut Microbiota. Food Chem. Toxicol. 2019, 131, 110562. [Google Scholar] [CrossRef]
- Dai, Z.-L.; Wu, G.; Zhu, W.-Y. Amino Acid Metabolism in Intestinal Bacteria: Links between Gut Ecology and Host Health. Front. Biosci. 2011, 16, 1768–1786. [Google Scholar] [CrossRef]
- Chen, M.H.; Zhou, J.; Wu, C.Y.; Zhang, W.; Long, F.; Zhou, S.S.; Xu, J.D.; Wu, J.; Zou, Y.T.; Li, S.L.; et al. Gut Microbiota Influenced the Xenograft Mc38 Tumor Growth Potentially through Interfering Host Lipid and Amino Acid Metabolisms, Basing on the Integrated Analysis of Microbiome and Metabolomics. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2022, 1192, 123136. [Google Scholar] [CrossRef]
- Song, S.Y.; Lou, Y.; Mao, Y.Y.; Wen, X.H.; Fan, M.Q.; He, Z.X.; Shen, Y.; Wen, C.P.; Shao, T.J. Alteration of Gut Microbiome and Correlated Amino Acid Metabolism Contribute to Hyperuricemia and Th17-Driven Inflammation in Uox-Ko Mice. Front. Immunol. 2022, 13, 804306. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Y.; Ai, C.; Tian, W.; Wen, C.; Song, S.; Zhu, B. An Acidic Polysaccharide from Patinopecten yessoensis Skirt Prevents Obesity and Improves Gut Microbiota and Metabolism of Mice Induced by High-Fat Diet. Food Res. Int. 2022, 154, 110980. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.-L.; Li, S.-Z.; Xiao, P.-T.; Cai, Y.-Y.; Chu, C.; Chen, B.-Z.; Li, P.; Li, J.; Liu, E.-H. Citrus Polymethoxyflavones Attenuate Metabolic Syndrome by Regulating Gut Microbiome and Amino Acid Metabolism. Sci. Adv. 2020, 6, eaax6208. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MAG No. | Spearman’s Correlation (MAG vs. Bodyweight). a | Minimum JS Divergence b | 16S rRNA Blasted against MAG c | |||
|---|---|---|---|---|---|---|
| 16S rRNA No. | 16S rRNA Taxonomic Classification | Identity (%) | Alignment Length | |||
| bin.293 | −0.191 (p > 0.05) | 0.0505 | repseq1 | g_Alistipes; s_Alistipes_inops | 91.667 | 48 |
| bin.72 | 0.402 (p > 0.05) | 0.0419 | repseq2 | f_Atopobiaceae | 98.795 | 83 |
| bin.326 | −0.136 (p > 0.05) | 0.0493 | repseq3 | f_Lachnospiraceae | 97.872 | 423 |
| bin.33 | 0.599 (*) | 0.0454 | repseq4 | g_Desulfovibrio | 99.342 | 304 |
| bin.53 | 0.492 (*) | 0.0570 | repseq5 | g_Desulfovibrio | 100 | 66 |
| bin.50 | −0.247 (p > 0.05) | 0.0633 | repseq6 | g_Helicobacter | 99.213 | 127 |
| bin.324 | −0.413 (p > 0.05) | 0.0462 | repseq7 | f_Oscillospiraceae; g_uncultured | 96.948 | 426 |
| bin.80 | 0.870 (***) | 0.0304 | repseq8 | g_Akkermansia | 100 | 430 |
| bin.271 | 0.682 (**) | 0.0385 | repseq9 | g_Muribaculum | 100 | 443 |
| bin.216 | −0.506 (*) | 0.0329 | repseq10 | g_Muribaculum | 100 | 443 |
| bin.111 | 0.263 (p > 0.05) | 0.0436 | repseq11 | g_Turicibacter | 93.438 | 320 |
| bin.256 | −0.489 (*) | 0.0326 | repseq12 | g_Prevotellaceae_UCG-001; s_uncultured | 90.787 | 445 |
| bin.322 | −0.432 (p > 0.05) | 0.0473 | repseq13 | g_Alistipes | 97.065 | 443 |
| bin.120 | 0.845 (***) | 0.0340 | repseq14 | g_Muribaculaceae; s_uncultured | 99.097 | 443 |
| bin.46 | 0.514 (*) | 0.0453 | repseq15 | g_Blautia; s_uncultured | 88.961 | 154 |
| bin.268 | −0.390 (p > 0.05) | 0.0437 | repseq16 | f_Lachnospiraceae | 97.872 | 423 |
| bin.161 | 0.567 (*) | 0.0529 | repseq17 | g_Lachnospiraceae_NK4A136_group | 99.291 | 423 |
| bin.117 | −0.475 (p > 0.05) | 0.0581 | NA | |||
| bin.302 | 0.712 (**) | 0.0381 | repseq19 | g_Allobaculum; s_uncultured | 90.667 | 450 |
| bin.219 | 0.594 | 0.0519 | repseq20 | g_Clostridium_sp; s_uncultured | 98.126 | 427 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Lu, D.; Wang, H.; Zhou, Z.; Luo, X.; Ma, M.; Ke, S.; Wang, H.; Yu, Y.; Wei, B. Metagenomic Insights into the Anti-Obesity Effect of a Polysaccharide from Saccharina japonica. Foods 2023, 12, 665. https://doi.org/10.3390/foods12030665
Song Y, Lu D, Wang H, Zhou Z, Luo X, Ma M, Ke S, Wang H, Yu Y, Wei B. Metagenomic Insights into the Anti-Obesity Effect of a Polysaccharide from Saccharina japonica. Foods. 2023; 12(3):665. https://doi.org/10.3390/foods12030665
Chicago/Turabian StyleSong, Ying, Dongze Lu, Honggang Wang, Zhenyi Zhou, Xian Luo, Manjing Ma, Songze Ke, Hong Wang, Yanlei Yu, and Bin Wei. 2023. "Metagenomic Insights into the Anti-Obesity Effect of a Polysaccharide from Saccharina japonica" Foods 12, no. 3: 665. https://doi.org/10.3390/foods12030665