
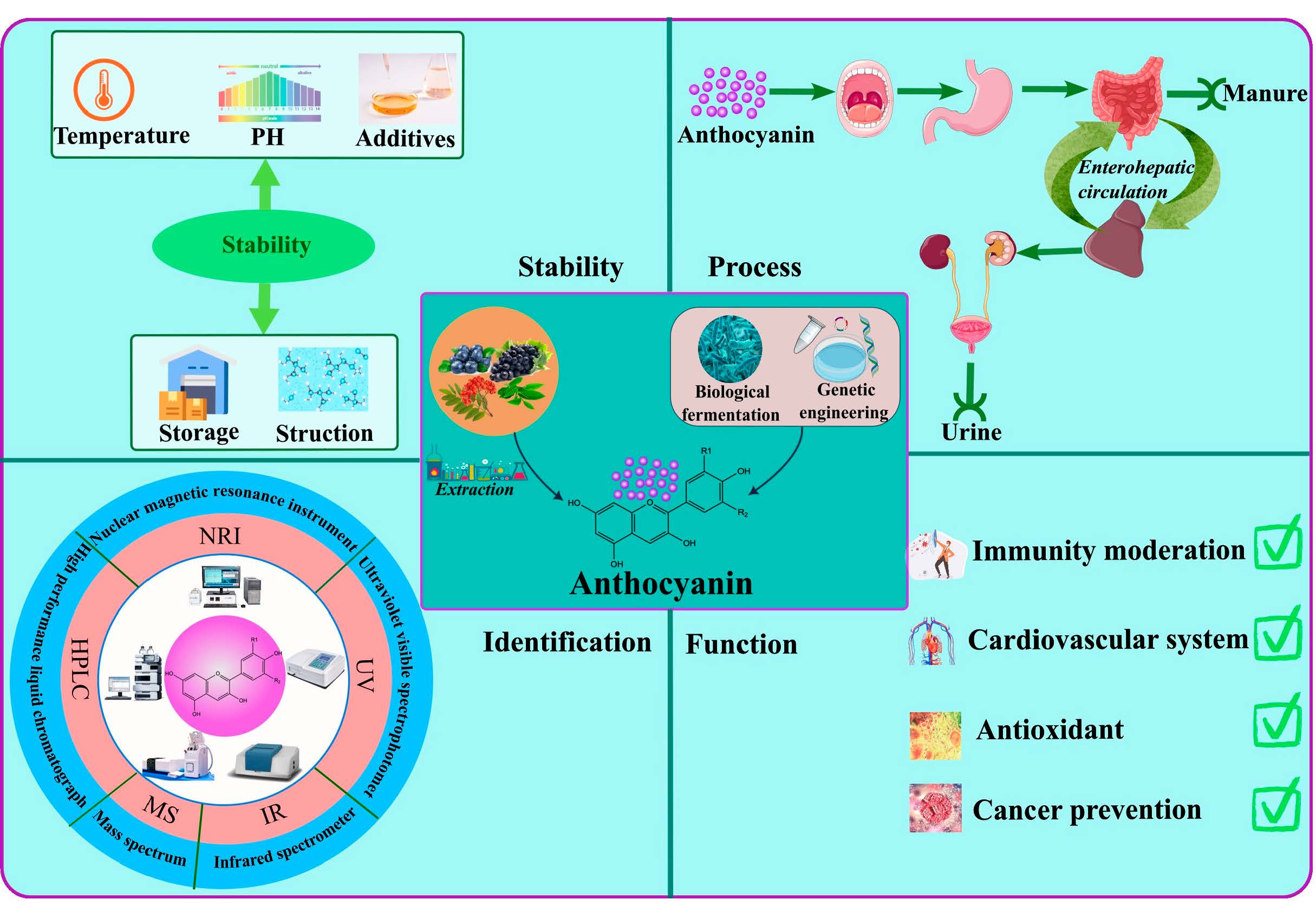
Advances in the Preparation, Stability, Metabolism, and Physiological Roles of Anthocyanins: A Review

Abstract
:
1. Introduction
2. Molecular Structure of Anthocyanins
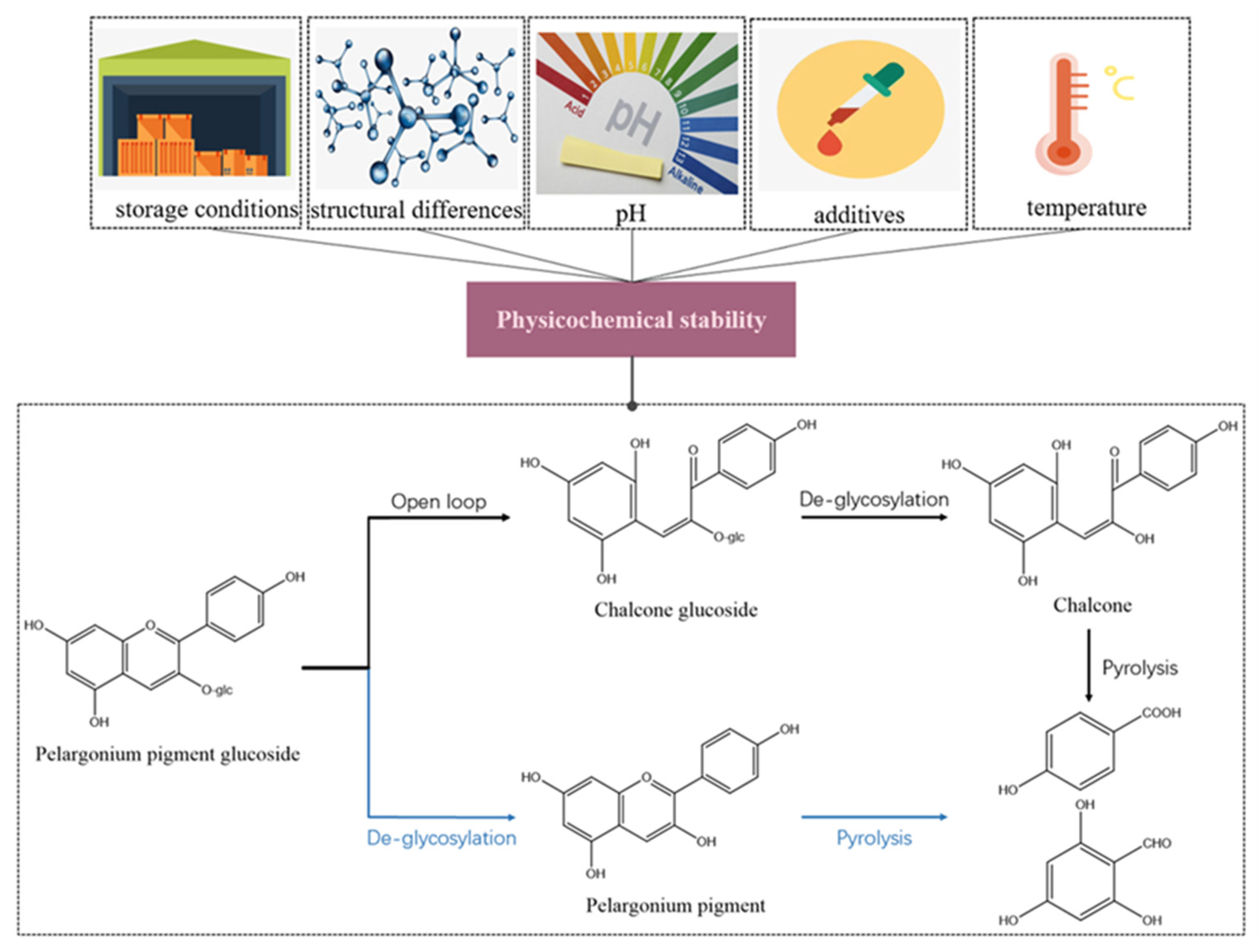
3. Physicochemical Properties of Anthocyanins
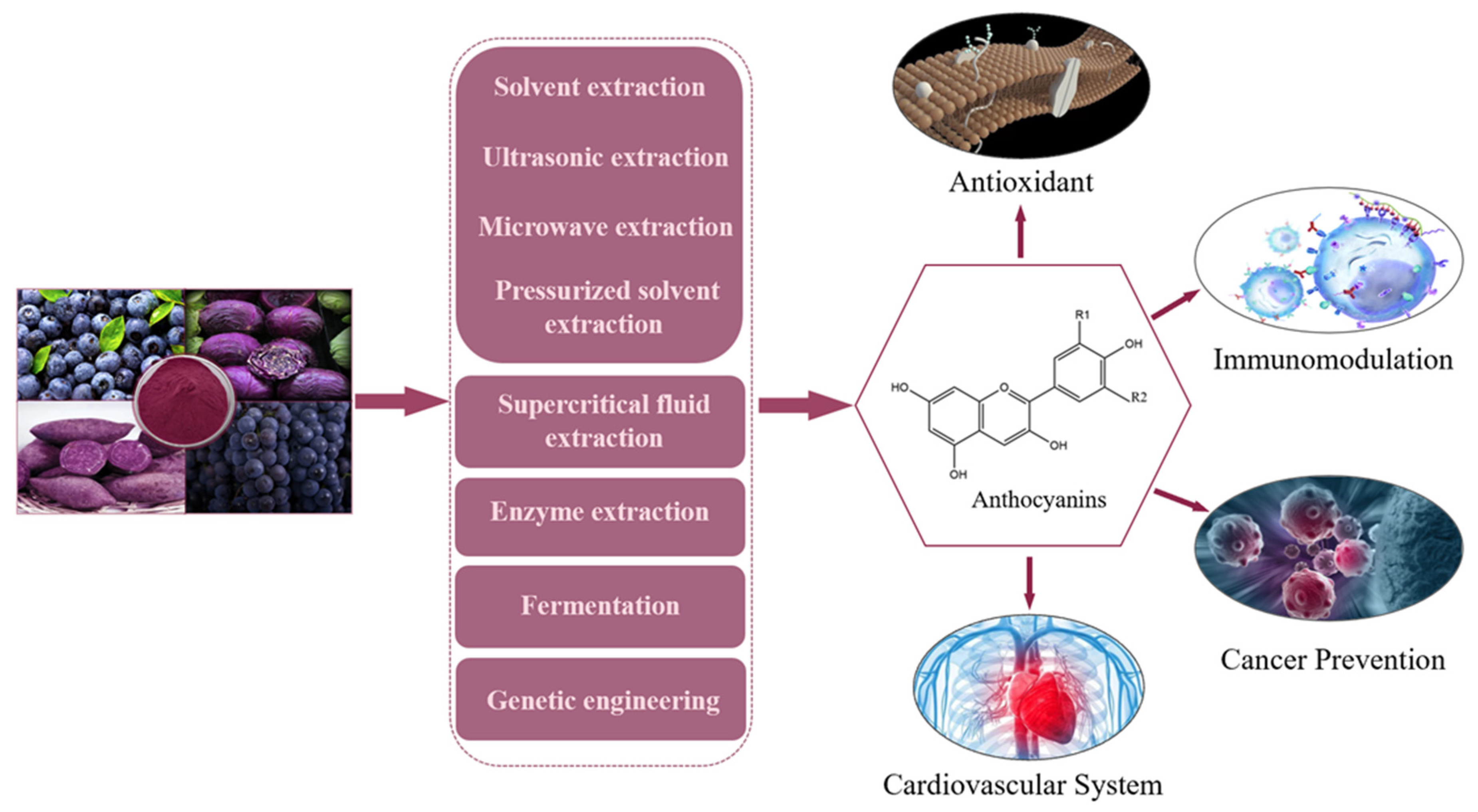
4. Extraction and Production of Anthocyanins
4.1. Solvent Extraction
4.2. Ultrasonic Extraction
4.3. Microwave Extraction
4.4. Supercritical Fluid Extraction Method
4.5. Pressurized Solvent Extraction
4.6. Enzyme Extraction
4.7. Fermentation
4.8. Genetic Engineering
5. Stability of Anthocyanins
5.1. Light Stability
5.2. Thermal Stability
6. Metabolism of Anthocyanins
6.1. Intake of Anthocyanins
6.2. Metabolism of Anthocyanins
6.3. Bioavailability of Anthocyanins
7. Physiological Functions of Anthocyanins
7.1. Antioxidant Effect of Anthocyanins
7.2. Immunomodulation by Anthocyanins
7.3. Cancer Prevention
7.4. Anthocyanins Regulate the Cardiovascular System
8. Encapsulation of Anthocyanins
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Escalante-Aburto, A.; Mendoza-Córdova, M.Y.; Mahady, G.B.; Luna-Vital, D.A.; Gutiérrez-Uribe, J.A.; Chuck-Hernández, C. Consumption of dietary anthocyanins and their association with a reduction in obesity biomarkers and the prevention of obesity. Trends Food Sci. Technol. 2023, 140, 104140. [Google Scholar] [CrossRef]
- Merecz-Sadowska, A.; Sitarek, P.; Kowalczyk, T.; Zajdel, K.; Jęcek, M.; Nowak, P.; Zajdel, R. Food anthocyanins: Malvidin and its glycosides as promising antioxidant and anti-inflammatory agents with potential health benefits. Nutrients 2023, 15, 3016. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, C.; Li, X.Y.; Wu, C.; Liu, C.; Xue, Z.H.; Kou, X.H. Investigation on the biological activity of anthocyanins and polyphenols in blueberry. J. Food Sci. 2021, 86, 614–627. [Google Scholar] [CrossRef]
- Im, Y.R.; Kim, I.; Lee, J. Phenolic composition and antioxidant activity of purple sweet potato (Ipomoea batatas (L.) Lam.): Varietal comparisons and physical distribution. Antioxidants 2021, 10, 462–478. [Google Scholar] [CrossRef] [PubMed]
- Deineka, V.I.; Oleinits, E.Y.; Blinova, I.P.; Deineka, L.A. Replacement of acetonitrile with ethanol in the determination of anthocyanins by reversed-phase high-performance liquid chromatography. J. Anal. Chem. 2023, 78, 63–67. [Google Scholar] [CrossRef]
- Oladzadabbasabadi, N.; Nafchi, A.M.; Ghasemlou, M.; Ariffin, F.; Singh, Z.; Al-Hassan, A.A. Natural anthocyanins: Sources, extraction, characterization, and suitability for smart packaging. Food Packag. Shelf 2022, 33, 100872. [Google Scholar] [CrossRef]
- Gérard, V.; Ay, E.; Morlet-Savary, F.; Graff, B.; Galopin, C.C.; Ogren, T.; Mutilangi, W.; Lalevée, J. Thermal and photochemical stability of anthocyanins from black carrot, grape juice and purple sweet potato in model beverages in presence of ascorbic acid. J. Agric. Food Chem. 2019, 67, 5647–5660. [Google Scholar] [CrossRef]
- Gui, H.; Sun, L.; Liu, R.; Si, X.; Li, D.; Wang, Y.; Shu, C.; Sun, X.; Jiang, Q.; Qiao, Y. Current knowledge of anthocyanin metabolism in the digestive tract: Absorption, distribution, degradation, and interconversion. Crit. Rev. Food Sci. Nutr. 2023, 63, 5953–5966. [Google Scholar] [CrossRef]
- Qi, Q.; Chu, M.; Yu, X.; Xie, Y.; Li, Y.; Du, Y.; Liu, X.; Zhang, Z.; Shi, J.; Yan, N. Anthocyanins and proanthocyanidins: Chemical structures, food sources, bioactivities, and product development. Food Rev. Int. 2023, 39, 4581–4609. [Google Scholar] [CrossRef]
- Mostafa, H.; Meroño, T.; Miñarro, A.; Sánchez-Pla, A.; Lanuza, F.; Zamora-Ros, R.; Rostgaard-Hansen, A.L.; Estanyol-Torres, N.; Cubedo-Culleré, M.; Tjønneland, A. Dietary sources of anthocyanins and their association with metabolome biomarkers and cardiometabolic risk factors in an observational study. Nutrients 2023, 15, 1208. [Google Scholar] [CrossRef]
- Mollaamin, F. Chemical structural and thermodynamic study of anthocyanins family in viewpoint of electron transfer. Biointerface Res. Appl. Chem. 2019, 9, 4349–4355. [Google Scholar]
- Espada-Bellido, E.; Ferreiro-González, M.; Carrera, C.; Palma, M.; Barbero, G.F. Optimization of the ultrasound-assisted extraction of anthocyanins and total phenolic compounds in mulberry (Morus nigra) pulp. Food Chem. 2017, 219, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Nie, Q.X.; Feng, L.; Hu, J.L.; Wang, S.N.; Chen, H.H. Effect of fermentation and sterilization on anthocyanins in blueberry. J. Sci. Food Agric. 2017, 97, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Ge, J.; Yue, P.; Yue, X.Y.; Fu, R.; Liang, J.; Gao, X. Loading of anthocyanins on chitosan nanoparticles influences anthocyanin degradation in gastrointestinal fluids and stability in a beverage. Food Chem. 2017, 221, 1671–1677. [Google Scholar] [CrossRef] [PubMed]
- Darniadi, S.; Ifie, I.; Ho, P.; Murray, B.S. Evaluation of total monomeric anthocyanin, total phenolic content and individual anthocyanins of foam-mat freeze-dried and spray-dried blueberry powder. J. Food Meas. Charact. 2019, 13, 1599–1606. [Google Scholar] [CrossRef]
- Fedenko, V.S.; Shemet, S.A.; Landi, M. Uv-vis spectroscopy and colorimetric models for detecting anthocyanin-metal complexes in plants: An overview of in vitro and in vivo techniques. J. Plant Physiol. 2017, 212, 13–28. [Google Scholar] [CrossRef]
- Stuppner, S.; Mayr, S.; Beganovic, A.; Bec, K.; Grabska, J.; Aufschnaiter, U.; Groeneveld, M.; Rainer, M.; Jakschitz, T.; Bonn, G.K.; et al. Near-infrared spectroscopy as a rapid screening method for the determination of total anthocyanin content in sambucus fructus. Sensors 2020, 20, 4983–4999. [Google Scholar] [CrossRef]
- Purwaningsih, I.; Fathiah, F.; Amaliyah, N.; Kuswiyanto, K. The phenolic, flavonoid, and anthocyanin content from methanol extract of senggani fruit and its antioxidant activity. Indones. J. Chem. Res. 2023, 10, 195–202. [Google Scholar] [CrossRef]
- Li, J.; Guo, X.; Wang, R.; Geng, Z.; Jia, J.; Pang, S.; Du, Y.; Jia, S.; Cui, J. Ultrasonic assisted extraction of anthocyanins from rose flower petal in des system and enzymatic acylation. LWT 2023, 180, 114693. [Google Scholar] [CrossRef]
- Rodsamran, P.; Sothornvit, R. Extraction of phenolic compounds from lime peel waste using ultrasonic-assisted and microwave-assisted extractions. Food Biosci. 2019, 28, 66–73. [Google Scholar] [CrossRef]
- Choommongkol, V.; Punturee, K.; Klumphu, P.; Rattanaburi, P.; Meepowpan, P.; Suttiarporn, P. Microwave-assisted extraction of anticancer flavonoid, 2′,4′-dihydroxy-6′-methoxy-3′,5′-dimethyl chalcone (dmc), rich extract from Syzygium nervosum fruits. Molecules 2022, 27, 1397–1411. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Dubey, K.K.; Marathe, S.J.; Singhal, R. Supercritical fluid extraction of bioactives from fruit waste and its therapeutic potential. Food Biosci. 2023, 52, 102418. [Google Scholar] [CrossRef]
- Espada-Bellido, E.; Ferreiro-Gonzalez, M.; Barbero, G.F.; Carrera, C.; Palma, M.; Barroso, C.G. Alternative extraction method of bioactive compounds from mulberry (Morus nigra L.) Pulp using pressurized-liquid extraction. Food Anal. Methods 2018, 11, 2384–2395. [Google Scholar] [CrossRef]
- Tan, J.; Li, Q.; Xue, H.; Tang, J. Ultrasound-assisted enzymatic extraction of anthocyanins from grape skins: Optimization, identification, and antitumor activity. J. Food Sci. 2020, 85, 3731–3744. [Google Scholar] [CrossRef]
- Jian, Z.; Ying, Z.; Matthew, M.; Jens, P.; Mamta, G.; Xia, W.; Sonya, C.; Koffas, M.A.G. Metabolic engineering of Corynebacterium glutamicum for anthocyanin production. Microb. Cell Factories 2018, 17, 143–156. [Google Scholar]
- Pereira, D.T.V.; Tarone, A.G.; Cazarin, C.B.B.; Barbero, G.F.; Martinez, J. Pressurized liquid extraction of bioactive compounds from grape marc. J. Food Eng. 2019, 240, 105–113. [Google Scholar] [CrossRef]
- Saad, K.R.; Parvatam, G.; Shetty, N.P. Medium composition potentially regulates the anthocyanin production from suspension culture of Daucus carota. 3 Biotech 2018, 8, 134–146. [Google Scholar] [CrossRef]
- Xie, L.; Su, H.; Sun, C.; Zheng, X.; Chen, W. Recent advances in understanding the anti-obesity activity of anthocyanins and their biosynthesis in microorganisms. Trends Food Ence Technol. 2017, 72, 13–24. [Google Scholar] [CrossRef]
- Peralta, K.D. Kinetics and modeling of cell growth for potential anthocyanin induction in cultures of Taraxacum officinale GH. Weber ex wiggers (dandelion) in vitro. Electron. J. Biotechnol. 2018, 36, 15–23. [Google Scholar]
- D’Amelia, V.; Villano, C.; Batelli, G.; Obanolu, Z.; Carputo, D. Genetic and epigenetic dynamics affecting anthocyanin biosynthesis in potato cell culture. Plant Sci. 2020, 298, 110597. [Google Scholar] [CrossRef]
- Belwal, T.; Singh, G.; Jeandet, P.; Pandey, A.; Luo, Z. Anthocyanins, multi-functional natural products of industrial relevance: Recent biotechnological advances. Biotechnol. Adv. 2020, 43, 107600. [Google Scholar] [CrossRef] [PubMed]
- Ingo, A.; Keim, W.V.A.; Micael, W.; Anne-Kathrine, H.E.; Julia, R.; Anne, O.; Stefan, M.; Hans-Peter, M.; Cathie, M.; Andrea, M. Colour bio-factories: Towards scale-up production of anthocyanins in plant cell cultures. Metab. Eng. 2018, 48, 218–232. [Google Scholar]
- Zhang, Y.; Luo, M.W.; Ma, X.H.; Hou, W.; Tang, H.R. Effects of sucrose and l-rhamnose on anthocyanin and related gene expression in rose. Fresenius Environ. Bull. 2019, 28, 2450–2456. [Google Scholar]
- Yunting, Z.; Leiyu, J.; Yali, L.; Qing, C.; Yuntian, Y.; Yong, Z.; Ya, L.; Bo, S.; Xiaorong, W.; Haoru, T. Effect of red and blue light on anthocyanin accumulation and differential gene expression in strawberry (Fragaria × ananassa). Molecules 2018, 23, 820–836. [Google Scholar]
- Wang, N.; Zhang, Z.; Jiang, S.; Xu, H.; Chen, X. Synergistic effects of light and temperature on anthocyanin biosynthesis in callus cultures of red-fleshed apple (Malus sieversii f. niedzwetzkyana). Plant Cell Tissue Organ Cult. 2016, 127, 217–227. [Google Scholar] [CrossRef]
- Zhou, X.R.; Xiao, Y.J.; Meng, X.H.; Liu, B.J. Full inhibition of whangkeumbae pear polyphenol oxidase enzymatic browning reaction by l-cysteine. Food Chem. 2018, 266, 1–8. [Google Scholar] [CrossRef]
- Pandey, R.P.; Parajuli, P.; Koffas, M.; Sohng, J.K. Microbial production of natural and non-natural flavonoids: Pathway engineering, directed evolution and systems/synthetic biology. Biotechnol. Adv. 2016, 34, 634–662. [Google Scholar] [CrossRef]
- Mark, L.; Constantinos, P.; Sascha, H.; De, G.P.A.; Jean-Marc, D.; Hall, R.D.; Stefan, M.; Jules, B. Engineering de novo anthocyanin production in saccharomyces cerevisiae. Microb. Cell Factories 2018, 17, 103–119. [Google Scholar]
- Cress, B.F.; Leitz, Q.D.; Kim, D.C.; Amore, T.D.; Koffas, M.A.G. Crispri-mediated metabolic engineering of e. Coli for o-methylated anthocyanin production. Microb. Cell Factories 2017, 16, 10–24. [Google Scholar] [CrossRef]
- Ying, L.; Yury, T.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Arnaud, B. Anthocyanin biosynthesis and degradation mechanisms in solanaceous vegetables: A review. Front. Chem. 2018, 6, 52–68. [Google Scholar]
- Zha, J.; Koffas, M. Production of anthocyanins in metabolically engineered microorganisms: Current status and perspectives. Synth. Syst. Biotechnol. 2017, 2, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.L.; Yu, Y.Q.; Chen, Z.J.; Wen, G.S.; Wei, F.G.; Zheng, Q.; Wang, C.D.; Xiao, X.L. Stability-increasing effects of anthocyanin glycosyl acylation. Food Chem. 2017, 214, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Pina, F. Chemical applications of anthocyanins and related compounds. A source of bio inspiration. J. Agric. Food Chem. 2014, 62, 6885–6897. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.O.; Freitas, A.A.; Ma Anita, A.L.; Quina, F.H. Chemistry and photochemistry of natural plant pigments: The anthocyanins. J. Phys. Org. Chem. 2016, 29, 594–599. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, Y.; Gao, Y. Effects of physical and chemical factors on anthocyanin stability in Lycoris sprengeri. J. Zhejiang A F Univ. 2021, 38, 587–596. [Google Scholar]
- Hu, N.; Zheng, J.; Li, W.; Suo, Y. Isolation, stability, and antioxidant activity of anthocyanins from Lycium ruthenicum murray and Nitraria Tangutorum bobr of qinghai-tibetan plateau. Sep. Sci. Technol. 2014, 49, 2897–2906. [Google Scholar] [CrossRef]
- Escher, G.B.; Wen, M.; Zhang, L.; Rosso, N.D.; Granato, D. Phenolic composition by uhplc-q-tof-ms/ms and stability of anthocyanins from Clitoria ternatea L. (Butterfly pea) blue petals. Food Chem. 2020, 331, 127341. [Google Scholar]
- Song, B.J.; Sapper, T.N.; Burtch, C.E.; Brimmer, K.; Goldschmidt, M.; Ferruzzi, M.G. Photo- and thermodegradation of anthocyanins from grape and purple sweet potato in model beverage systems. J. Agric. Food Chem. 2013, 61, 1364–1372. [Google Scholar] [CrossRef]
- Tachibana, N.; Kimura, Y.; Ohno, T. Examination of molecular mechanism for the enhanced thermal stability of anthocyanins by metal cations and polysaccharides. Food Chem. 2014, 143, 452–458. [Google Scholar] [CrossRef]
- Xu, K.; Zhang, M.; Fang, Z.; Wang, B. Degradation and regulation of edible flower pigments under thermal processing: A review. Crit. Rev. Food Sci. Nutr. 2020, 61, 1038–1048. [Google Scholar] [CrossRef]
- Connor, C.A.; Johnson, I.T.; Adriaens, M.; Belshaw, N.J.; Pierini, R. Procyanidin induces apoptosis of esophageal adenocarcinoma cells via jnk activation of c-jun. Nutr. Cancer Int. J. 2014, 66, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Cedo, L.; Castell-Auvi, A.; Pallares, V.; Blay, M.; Ardevol, A.; Arola, L.; Pinent, M. Grape seed procyanidin extract modulates proliferation and apoptosis of pancreatic beta-cells. Food Chem. 2013, 138, 524–530. [Google Scholar] [CrossRef]
- Deylami, M.Z.; Rahman, R.A.; Tan, C.P.; Bakar, J.; Olusegun, L. Effect of blanching on enzyme activity, color changes, anthocyanin stability and extractability of mangosteen pericarp: A kinetic study. J. Food Eng. 2016, 178, 12–19. [Google Scholar] [CrossRef]
- Burgos, G.; Amoros, W.; Muñoa, L.; Sosa, P.; Bonierbale, M. Total phenolic, total anthocyanin and phenolic acid concentrations and antioxidant activity of purple-fleshed potatoes as affected by boiling. J. Food Compos. Anal. 2013, 30, 6–12. [Google Scholar] [CrossRef]
- Kopjar, M.; Pilizota, V. Prevention of thermal degradation of anthocyanins in blackberry juice with addition of different sugars. Cyta J. Food 2011, 9, 237–242. [Google Scholar] [CrossRef]
- Wu, J.; Guan, Y.; Zhong, Q. Yeast mannoproteins improve thermal stability of anthocyanins at pH 7.0. Food Chem. 2015, 172, 121–128. [Google Scholar] [CrossRef]
- Han, F.; Yang, P.; Wang, H.; Fernandes, I.; Mateus, N.; Liu, Y. Digestion and absorption of red grape and wine anthocyanins through the gastrointestinal tract. Trends Food Sci. Technol. 2018, 83, 211–224. [Google Scholar] [CrossRef]
- Mallery, S.R.; Budendorf, D.E.; Larsen, M.P.; Pei, P.; Tong, M.; Holpuch, A.S.; Larsen, P.E.; Stoner, G.D.; Fields, H.W.; Chan, K.K. Effects of human oral mucosal tissue, saliva, and oral microflora on intraoral metabolism and bioactivation of black raspberry anthocyanins. Cancer Prev. Res. 2011, 4, 1209–1221. [Google Scholar] [CrossRef]
- Liu, W.L.; Fu, D.Y.; Zhang, X.J.; Chai, J.Y.; Tian, S.Y.; Han, J.Z. Development and validation of a new artificial gastric digestive system. Food Res. Int. 2019, 122, 183–190. [Google Scholar] [CrossRef]
- Oliveira, H.; Fernandes, I.; Brás, N.F.; Faria, A.; Freitas, V.D.; Calhau, C.; Mateus, N. Experimental and theoretical data on the mechanism by which red wine anthocyanins are transported through a human mkn-28 gastric cell model. J. Agric. Food Chem. 2015, 63, 7685–7692. [Google Scholar] [CrossRef]
- Allison, A.; Gregory, S.; Joshua, B.; Giusti, M. Time, concentration, and ph-dependent transport and uptake of anthocyanins in a human gastric epithelial (nci-n87) cell line. Int. J. Mol. Sci. 2017, 18, 446–457. [Google Scholar]
- Oliveira, H.; Wu, N.; Zhang, Q.; Wang, J.; Oliveira, J.; De Freitas, V.; Mateus, N.; He, J.; Fernandes, I. Bioavailability studies and anticancer properties of malvidin based anthocyanins, pyranoanthocyanins and non-oxonium derivatives. Food Funct. 2016, 7, 2462–2468. [Google Scholar] [CrossRef] [PubMed]
- Fawaz, A.; Hoi-Man, C.; Preedy, V.R.; Sharp, P.A.; Fran Ois, B. Regulation of glucose transporter expression in human intestinal caco-2 cells following exposure to an anthocyanin-rich berry extract. PLoS ONE 2013, 8, e78932. [Google Scholar]
- Baron, G.; Altomare, A.; Regazzoni, L.; Redaelli, V.; Grandi, S.; Riva, A.; Morazzoni, P.; Mazzolari, A.; Carini, M.; Vistoli, G. Pharmacokinetic profile of bilberry anthocyanins in rats and the role of glucose transporters: Lc–ms/ms and computational studies. J. Pharm. Biomed. Anal. 2017, 144, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Earling, M.; Beadle, T.; Niemeyer, E.D. Aai berry (Euterpe oleracea) dietary supplements: Variations in anthocyanin and flavonoid concentrations, phenolic contents, and antioxidant properties. Plant Foods Hum. Nutr. 2019, 74, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Ferrars, R.D.; Czank, C.; Zhang, Q.; Botting, N.P.; Kroon, P.A.; Cassidy, A.; Kay, C.D. The pharmacokinetics of anthocyanins and their metabolites in humans. Br. J. Pharmacol. 2014, 171, 3268–3282. [Google Scholar] [CrossRef]
- Eker, M.; Aaby, K.; Leto, I.B.; Teresa, S. A review of factors affecting anthocyanin bioavailability: Possible implications for the inter-individual variability. Multidiscip. Digit. Publ. Inst. 2019, 9, 2–19. [Google Scholar] [CrossRef]
- Lila, M.A.; Burton-Freeman, B.; Grace, M.; Kalt, W. Unraveling anthocyanin bioavailability for human health. Annu. Rev. Food Sci. Technol. 2016, 7, 375–393. [Google Scholar] [CrossRef]
- Wilhelmina, K.; Yan, L.; Jane, E.M.; Melinda, R.V.; Sherry, A.E.F. Anthocyanin metabolites are abundant and persistent in human urine. J. Agric. Food Chem. 2014, 62, 3926–3934. [Google Scholar]
- Chen, Y.; Chen, H.; Zhang, W.J.; Ding, Y.Y.; Zhao, T.; Zhang, M.; Mao, G.H.; Feng, W.W.; Wu, X.Y.; Yang, L.Q. Bioaccessibility and biotransformation of anthocyanin monomers following in vitro simulated gastric-intestinal digestion and in vivo metabolism in rats. Food Funct. 2019, 10, 6052–6061. [Google Scholar] [CrossRef]
- Braga, A.; Murador, D.C.; Mendes, D.S.M.L.; De Rosso, V.V. Bioavailability of anthocyanins: Gaps in knowledge, challenges and future research. J. Food Compos. Anal. 2017, 68, 31–40. [Google Scholar] [CrossRef]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.H.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black currant anthocyanins attenuate weight gain and improve glucose metabolism in diet-induced obese mice with intact, but not disrupted, gut microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef] [PubMed]
- Krga, I.; Milenkovic, D. Anthocyanins: From sources and bioavailability to cardiovascular-health benefits and molecular mechanisms of action. J. Agric. Food Chem. 2019, 67, 1771–1783. [Google Scholar] [CrossRef] [PubMed]
- Fang, J. Some anthocyanins could be efficiently absorbed across the gastrointestinal mucosa: Extensive presystemic metabolism reduces apparent bioavailability. J. Agric. Food Chem. 2014, 62, 3904–3911. [Google Scholar] [CrossRef] [PubMed]
- Wiczkowski, W.; Szawara-Nowak, D.; Romaszko, J. The impact of red cabbage fermentation on bioavailability of anthocyanins and antioxidant capacity of human plasma. Food Chem. 2016, 190, 730–740. [Google Scholar] [CrossRef]
- Court, M.H. Interindividual variability in hepatic drug glucuronidation: Studies into the role of age, sex, enzyme inducers, and genetic polymorphism using the human liver bank as a model system. Drug Metab. Rev. 2010, 42, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.R.; Liu, X.M.; Chen, Z.Y.; Zhang, Y.S.; Zhang, Y.H. Mulberry anthocyanin biotransformation by intestinal probiotics. Food Chem. 2016, 213, 721–727. [Google Scholar] [CrossRef]
- Tena, N.; Martín, J.; Asuero, A.G. State of the art of anthocyanins: Antioxidant activity, sources, bioavailability, and therapeutic effect in human health. Antioxidants 2020, 9, 451–478. [Google Scholar] [CrossRef]
- Ma, Y.; Feng, Y.; Diao, T.; Zeng, W.; Zuo, Y. Experimental and theoretical study on antioxidant activity of the four anthocyanins. J. Mol. Struct. 2020, 1204, 127509. [Google Scholar] [CrossRef]
- Ali, H.M.; Almagribi, W.; Al-Rashidi, M.N. Antiradical and reductant activities of anthocyanidins and anthocyanins, structure-activity relationship and synthesis. Food Chem. 2016, 194, 1275–1282. [Google Scholar] [CrossRef]
- Blando, F.; Calabriso, N.; Berland, H.; Maiorano, G.; Gerardi, C.; Carluccio, M.A.; Andersen, Y.M. Radical scavenging and biological activities of 2 representative anthocyanin groupings from 3 pigment-rich fruits and vegetables. Preprints 2018, 19, 196–213. [Google Scholar]
- Li, X.; Zhu, F.; Zeng, Z. Effects of different extraction methods on antioxidant properties of blueberry anthocyanins. Open Chem. 2021, 19, 138–148. [Google Scholar] [CrossRef]
- Shi, S.; Lv, M.; Jin, L.; Qin, G.; Gao, Y.; Ji, J.; Hao, L. Antioxidant properties of anthocyanin revealed through the hydrogen atom transfer: Combined effects of temperature and pH. Mol. Phys. 2021, 119, e1936246. [Google Scholar] [CrossRef]
- Triebel, S.; Trieu, H.L.; Richling, E. Modulation of inflammatory gene expression by a bilberry (Vaccinium myrtillus L.) Extract and single anthocyanins considering their limited stability under cell culture conditions. J. Agric. Food Chem. 2012, 60, 8902–8910. [Google Scholar] [CrossRef]
- Espley, R.V.; Butts, C.A.; Laing, W.A.; Martell, S.; Smith, H.; Mcghie, T.K.; Zhang, J.; Paturi, G.; Hedderley, D.; Bovy, A. Dietary flavonoids from modified apple reduce inflammation markers and modulate gut microbiota in mice. J. Nutr. 2014, 144, 146–154. [Google Scholar] [CrossRef]
- Clark, I.A.; Vissel, B. Broader insights into understanding tumor necrosis factor and neurodegenerative disease pathogenesis infer new therapeutic approaches. J. Alzheimer’s Dis. 2021, 79, 931–948. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, N.; Zhang, Z.T.; Jing, P. Effects of dietary cyanidin-3-diglucoside-5-glucoside complexes with rutin/mg(ii) against h2o2-induced cellular oxidative stress. Food Res. Int. 2019, 126, 108591. [Google Scholar] [CrossRef]
- Jaiswal, V.; Park, M.; Lee, H.J. Comparative transcriptome analysis of the expression of antioxidant and immunity genes in the spleen of a cyanidin 3-o-glucoside-treated alzheimer’s mouse model. Antioxidants 2021, 10, 1435–1450. [Google Scholar] [CrossRef]
- Hurst, R.D.; Lyall, K.A.; Roberts, J.M.; Perthaner, A.; Wells, R.W.; Cooney, J.M.; Jensen, D.J.; Burr, N.S.; Hurst, S.M. Consumption of an anthocyanin-rich extract made from new zealand blackcurrants prior to exercise may assist recovery from oxidative stress and maintains circulating neutrophil function: A pilot study. Front. Nutr. 2019, 6, 73–86. [Google Scholar] [CrossRef]
- Hui, T.; Fang, T.; Lin, Q.; Song, H.; Liu, B.; Lei, C. Red raspberry and its anthocyanins: Bioactivity beyond antioxidant capacity. Trends Food Sci. Technol. 2017, 66, 153–165. [Google Scholar]
- Hee, J.; Sunga, C.; Yu, L.; Eun, L.; Myoung, P.; Kyu, P.; Cuk-Seong, K.; Yong, L.; Jong-Tae, P.; Byeong, J. Anthocyanin-rich extract from red chinese cabbage alleviates vascular inflammation in endothelial cells and apo e−/− mice. Int. J. Mol. Sci. 2018, 19, 816–832. [Google Scholar]
- Grunt, T.W.; Heller, G. A critical appraisal of the relative contribution of tissue architecture, genetics, epigenetics and cell metabolism to carcinogenesis. Prog. Biophys. Mol. Biol. 2023, 182, 26–33. [Google Scholar] [CrossRef]
- Caldern-Reyes, C.; Pezoa, R.S.; Leal, P.; Ribera-Fonseca, A.; Cceres, C.; Riquelme, I.; Zambrano, T.; Pea, D.; Alberdi, M.; Reyes-Daz, M. Anthocyanin-rich extracts of calafate (Berberis microphylla G. Forst.) Fruits decrease in vitro viability and migration of human gastric and gallbladder cancer cell lines. J. Soil Sci. Plant Nutr. 2020, 20, 1891–1903. [Google Scholar] [CrossRef]
- Marty, R.; Kaabinejadian, S.; Rossell, D.; Slifker, M.J.; Joris, V.D.H.; Engin, H.B.; De Prisco, N.; Ideker, T.; Hildebrand, W.H.; Font-Burgada, J. Mhc-i genotype restricts the oncogenic mutational landscape. Cell 2017, 171, 1272–1283. [Google Scholar] [CrossRef]
- Peiffer, D.S.; Zimmerman, N.P.; Wang, L.S.; Ransom, B.W.S.; Carmella, S.G.; Kuo, C.T.; Siddiqui, J.; Chen, J.H.; Oshima, K.; Huang, Y.W. Chemoprevention of esophageal cancer with black raspberries, their component anthocyanins, and a major anthocyanin metabolite, protocatechuic acid. Cancer Prev. Res. 2014, 7, 574–584. [Google Scholar] [CrossRef]
- Lippert, E.; Ruemmele, P.; Obermeier, F.; Goelder, S.; Endlicher, E. Anthocyanins prevent colorectal cancer development in a mouse model. Digestion 2017, 95, 275–280. [Google Scholar] [CrossRef]
- Golemis, E.A.; Paul, S.; Beck, T.N.; Ed, S.; Hunter, D.J.; Ernest, H.; Nancy, H. Molecular mechanisms of the preventable causes of cancer in the United States. Genes Dev. 2018, 32, 868–902. [Google Scholar] [CrossRef]
- Luo, L.P.; Han, B.; Yu, X.P.; Chen, X.Y.; Zhou, J.; Chen, W.; Zhu, Y.F.; Peng, X.L.; Zou, Q.; Li, S.Y. Anti-metastasis activity of black rice anthocyanins against breast cancer: Analyses using an erbb2 positive breast cancer cell line and tumoral xenograft model. Asian Pac. J. Cancer Prev. APJCP 2014, 15, 6219–6225. [Google Scholar] [CrossRef]
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Muntner, P. Heart disease and stroke statistics—2017 update: A report from the american heart association. Circulation 2017, 131, e29. [Google Scholar] [CrossRef]
- Ziff, O.J.; Kotecha, D. Digoxin: The good and the bad. Trends Cardiovasc. Med. 2016, 26, 585–595. [Google Scholar] [CrossRef]
- Giudicessi, J.R.; Ackerman, M.J.; Camilleri, M. Cardiovascular safety of prokinetic agents: A focus on drug-induced arrhythmias. Neurogastroenterol. Motil. 2018, 30, e13302. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.D.; Luo, M.; Shang, A.; Mao, Q.Q.; Li, B.Y.; Gan, R.Y.; Li, H.B. Antioxidant food components for the prevention and treatment of cardiovascular diseases: Effects, mechanisms, and clinical studies. Oxid. Med. Cell. Longev. 2021, 2021, 6627355. [Google Scholar] [CrossRef] [PubMed]
- Aboonabi, A.; Aboonabi, A. Anthocyanins reduce inflammation and improve glucose and lipid metabolism associated with inhibiting nuclear factor-kappab activation and increasing ppar-γ gene expression in metabolic syndrome subjects. Free Radic. Biol. Med. 2020, 150, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Yang, J.; Yu, W.; Wei, J. Anthocyanins from black chokeberry delayed ageing-related degenerative changes in the heart. Indian J. Pharm. Educ. Res. 2019, 53, 112–116. [Google Scholar] [CrossRef]
- Wang, Z.; Pang, W.; He, C.; Li, Y.; Jiang, Y.; Guo, C. Blueberry anthocyanin-enriched extracts attenuate fine particulate matter (pm2.5)-induced cardiovascular dysfunction. J. Agric. Food Chem 2016, 65, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Cutler, B.R.; Petersen, C.; Anandh Babu, P.V. Mechanistic insights into the vascular effects of blueberries: Evidence from recent studies. Mol. Nutr. Food Res. 2017, 61, 1600271. [Google Scholar] [CrossRef]
- Liu, R.; Wang, X.; Yang, L.; Wang, Y.; Gao, X. Coordinated encapsulation by β-cyclodextrin and chitosan derivatives improves the stability of anthocyanins. Int. J. Biol. Macromol. 2023, 242, 125060. [Google Scholar] [CrossRef]
- Svanberg, L.; Malmberg, K.; Gustinelli, G.; Öhgren, C.; Persson, I.; Brive, L.; Wassén, S. Effect of anthocyanins on lipid oxidation and microbial spoilage in value-added emulsions with bilberry seed oil, anthocyanins and cold set whey protein hydrogels. Food Chem. 2019, 272, 273–278. [Google Scholar] [CrossRef]
- Nazareth, M.S.; Shreelakshmi, S.V.; Rao, P.J.; Shetty, N.P. Micro and nanoemulsions of Carissa spinarum fruit polyphenols, enhances anthocyanin stability and anti-quorum sensing activity: Comparison of degradation kinetics. Food Chem. 2021, 359, 129876. [Google Scholar] [CrossRef]
- Rabelo, C.A.; Taarji, N.; Khalid, N.; Kobayashi, I.; Nakajima, M.; Neves, M.A. Formulation and characterization of water-in-oil nanoemulsions loaded with açaí berry anthocyanins: Insights of degradation kinetics and stability evaluation of anthocyanins and nanoemulsions. Food Res. Int. 2018, 106, 542–548. [Google Scholar] [CrossRef]
- Li, J.; Guo, C.; Cai, S.; Yi, J.; Zhou, L. Fabrication of anthocyanin–rich w1/o/w2 emulsion gels based on pectin–gdl complexes: 3d printing performance. Food Res. Int. 2023, 168, 112782. [Google Scholar] [CrossRef]
- Ozcan, B.E.; Saroglu, O.; Karakas, C.Y.; Karadag, A. Encapsulation of purple basil leaf extract by electrospraying in double emulsion (w/o/w) filled alginate-carrageenan beads to improve the bioaccessibility of anthocyanins. Int. J. Biol. Macromol. 2023, 250, 126207. [Google Scholar] [CrossRef] [PubMed]
- Ju, M.; Zhu, G.; Huang, G.; Shen, X.; Zhang, Y.; Jiang, L.; Sui, X. A novel pickering emulsion produced using soy protein-anthocyanin complex nanoparticles. Food Hydrocolloid. 2020, 99, 105329. [Google Scholar] [CrossRef]
- Ge, J.; Yue, X.; Wang, S.; Chi, J.; Liang, J.; Sun, Y.; Gao, X.; Yue, P. Nanocomplexes composed of chitosan derivatives and β-lactoglobulin as a carrier for anthocyanins: Preparation, stability and bioavailability in vitro. Food Res. Int. 2019, 116, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Ravanfar, R.; Tamaddon, A.M.; Niakousari, M.; Moein, M.R. Preservation of anthocyanins in solid lipid nanoparticles: Optimization of a microemulsion dilution method using the placket–burman and box–behnken designs. Food Chem. 2016, 199, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Hao, J.; Wang, Q.; Li, X.; Xu, D. Extraction of structurally intact and well-stabilized rice bran oil bodies as natural pre-emulsified o/w emulsions and investigation of their rheological properties and components interaction. Food Res. Int. 2023, 164, 112457. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Raw Material | Advantages and Disadvantages | References |
|---|---|---|---|
| Solvent extraction | Senggani fruit | minimize damage or loss of compounds; low yield, low product quality, high energy input, time-consuming, need substantial amounts of solvents | [18] |
| Ultrasonic extraction | rose flower petal | significantly improve anthocyanins extraction rate, efficient; require complex and expensive equipment, difficult to realize industrialization | [19,20] |
| Microwave extraction | the fruits of Syzygium nervosum (ma-kiang in Thai) | green, rapid extraction time, higher extraction rate, with less organic solvent, wastewater removal, and energy use | [20,21] |
| Supercritical fluid extraction method | fruit wastes | green, reduced cost of operation; high cost, have limits to realize industrialization | [22] |
| Pressurized solvent extraction | black mulberry (Morus nigra L.) pulp | require less methanol consumption, higher extraction efficiency | [23] |
| Enzyme extraction | grape skins | enhance the yield, effective and environmentally friendly | [24] |
| Genetic engineering | Corynebacterium glutamicum | a sustainable method to produce anthocyanins, ease of cultivation and fast growth, availability of sophisticated genetic tools, and well-defined metabolic networks and models; the heterologous expression of plant-derived genes, optimal expression depends on the host strains | [25] |
| Type | Composition | Property | Function | References |
|---|---|---|---|---|
| Nanoparticle | β-cyclodextrin, chitosan hydrochloride, and carboxymethyl chitosan | particle size, 333.86 nm; zeta-potential, +45.97 mV | Improve the stability of anthocyanins, light irradiation for 12 days, the retention rate of encapsulated anthocyanin and free anthocyanin was 77.6% and 58.1% | [107] |
| Emulsion | bilberry seed oil | particle size, 1.7 nm; zeta-potential, −71.45 mV; isoelectric point, pH = 4.0 | Display a protective effect against lipid oxidation | [108] |
| Nanoemulsion | organic phase (ethanol), aqueous phase (glycerol-sodium benzoate buffer solution) | particle size, 20–500 nm | Enhance the bioavailability and absorption, kinetically stable systems; thermal processing at 90 °C for 3 min, the retention of anthocyanins was 72.24% | [109] |
| Microemulsion | organic phase (ethanol), aqueous phase (glycerol-sodium benzoate buffer solution), and surfactant | particle size, 10–100 nm | Enhance the bioavailability and absorption, thermodynamically stable isotropic liquids; thermal processing at 90 °C for 3 min, the retention of anthocyanins was around 54% | [109] |
| Water-in-oil (W/O) nanoemulsion | a continuous oil phase, the dispersed phase, and an emulsifier | particle size, 131.5 nm to 195.3 nm; polydispersity (after 30 days of storage), 0.2 to 0.6 | No phase separation after 30 days of storage, the samples had decreased concentrations of polyphenols after 30 days of storage at 4 °C, the highest retention was 94.6% | [110] |
| Water-in-oil-in-water (W1/O/W2) double emulsion | oil phase, the internal aqueous phase (W1), and the aqueous phase (W2) | particle size, 625 nm; zeta-potential, −48 mV; viscosity, 29.1 Pa·s | Improve the stability of anthocyanins, 28 days of storage at 4 °C and 25 °C, the retention rate of encapsulated anthocyanin were 93.2% and 88.9% | [111,112] |
| Pickering emulsion | soy protein isolate, anthocyanins | particles size, 186 nm to 675 nm | Notable improve radical scavenging activity and improved antioxidant capacity, the FFA releasing rate could be retarded by up to 6.9% | [113] |
| Nanocomplexes | chitosan hydrochloride, carboxymethyl chitosan, and β-Lactoglobulin | particles size, 91.71 nm; encapsulation efficiency, 69.33% | Improve the stability and bioavailability, anthocyanins retention rate was 68.9% | [114] |
| Chitosan nanoparticle | carboxymethyl chitosan, chitosan hydrochloride | particles size, 219.53 nm; encapsulation efficiency, 63.15% | Improve the stability of anthocyanins, after 35 days at 4 °C, the retention rate of encapsulated anthocyanin and free anthocyanin were 84.5% and 71.2% | [14] |
| Solid lipid nanoparticle | the lipid phase (palmitic acid, Span 85, and egg lecithin), cosurfactant (ethanol or isobutanol), and distilled water | particle size, 455 ± 2 nm; entrapment efficiency 89.2 ± 0.3% | Improve the stability of anthocyanins and enhances the oral bioavailability of anthocyanins | [115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Zhang, F.; Wang, Z.; Feng, Y.; Han, Y. Advances in the Preparation, Stability, Metabolism, and Physiological Roles of Anthocyanins: A Review. Foods 2023, 12, 3969. https://doi.org/10.3390/foods12213969
Li Q, Zhang F, Wang Z, Feng Y, Han Y. Advances in the Preparation, Stability, Metabolism, and Physiological Roles of Anthocyanins: A Review. Foods. 2023; 12(21):3969. https://doi.org/10.3390/foods12213969
Chicago/Turabian StyleLi, Qi, Fengzhen Zhang, Zhenzhen Wang, Yaoze Feng, and Yahong Han. 2023. "Advances in the Preparation, Stability, Metabolism, and Physiological Roles of Anthocyanins: A Review" Foods 12, no. 21: 3969. https://doi.org/10.3390/foods12213969