
Capsular Exopolysaccharides from Two Streptococcus thermophilus Strains Differ in Their Moisture Sorption Behavior
 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cultivation Conditions
2.3. Biomass Determination and Exopolysaccharide Quantification
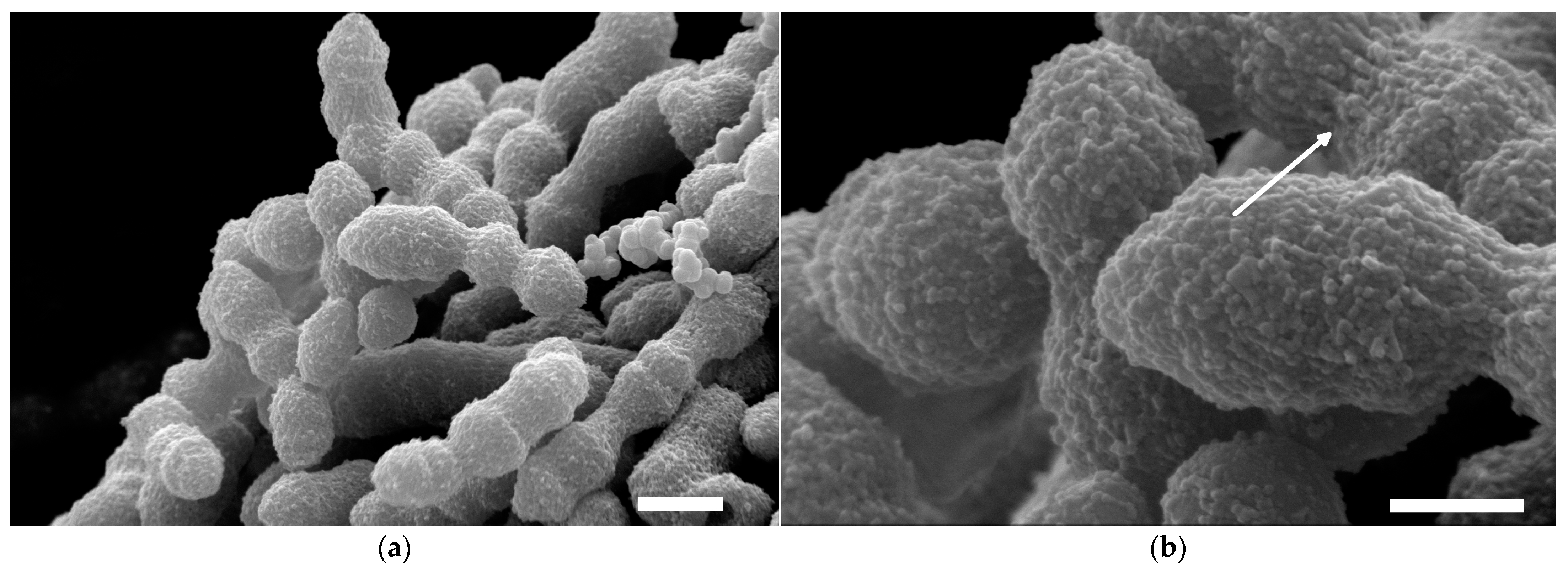
2.4. Visualization of Capsular Exopolysaccharides
2.5. Preparation of Cell and Exopolysaccharide Isolates
2.6. Structural Composition of Free Exopolysaccharides
2.7. Macromolecular Properties of Exopolysaccharides
2.7.1. Intrinsic Viscosity
2.7.2. Molecular Mass
2.7.3. Water Binding Capacity
2.8. Statistics
3. Results and Discussion
3.1. Microbial Production of Free and Capsular Exopolysaccharides
3.2. Exopolysaccharide Structure
3.3. Macromolecular Properties of Free Exopolysaccharides
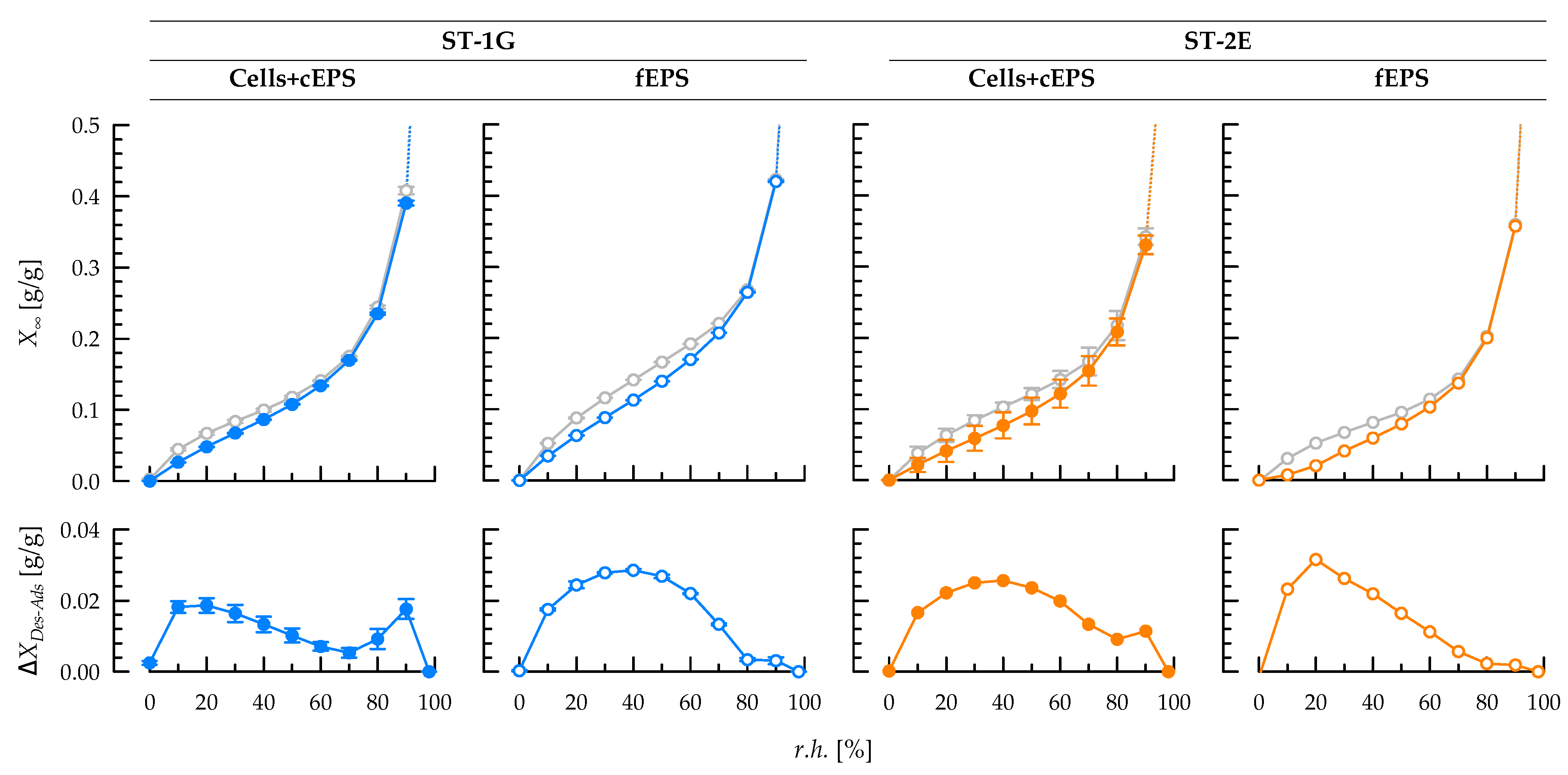
3.4. Sorption Behaviour of Exopolysaccharides at Equilibrium State
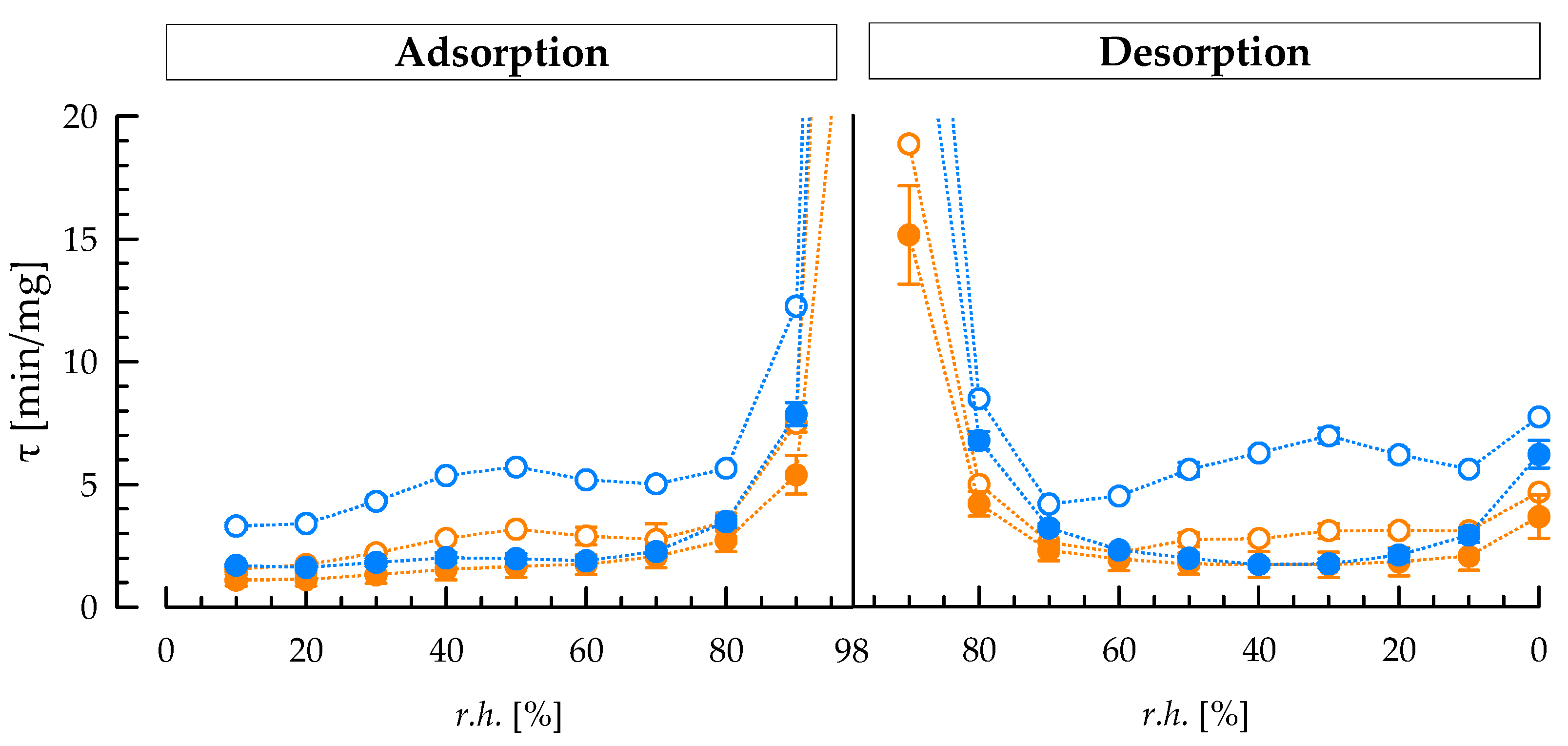
3.5. Sorption Kinetics of Exopolysaccharides
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zeidan, A.A.; Poulsen, V.K.; Janzen, T.; Buldo, P.; Derkx, P.M.F.; Øregaard, G.; Neves, A.R. Polysaccharide Production by Lactic Acid Bacteria: From Genes to Industrial Applications. FEMS Microbiol. Rev. 2017, 41, 168–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mende, S.; Rohm, H.; Jaros, D. Influence of Exopolysaccharides on the Structure, Texture, Stability and Sensory Properties of Yoghurt and Related Products. Int. Dairy J. 2016, 52, 57–71. [Google Scholar] [CrossRef]
- Nachtigall, C.; Vogel, C.; Rohm, H.; Jaros, D. How Capsular Exopolysaccharides Affect Cell Surface Properties of Lactic Acid Bacteria. Microorganisms 2020, 8, 1904. [Google Scholar] [CrossRef]
- Hassan, A.N.; Frank, J.F.; Farmer, M.A.; Schmidt, K.A.; Shalabi, S.I. Observation of Encapsulated Lactic Acid Bacteria Using Confocal Scanning Laser Microscopy. J. Dairy Sci. 1995, 78, 2624–2628. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.N.; Frank, J.F.; Shalabi, S.I. Factors Affecting Capsule Size and Production by Lactic Acid Bacteria Used as Dairy Starter Cultures. Int. J. Food Microbiol. 2001, 64, 199–203. [Google Scholar] [CrossRef]
- Hassan, A.N.; Frank, J.F.; Schmidt, K.A.; Shalabi, S.I. Textural Properties of Yogurt Made with Encapsulated Nonropy Lactic Cultures. J. Dairy Sci. 1996, 79, 2098–2103. [Google Scholar] [CrossRef]
- Amatayakul, T.; Sherkat, F.; Shah, N.P. Physical Characteristics of Set Yoghurt Made with Altered Casein to Whey Protein Ratios and EPS-Producing Starter Cultures at 9 and 14% Total Solids. Food Hydrocoll. 2006, 20, 314–324. [Google Scholar] [CrossRef]
- Mende, S.; Mentner, C.; Thomas, S.; Rohm, H.; Jaros, D. Exopolysaccharide Production by Three Different Strains of Streptococcus thermophilus and Its Effect on Physical Properties of Acidified Milk. Eng. Life Sci. 2012, 12, 466–474. [Google Scholar] [CrossRef]
- Timmermann, E.O. Multilayer Sorption Parameters: BET or GAB Values? Colloids Surf. A 2003, 220, 235–260. [Google Scholar] [CrossRef] [Green Version]
- Al-Muhtaseb, A.H.; McMinn, W.A.M.; Magee, T.R.A. Moisture Sorption Isotherm Characteristics of Food Products: A Review. Food Bioprod. Process. 2002, 80, 118–128. [Google Scholar] [CrossRef]
- Thybring, E.E.; Glass, S.V.; Zelinka, S.L. Kinetics of Water Vapor Sorption in Wood Cell Walls: State of the Art and Research Needs. Forrests 2019, 10, 704. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Soer, W.-J.; Scheerder, J.; Satgurunathan, G.; Keddie, J.L. Water Vapor Sorption and Diffusion in Secondary Dispersion Barrier Coatings: A Critical Comparison with Emulsion Polymers. ACS Appl. Mater. Interfaces 2015, 7, 12147–12157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murrieta-Pazos, I.; Galet, L.; Patry, S.; Gaiani, C.; Scher, J. Evolution of Particle Structure during Water Sorption Observed on Different Size Fractions of Durum Wheat Semolina. Powder Technol. 2014, 255, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Crank, J. A Theoretical Investigation of the Influence of Molecular Relaxation and Internal Stress on Diffusion in Polymers. J. Polym. Sci. 1953, 11, 151–168. [Google Scholar] [CrossRef]
- Wimmer, R.; Kläusler, O.; Niemz, P. Water Sorption Mechanisms of Commercial Wood Adhesive Films. Wood Sci. Technol. 2013, 47, 763–775. [Google Scholar] [CrossRef]
- Willems, W. Thermally Limited Wood Moisture Changes: Relevance for Dynamic Vapour Sorption Experiments. Wood Sci. Technol. 2017, 51, 751–770. [Google Scholar] [CrossRef]
- Kohler, R.; Dück, R.; Ausperger, B.; Alex, R. A Numeric Model for the Kinetics of Water Vapor Sorption on Cellulosic Reinforcement Fibers. Compos. Interfaces 2003, 10, 255–276. [Google Scholar] [CrossRef]
- Hill, C.A.S.; Norton, A.; Newman, G. Analysis of the Water Vapour Sorption Behaviour of Sitka Spruce [Picea Sitchensis (Bongard) Carr.] Based on the Parallel Exponential Kinetics Model. Holzforschung 2010, 64, 469–473. [Google Scholar] [CrossRef]
- Madamba, P.S.; Driscoll, R.H.; Buckle, K.A. The Thin-Layer Drying Characteristics of Garlic Slices. J. Food Eng. 1996, 29, 75–97. [Google Scholar] [CrossRef]
- Rahman, M.S.; Perera, C.O.; Thebaud, C. Desorption Isotherm and Heat Pump Drying Kinetics of Peas. Food Res. Int. 1997, 30, 485–491. [Google Scholar] [CrossRef]
- Tang, X.; De Rooij, M.R.; Van Duynhoven, J.; Van Breugel, K. Dynamic Volume Change Measurements of Cereal Materials by Environmental Scanning Electron Microscopy and Videomicroscopy. J. Microsc. 2008, 230, 100–107. [Google Scholar] [CrossRef]
- Thybring, E.E.; Boardman, C.R.; Glass, S.V.; Zelinka, S.L. The Parallel Exponential Kinetics Model Is Unfit to Characterize Moisture Sorption Kinetics in Cellulosic Materials. Cellulose 2019, 26, 723–735. [Google Scholar] [CrossRef]
- Surber, G.; Rohm, H.; Jaros, D. The Role of Exopolysaccharide-Producing Streptococcus thermophilus on Physical Properties of Stirred Skim Milk Gel. Dairy 2022, 3, 761–775. [Google Scholar] [CrossRef]
- Nachtigall, C.; Surber, G.; Herbi, F.; Wefers, D.; Jaros, D.; Rohm, H. Production and Molecular Structure of Heteropolysaccharides from Two Lactic Acid Bacteria. Carbohydr. Polym. 2020, 236, 116019. [Google Scholar] [CrossRef] [PubMed]
- Surber, G.; Mende, S.; Jaros, D.; Rohm, H. Clustering of Streptococcus thermophilus Strains to Establish a Relation between Exopolysaccharide Characteristics and Gel Properties of Acidified Milk. Foods 2019, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Nachtigall, C.; Surber, G.; Bulla, J.; Rohm, H.; Jaros, D. Pilot Scale Isolation of Exopolysaccharides from Streptococcus thermophilus DGCC7710: Impact of Methodical Details on Macromolecular Properties and Techno-Functionality. Eng. Life Sci. 2021, 21, 220–232. [Google Scholar] [CrossRef]
- Fels, L.; Jakob, F.; Vogel, R.F.; Wefers, D. Structural Characterization of the Exopolysaccharides from Water Kefir. Carbohydr. Polym. 2018, 189, 296–303. [Google Scholar] [CrossRef]
- Nachtigall, C.; Berger, C.; Kovanović, T.; Wefers, D.; Jaros, D.; Rohm, H. Shear Induced Molecular Changes of Exopolysaccharides from Lactic Acid Bacteria. Food Hydrocoll. 2019, 97, 105181. [Google Scholar] [CrossRef]
- Antoniou, E.; Themistou, E.; Sarkar, B.; Tsianou, M.; Alexandridis, P. Structure and Dynamics of Dextran in Binary Mixtures of a Good and a Bad Solvent. Coll. Polym. Sci. 2010, 288, 1301–1312. [Google Scholar] [CrossRef]
- Passauer, L.; Struch, M.; Schuldt, S.; Appelt, J.; Schneider, Y.; Jaros, D.; Rohm, H. Dynamic Moisture Sorption Characteristics of Xerogels from Water-Swellable Oligo(Oxyethylene) Lignin Derivatives. ACS Appl. Mater. Interf. 2012, 4, 5852–5862. [Google Scholar] [CrossRef] [PubMed]
- Skov Sørensen, U.B.; Yao, K.; Yang, Y.; Tettelin, H.; Kilian, M. Capsular Polysaccharide Expression in Commensal Streptococcus Species: Genetic and Antigenic Similarities to Streptococcus pneumoniae. mBio 2016, 7, e01844-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, J. Recent Insights in Microbial Exopolysaccharide Biosynthesis and Engineering Strategies. Curr. Opin. Biotechnol. 2018, 53, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Cui, Y.; Yue, F.; Liu, L.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Exopolysaccharides Produced by Lactic Acid Bacteria and Bifidobacteria: Structures, Physiochemical Functions and Applications in the Food Industry. Food Hydrocoll. 2019, 94, 475–499. [Google Scholar] [CrossRef]
- Zhou, Y.; Cui, Y.; Qu, X. Exopolysaccharides of Lactic Acid Bacteria: Structure, Bioactivity and Associations: A Review. Carbohydr. Polym. 2019, 207, 317–332. [Google Scholar] [CrossRef]
- Bayer, M.E. Visualization of the Bacterial Polysaccharide Capsule. In Bacterial Capsules; Jann, K., Jann, B., Eds.; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 1990; Volume 150, pp. 129–157. [Google Scholar]
- Springer, E.L.; Roth, I.L. Ultrastructure of the Capsule of Klebsiella pneumoniae and Slime of Enterobacter aerogenes Revealed by Freeze Etching. Archiv. Mikrobiol. 1973, 93, 277–286. [Google Scholar] [CrossRef]
- Stellner, K.; Saito, H.; Hakomori, S.I. Determination of Aminosugar Linkages in Glycolipids by Methylation: Aminosugar Linkages of Ceramide Pentasaccharides of Rabbit Erythrocytes and of Forssman Antigen. Arch. Biochem. Biophys. 1973, 155, 464–472. [Google Scholar] [CrossRef]
- Faber, E.J.; Zoon, P.; Kamerling, J.P.; Vliegenthart, J.F.G. The Exopolysaccharides Produced by Streptococcus thermophilus Rs and Sts Have the Same Repeating Unit but Differ in Viscosity of Their Milk Cultures. Carbohydr. Res. 1998, 310, 269–276. [Google Scholar] [CrossRef]
- Ruas-Madiedo, P.; Tuinier, R.; Kanning, M.; Zoon, P. Role of Exopolysaccharides Produced by Lactococcus lactis subsp. cremoris on the Viscosity of Fermented Milks. Int. Dairy J. 2002, 12, 689–695. [Google Scholar]
- Nachtigall, C. Makromolekulare Eigenschaften extrazellulärer polymerer Kohlenhydrate von ausgewählten Milchsäurebakterien, Thesis, Technische Universität Dresden. 2021. Available online: https://nbn-resolving.org/urn:nbn:de:bsz:14-qucosa2-769682 (accessed on 26 October 2022).
- Gentès, M.-C.; St-Gelais, D.; Turgeon, S.L. Gel Formation and Rheological Properties of Fermented Milk with in situ Exopolysaccharide Production by Lactic Acid Bacteria. Dairy Sci. Technol. 2011, 91, 645. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.T.-B.; Nigen, M.; Jimenez, L.; Ait-Abderahim, H.; Cunault, C.; Marchesseau, S.; Picart-Palmade, L. A Multi-Scale Approach to Identify the Role of Heat Treatment, Milk Protein Composition and Starter Culture on the Gel Formation and the Texture Defects of Acid Milk Gel. Food Hydrocoll. 2018, 85, 299–310. [Google Scholar] [CrossRef]
- Brunauer, S.; Deming, L.S.; Deming, W.E.; Teller, E. On a Theory of the van Der Waals Adsorption of Gases. J. Am. Chem. Soc. 1940, 62, 1723–1732. [Google Scholar] [CrossRef]
- Andrade, R.D.; Lemus, R.; Perez, C.E. Models of Sorption Isotherms for Food: Uses and Limitations. Vitae 2011, 18, 325–334. [Google Scholar] [CrossRef]
- Caurie, M. Hysteresis Phenomenon in Foods. Int. J. Food Sci. Technol. 2007, 42, 45–49. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | ST-1G | ST-2E |
|---|---|---|
| EPS amount | ||
| tEPS [mg GE/L] | 659 ± 14 | 601 ± 67 |
| fEPS [mg GE/L] (% of tEPS) | 311 ± 29 (47%) | 540 ± 12 (90%) |
| cEPS [mg GE/L] (% of tEPS) | 348 (53%) | 61 (10%) |
| Biomass and Yield | ||
| X [g/L] | 4.31 ± 0.13 | 3.65 ± 0.06 |
| YEPS/X [mg/g] | 153 | 165 |
| YcEPS/X [mg/g] | 75 | 17 |
| Monosaccharides | ST-1G fEPS | ST-2E fEPS | Glycosidic Linkages | ST-1G fEPS | ST-2E fEPS |
|---|---|---|---|---|---|
| Glucose | 36.5 | 15.7 | t-Glcp | 21.0 | 21.0 |
| 1,3-Glcp | 6.2 | 4.1 | |||
| 1,4-Glcp | 2.1 | - | |||
| 1,3,6-Glcp | 7.6 | 3.1 | |||
| 1,6-Glcp | 4.2 | - | |||
| Galactose | 2.8 | 3.7 | t-Galf | - | 1.0 |
| t-Galp | - | 9.4 | |||
| Rhamnose | 43.8 | 64.1 | 1,2-Rhap | 37.8 | 15.3 |
| 1,2,3-Rhap | 8.3 | 15.8 | |||
| 1,2,3,4-Rhap | 12.8 | 22.1 | |||
| 1,3-Rhap | - | 8.4 | |||
| Glucosamine, N-Acetyl glucosamine | 16.9 | 16.5 | 1,3-GlcNp | - | - 2 |
| 1,4-GlcNp | - 2 | - 2 |
| Parameter | ST-1G fEPS | ST-2E fEPS |
|---|---|---|
| MW [106 Da] | 5.90 ± 0.11 | 12.66 ± 0.33 |
| Đ [-] | 5.0 ± 0.1 | 10.3 ± 0.4 |
| [η] [mL/mg] | 0.250 ± 0.012 | 0.269 ± 0.001 |
| RH [µm] | 61.5 ± 1.0 | 81.4 ± 0.1 |
| ST-1G | ST-2E | |||
|---|---|---|---|---|
| Cells + cEPS | fEPS | Cells + cEPS | fEPS | |
| X90 [g H2O/g dry matter] | 0.390 ± 0.003 | 0.420 ± 0.001 | 0.331 ± 0.013 | 0.357 ± 0.010 |
| Hysteresis area [area units] | 1.155 ± 0.069 | 1.667 ± 0.009 | 1.469 ± 0.079 | 1.394 ± 0.080 |
| GAB model parameters (adsorption) | ||||
| X0 [g/g] | 0.064 ± 0.001 | 0.083 ± 0.001 | 0.066 ± 0.003 | 0.057 ± 0.003 |
| CGAB [-] | 7.645 ± 0.188 | 8.519 ± 0.037 | 7.039 ± 0.073 | 8.441 ± 0.137 |
| kGAB [-] | 0.934 ± 0.002 | 0.894 ± 0.001 | 0.899 ± 0.002 | 0.947 ± 0.004 |
| GAB model parameters (desorption) | ||||
| X0 [g/g] | 0.063 ± 0.001 | 0.094 ± 0.001 | 0.071 ± 0.003 | 0.056 ± 0.006 |
| CGAB [-] | 25.163 ± 3.276 | 17.219 ± 0.284 | 16.834 ± 0.783 | 33.789 ± 12.919 |
| kGAB [-] | 0.941 ± 0.004 | 0.862 ± 0.001 | 0.883 ± 0.005 | 0.948 ± 0.010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nachtigall, C.; Surber, G.; Wefers, D.; Vogel, C.; Rohm, H.; Jaros, D. Capsular Exopolysaccharides from Two Streptococcus thermophilus Strains Differ in Their Moisture Sorption Behavior. Foods 2023, 12, 596. https://doi.org/10.3390/foods12030596
Nachtigall C, Surber G, Wefers D, Vogel C, Rohm H, Jaros D. Capsular Exopolysaccharides from Two Streptococcus thermophilus Strains Differ in Their Moisture Sorption Behavior. Foods. 2023; 12(3):596. https://doi.org/10.3390/foods12030596
Chicago/Turabian StyleNachtigall, Carsten, Georg Surber, Daniel Wefers, Cordula Vogel, Harald Rohm, and Doris Jaros. 2023. "Capsular Exopolysaccharides from Two Streptococcus thermophilus Strains Differ in Their Moisture Sorption Behavior" Foods 12, no. 3: 596. https://doi.org/10.3390/foods12030596