
The Impact of Sea Urchin as an Ingredient on the Physicochemical, Microbiological, and Sensory Properties of Fish Sauce Fermentation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials
2.2. Experimental Fish Sauce
2.3. Determination of pH and Water Activity (aw)
2.4. Determination of Total Volatile Bases (TVB-N) and Trimethylamine (TMA-N) Contents
2.5. Protease Activity Measurements
2.6. Determination of the Total Nitrogen Contents
2.7. Determination of Formaldehyde and Amino Nitrogen Contents
2.8. Determination of the Salt Content
2.9. Microbiological Analyses
2.10. Sensory Evaluation
2.11. Statistical Analysis
3. Results and Discussion
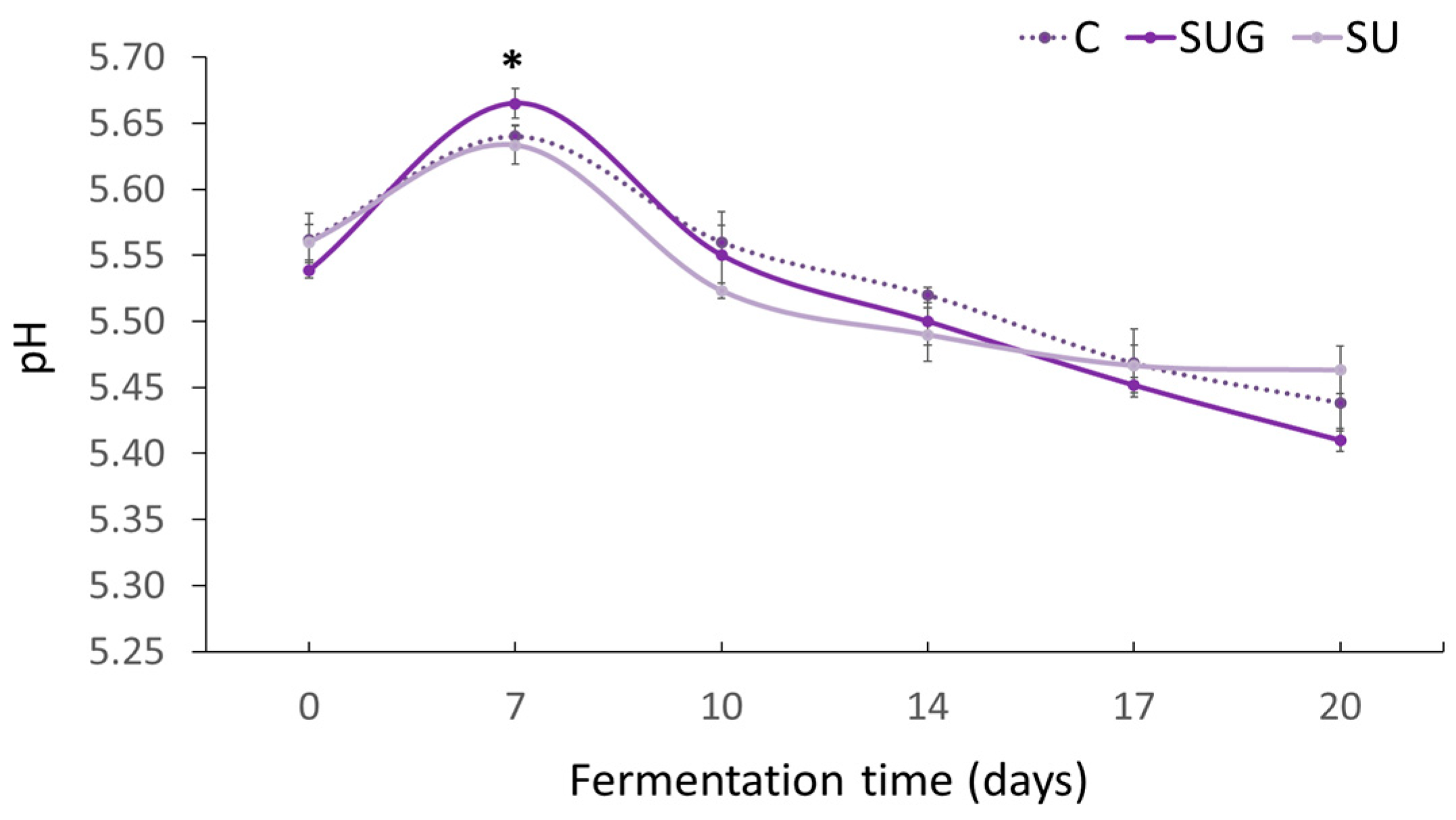
3.1. Evolution of pH
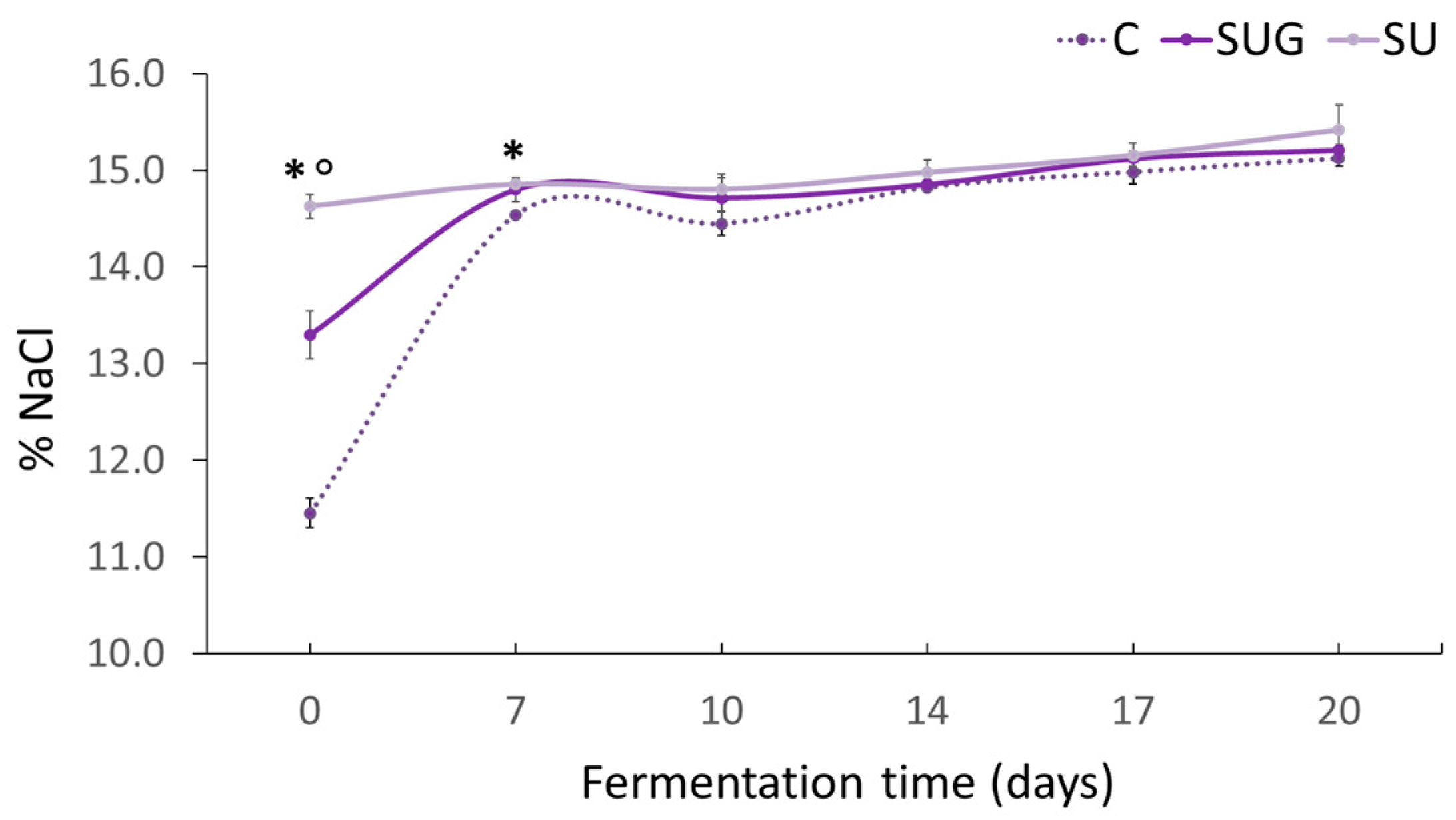
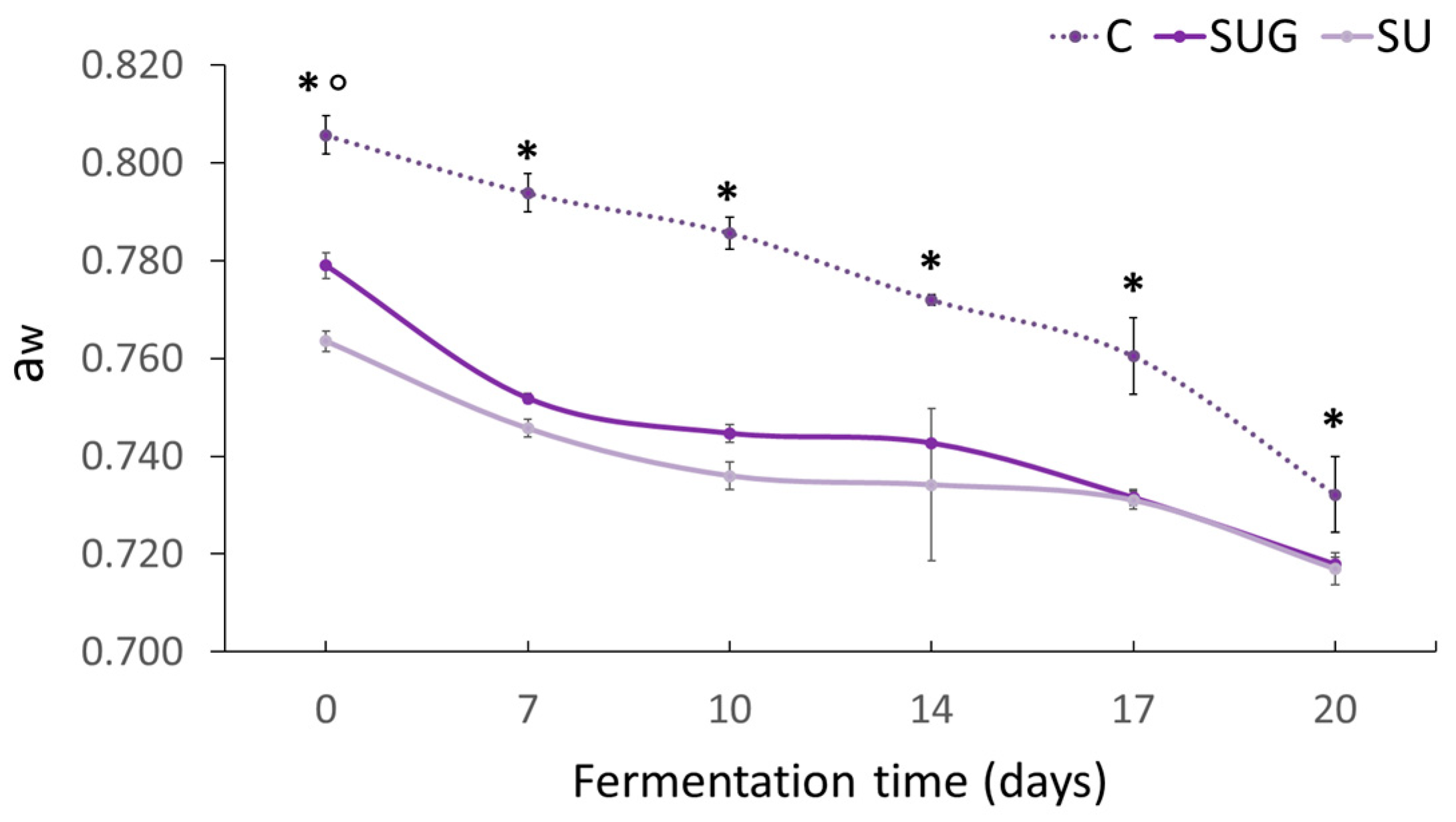
3.2. Evolution of the Salt Content (%NaCl) and Water Activity (aw)
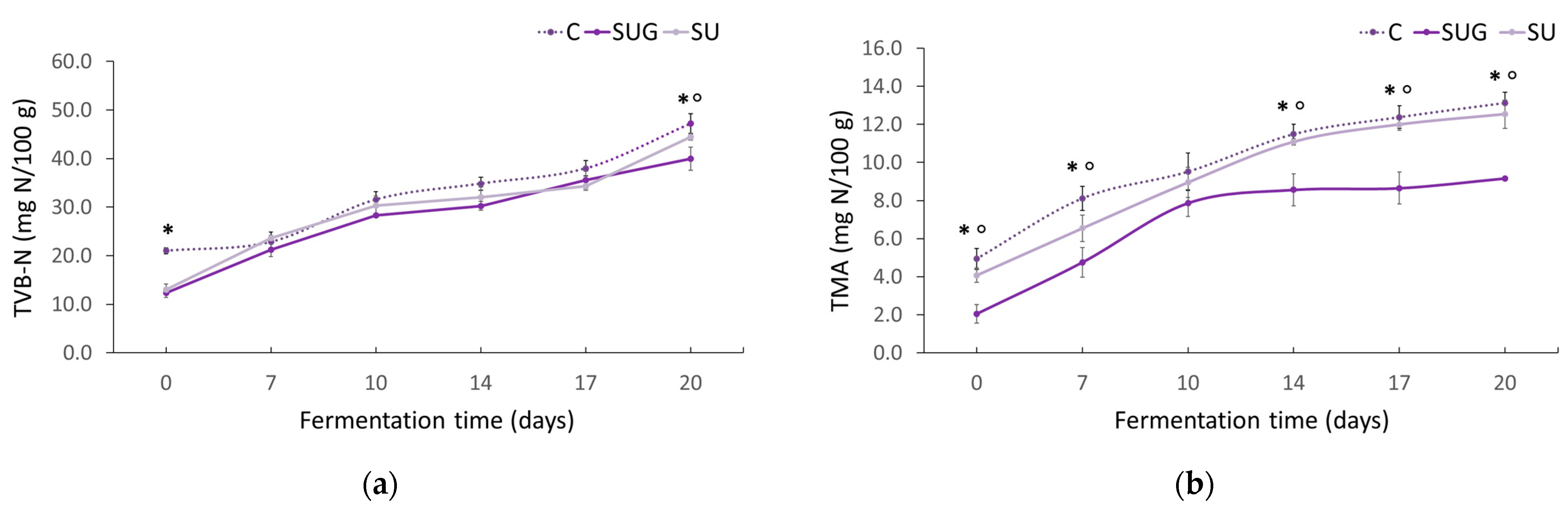
3.3. Evolution of the Total Volatile Bases (TVB-N) and Trimethylamine (TMA-N) Contents
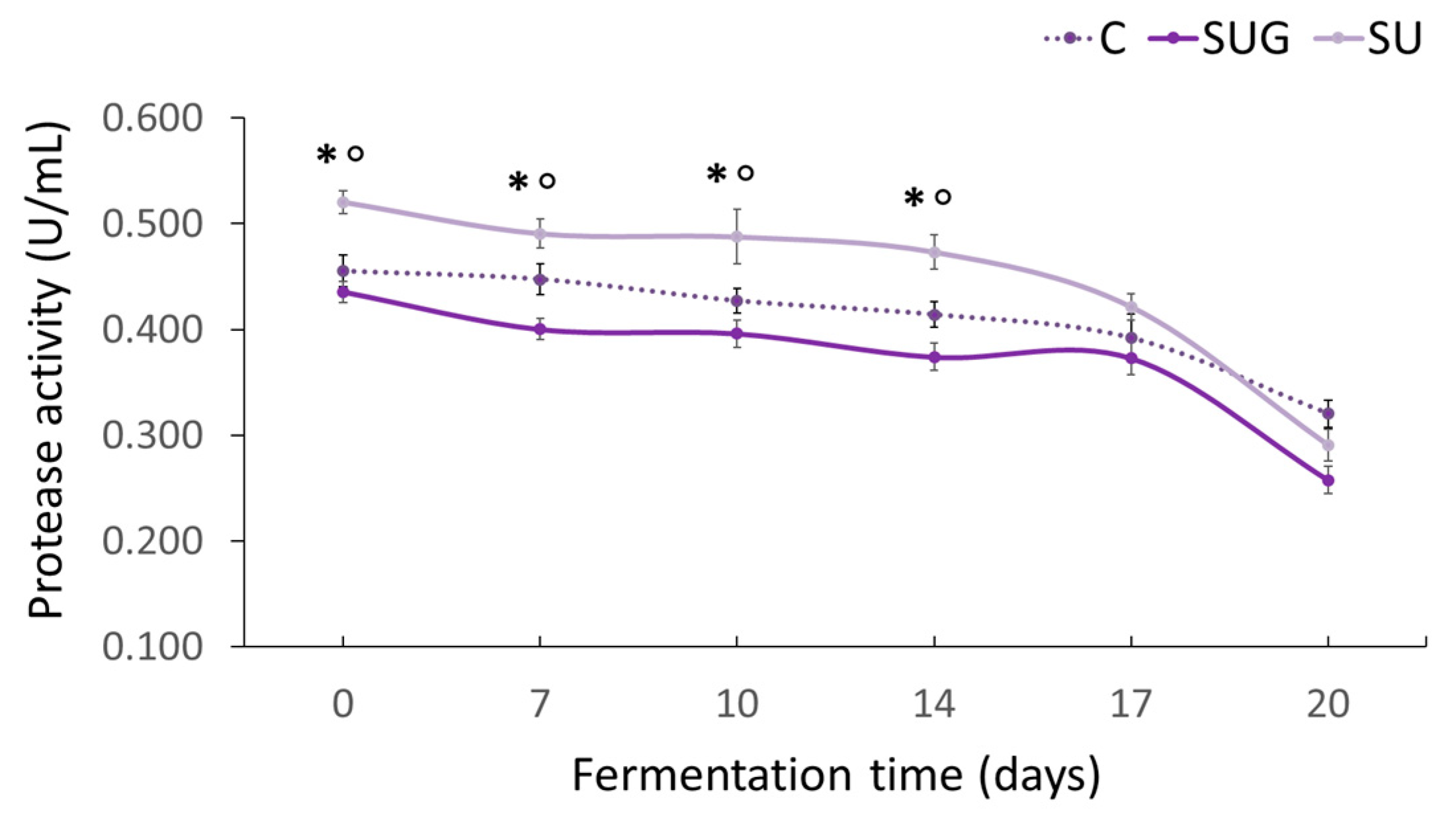
3.4. Evolution of the Protease Activity
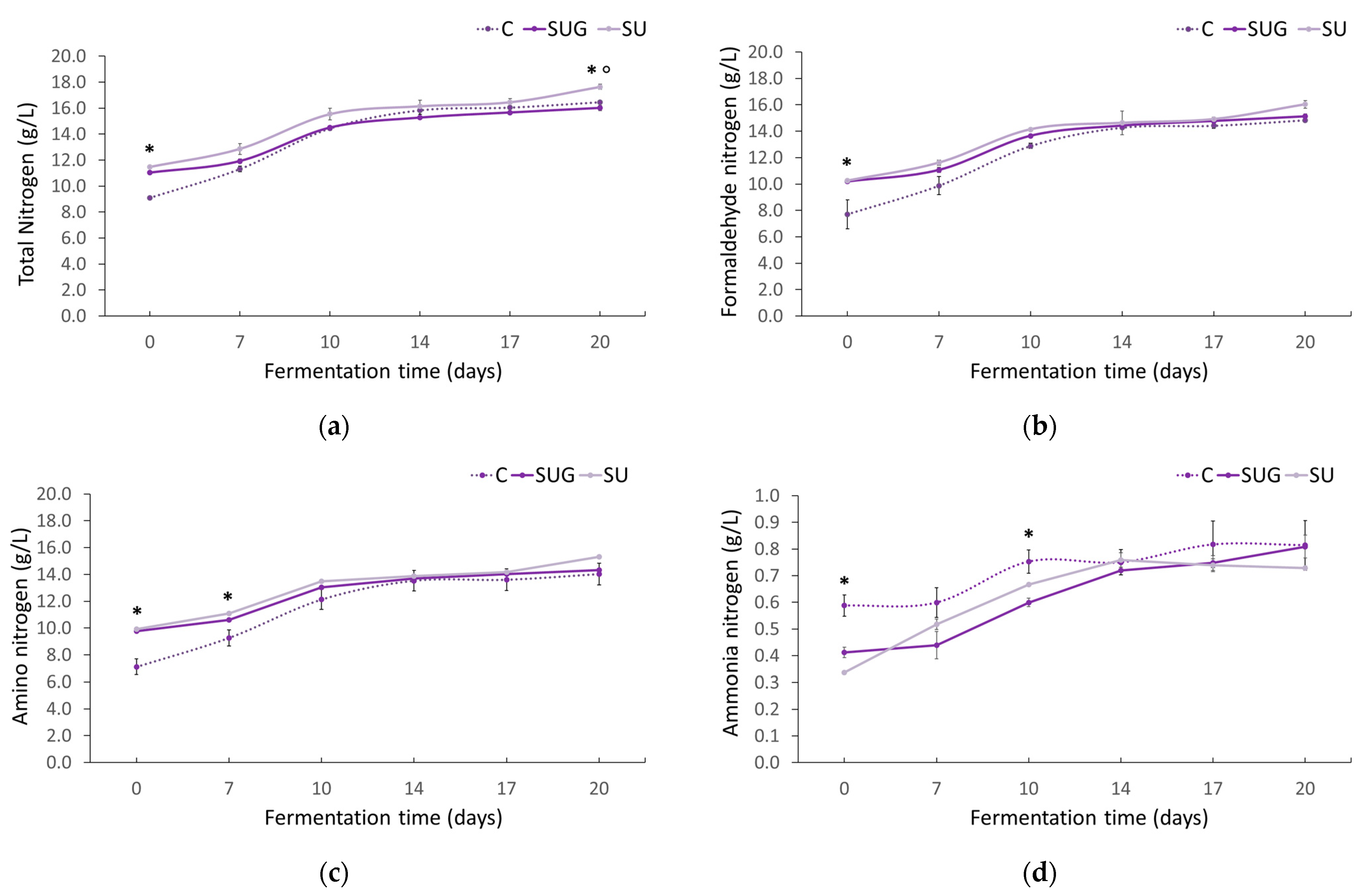
3.5. Evolution of the Total Nitrogen, Formaldehyde Nitrogen, and Ammonia Nitrogen Contents
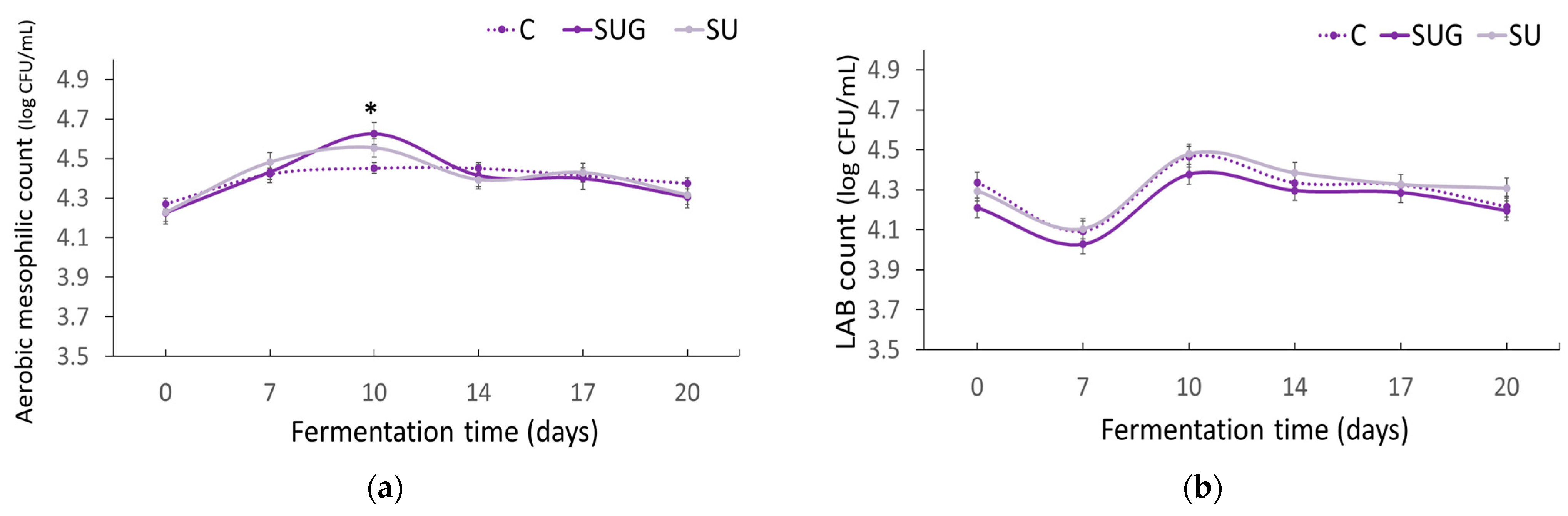
3.6. Evolution of the Microbiological Count
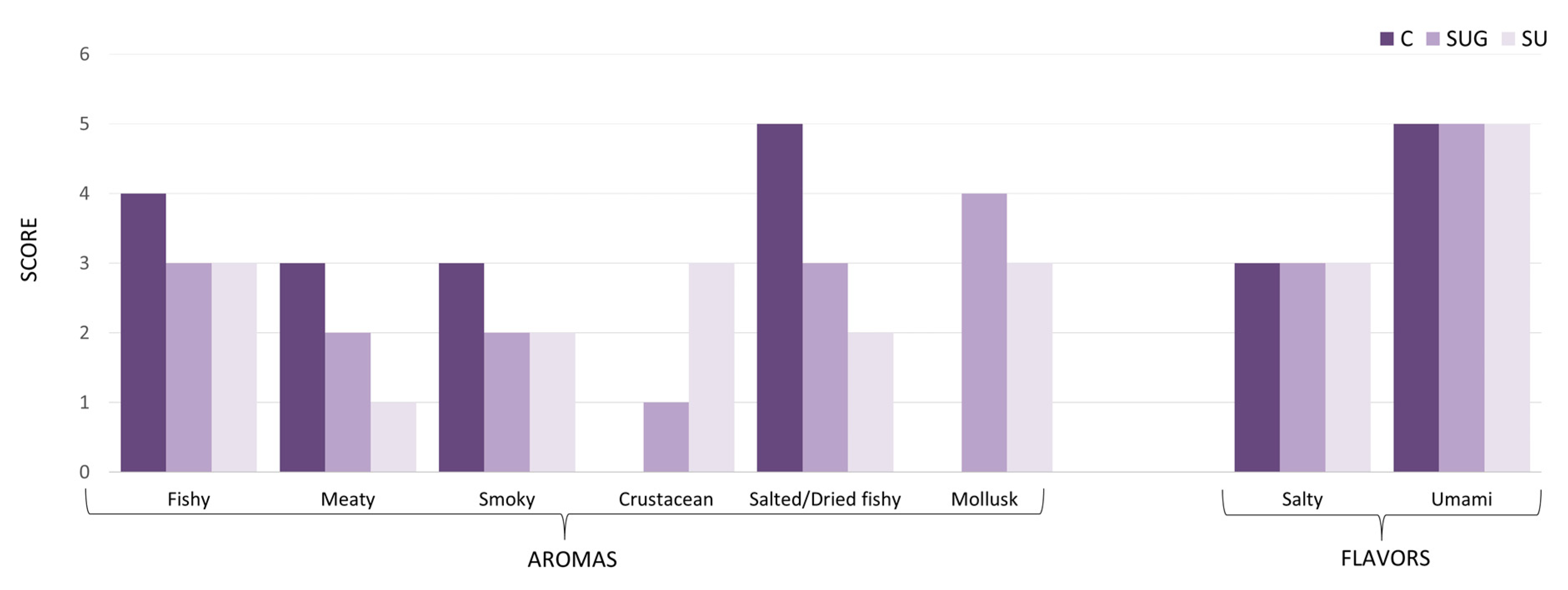
3.7. Quantitative Descriptive Analysis (QDA) of Sauces
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, J.; Zeng, Q.; Zhu, Z.; Zhang, L. Chemical and Sensory Changes Associated Yu-Lu Fermentation Process—A Traditional Chinese Fish Sauce. Food Chem. 2007, 104, 1629–1634. [Google Scholar] [CrossRef]
- Lai, Q.D.; Nguyen, H.D. Enhancement of Fish Sauce Quality by Application of Nanofiltration. LWT Food Sci. Technol. 2021, 151, 112181. [Google Scholar] [CrossRef]
- Chan, S.X.Y.; Fitri, N.; Mio Asni, N.S.; Sayuti, N.H.; Azlan, U.K.; Qadi, W.S.M.; Dawoud, E.A.D.; Kamal, N.; Sarian, M.N.; Mohd Lazaldin, M.A.; et al. A Comprehensive Review with Future Insights on the Processing and Safety of Fermented Fish and the Associated Changes. Foods 2023, 12, 558. [Google Scholar] [CrossRef]
- Lapsongphon, N.; Yongsawatdigul, J.; Cadwallader, K.R. Identification and Characterization of the Aroma-Impact Components of Thai Fish Sauce. J. Agric. Food Chem. 2015, 63, 2628–2638. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Duelund, L.; Calleja, G.; Frøst, M.B. Flavour of Fermented Fish, Insect, Game, and Pea Sauces: Garum Revisited. Int. J. Gastron. Food Sci. 2017, 9, 16–28. [Google Scholar] [CrossRef]
- Kilinc, B.; Cakli, S.; Tolasa, S.; Dincer, T. Chemical, Microbiological and Sensory Changes Associated with Fish Sauce Processing. Eur. Food Res. Technol. 2006, 222, 604–613. [Google Scholar] [CrossRef]
- Lopetcharat, K.; Choi, Y.J.; Park, J.W.; Daeschel, M.A. Fish Sauce Products and Manufacturing: A Review. Food Rev. Int. 2001, 17, 65–88. [Google Scholar] [CrossRef]
- Joint FAO/WHO Food Standards Program Codex Alimentarius Commission. Thirty-First Session Geneva, Switzerland, 30 June–4 July 2008. Available online: https://www.fao.org/3/i0420e/i0420e00.htm (accessed on 1 October 2023).
- Faisal, M.; Islami, S.N.E.; Islam, M.N.; Kamal, M.; Khan, M.N.A. Study on Microbial and Physical Changes in Fish Sauce during Fermentation. Res. Agric. Livest. Fish. 2015, 2, 375–383. [Google Scholar] [CrossRef]
- Klomklao, S.; Benjakul, S.; Visessanguan, W.; Kishimura, H.; Simpson, B.K. Effects of the Addition of Spleen of Skipjack Tuna (Katsuwonus pelamis) on the Liquefaction and Characteristics of Fish Sauce Made from Sardine (Sardinella gibbosa). Food Chem. 2006, 98, 440–452. [Google Scholar] [CrossRef]
- Gildberg, A. Utilisation of Male Arctic Capelin and Atlantic Cod Intestines for Fish Sauce Production—Evaluation of Fermentation Conditions. Bioresour. Technol. 2001, 76, 119–123. [Google Scholar] [CrossRef]
- Saisithi, P. Traditional Fermented Fish: Fish Sauce Production. In Fisheries Processing; Springer: Boston, MA, USA, 1994; pp. 111–131. [Google Scholar]
- Bu, Y.; Liu, Y.; Luan, H.; Zhu, W.; Li, X.; Li, J. Changes in Protease Activity during Fermentation of Fish Sauce and Their Correlation with Antioxidant Activity. J. Sci. Food Agric. 2022, 102, 3150–3159. [Google Scholar] [CrossRef] [PubMed]
- Koo, O.K.; Lee, S.J.; Chung, K.R.; Jang, D.J.; Yang, H.J.; Kwon, D.Y. Korean Traditional Fermented Fish Products: Jeotgal. J. Ethn. Foods 2016, 3, 107–116. [Google Scholar] [CrossRef]
- Kuda, T. Quality Improvement and Fermentation Control in Fish Products. In Advances in Fermented Foods and Beverages; Elsevier: Amsterdam, The Netherlands, 2015; pp. 377–390. [Google Scholar]
- Ohmori, T.; Mutaguchi, Y.; Yoshikawa, S.; Doi, K.; Ohshima, T. Amino Acid Components of Lees in Salmon Fish Sauce Are Tyrosine and Phenylalanine. J. Biosci. Bioeng. 2011, 112, 256–258. [Google Scholar] [CrossRef] [PubMed]
- Furesi, R.; Madau, F.A.; Pulina, P.; Sai, R.; Pinna, M.G.; Pais, A. Profitability and Sustainability of Edible Sea Urchin Fishery in Sardinia (Italy). J. Coast. Conserv. 2016, 20, 299–306. [Google Scholar] [CrossRef]
- Rocha, F.; Rocha, A.C.; Baião, L.F.; Gadelha, J.; Camacho, C.; Carvalho, M.L.; Arenas, F.; Oliveira, A.; Maia, M.R.G.; Cabrita, A.R.; et al. Seasonal Effect in Nutritional Quality and Safety of the Wild Sea Urchin Paracentrotus lividus Harvested in the European Atlantic Shores. Food Chem. 2019, 282, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Marzorati, S.; Martinelli, G.; Sugni, M.; Verotta, L. Green Extraction Strategies for Sea Urchin Waste Valorization. Front. Nutr. 2021, 8, 730747. [Google Scholar] [CrossRef] [PubMed]
- Zilia, F.; Bacenetti, J.; Sugni, M.; Matarazzo, A.; Orsi, L. From Waste to Product: Circular Economy Applications from Sea Urchin. Sustainability 2021, 13, 5427. [Google Scholar] [CrossRef]
- Sibiya, A.; Jeyavani, J.; Sivakamavalli, J.; Ravi, C.; Divya, M.; Vaseeharan, B. Bioactive Compounds from Various Types of Sea Urchin and Their Therapeutic Effects—A Review. Reg. Stud. Mar. Sci. 2021, 44, 101760. [Google Scholar] [CrossRef]
- Baião, L.F.; Moura, A.P.; Rocha, C.; Valente, L.M.P.; Cunha, L.M. Dimensions for the Valorisation of Sea Urchin (Paracentrotus Lividus) Gonads Production through the Eyes of Experienced Chefs. Int. J. Gastron. Food Sci. 2021, 26, 100438. [Google Scholar] [CrossRef]
- Matveeva, V.A.; Shulgina, L.V.; Prikhodko, Y.V.; Shulgin, Y.P.; Madej, K.; Piekoszewski, W. Nutritional Value of Sea Urchin Roe (Strongylocentrotidae)—Study of Composition and Storage Conditions. Separations 2021, 8, 174. [Google Scholar] [CrossRef]
- Kostetsky, E.Y.; Velansky, P.V.; Sanina, N.M. Phospholipids of the Organs and Tissues of Echinoderms and Tunicates from Peter the Great Bay (Sea of Japan). Russ. J. Mar. Biol. 2012, 38, 64–71. [Google Scholar] [CrossRef]
- Cobb, B.F.; Alaniz, I.; Thompson, C.A. Biochemical and Microbial Studies on Shrimp: Volatile Nitrogen and Amino Nitrogen Analysis. J. Food Sci. 1973, 38, 431–436. [Google Scholar] [CrossRef]
- Sánchez-García, F.; Hernández, I.; Palacios, V.M.; Roldán, A.M. Freshness Quality and Shelf Life Evaluation of the Seaweed Ulva Rigida through Physical, Chemical, Microbiological, and Sensory Methods. Foods 2021, 10, 181. [Google Scholar] [CrossRef] [PubMed]
- Kunitz, M. Crystaline Soybean Trypsin Inhibitor: II. General Properties. J. Gen. Physiol. 1947, 30, 291–310. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 2005; Volume 18, ISBN 0-935584-74. [Google Scholar]
- Dissaraphong, S.; Benjakul, S.; Visessanguan, W.; Kishimura, H. The Influence of Storage Conditions of Tuna Viscera before Fermentation on the Chemical, Physical and Microbiological Changes in Fish Sauce during Fermentation. Bioresour. Technol. 2006, 97, 2032–2040. [Google Scholar] [CrossRef]
- Angor, M. Physicochemical and Sensory Properties of Jameed Enriched with Orange Albedo and Apple Peel Powder. Afr. J. Food Agric. Nutr. Dev. 2023, 23, 22440–22451. [Google Scholar] [CrossRef]
- Ghayoomi, H.; Habibi Najafi, M.B.; Edalatian Dovom, M.R.; Pourfarzad, A. Low Salt and Biogenic Amines Fermented Fish Sauce (Mahyaveh) as Potential Functional Food and Ingredient. LWT 2023, 182, 114801. [Google Scholar] [CrossRef]
- Rajauria, G.; Sharma, S.; Emerald, M.; Jaiswal, A.K. Novel Fermented Marine-Based Products. In Novel Food Fermentation Technologies; Springer: Berlin/Heidelberg, Germany, 2016; pp. 235–262. [Google Scholar]
- Zeng, X.; Xia, W.; Jiang, Q.; Yang, F. Effect of Autochthonous Starter Cultures on Microbiological and Physico-Chemical Characteristics of Suan Yu, a Traditional Chinese Low Salt Fermented Fish. Food Control 2013, 33, 344–351. [Google Scholar] [CrossRef]
- Zang, J.; Xu, Y.; Xia, W.; Regenstein, J.M. Quality, Functionality, and Microbiology of Fermented Fish: A Review. Crit. Rev. Food Sci. Nutr. 2020, 60, 1228–1242. [Google Scholar] [CrossRef]
- Taira, W.; Funatsu, Y.; Satomi, M.; Takano, T.; Abe, H. Changes in Extractive Components and Microbial Proliferation during Fermentation of Fish Sauce from Underutilized Fish Species and Quality of Final Products. Fish. Sci. 2007, 73, 913–923. [Google Scholar] [CrossRef]
- Nakano, M.; Sagane, Y.; Koizumi, R.; Nakazawa, Y.; Yamazaki, M.; Watanabe, T.; Takano, K.; Sato, H. Data on the Chemical Properties of Commercial Fish Sauce Products. Data Brief 2017, 15, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Park, J.N.; Fukumoto, Y.; Fujita, E.; Tanaka, T.; Washio, T.; Otsuka, S.; Shimizu, T.; Watanabe, K.; Abe, H. Chemical Composition of Fish Sauces Produced in Southeast and East Asian Countries. J. Food Compos. Anal. 2001, 14, 113–125. [Google Scholar] [CrossRef]
- Horner, W.F.A. Preservation of Fish by Curing (Drying, Salting and Smoking). In Fish Processing Technology; Springer: Boston, MA, USA, 1997; pp. 32–73. [Google Scholar]
- Kuda, T.; Mihara, T.; Yano, T. Detection of Histamine and Histamine-Related Bacteria in Fish-Nukazuke, a Salted and Fermented Fish with Rice-Bran, by Simple Colorimetric Microplate Assay. Food Control 2007, 18, 677–681. [Google Scholar] [CrossRef]
- Mah, J.-H.; Hwang, H.-J. Effects of Food Additives on Biogenic Amine Formation in Myeolchi-Jeot, a Salted and Fermented Anchovy (Engraulis Japonicus). Food Chem. 2009, 114, 168–173. [Google Scholar] [CrossRef]
- Mathlouthi, M. Water Content, Water Activity, Water Structure and the Stability of Foodstuffs. Food Control 2001, 12, 409–417. [Google Scholar] [CrossRef]
- Varkoulis, A.; Voulgaris, K.; Zaoutsos, S.; Stratakis, A.; Vafidis, D. Chemical Composition and Microstructural Morphology of Spines and Tests of Three Common Sea Urchins Species of the Sublittoral Zone of the Mediterranean Sea. Animals 2020, 10, 1351. [Google Scholar] [CrossRef]
- Harikedua, S.D.; Wijaya, C.H.; Adawiyah, D.R. Relationship between Sensory Attributes of Bakasang (a Traditional Indonesian Fermented Fish Product) and Its Physicochemical Properties. Fish. Sci. 2012, 78, 187–195. [Google Scholar] [CrossRef]
- Hosseini, S.F.; Rezaei, M.; Zandi, M.; Ghavi, F.F. Effect of Fish Gelatin Coating Enriched with Oregano Essential Oil on the Quality of Refrigerated Rainbow Trout Fillet. J. Aquat. Food Prod. Technol. 2016, 25, 835–842. [Google Scholar] [CrossRef]
- Kaur, B.P.; Kaushik, N.; Rao, P.S.; Chauhan, O.P. Effect of High-Pressure Processing on Physical, Biochemical, and Microbiological Characteristics of Black Tiger Shrimp (Penaeus Monodon). Food Bioprocess Technol. 2013, 6, 1390–1400. [Google Scholar] [CrossRef]
- Park, S.; Sung, J.; Choi, H.-Y.; Jeong, J.; Jeong, H.-G.; Kim, J.-C.; Jang, M. Changes in the Physicochemical Properties and Metabolites of Myeolchi-Jeot (Salted-Fermented Anchovy) Based on Fermentation Time. LWT 2023, 177, 114456. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial Surface Colonization and Biofilm Development in Marine Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.M.; Lawrence, A.L.; Watts, S.A. Feeding, Digestion and Digestibility of Sea Urchins. In Sea Urchins: Biology and Ecology; Elsevier Science: Amsterdam, The Netherlands, 2013; pp. 135–154. [Google Scholar]
- Trenzado, C.E.; Hidalgo, F.; Villanueva, D.; Furné, M.; Díaz-Casado, M.E.; Merino, R.; Sanz, A. Study of the Enzymatic Digestive Profile in Three Species of Mediterranean Sea Urchins. Aquaculture 2012, 344–349, 174–180. [Google Scholar] [CrossRef]
- Pérez-Jiménez, A.; Cardenete, G.; Morales, A.E.; García-Alcázar, A.; Abellán, E.; Hidalgo, M.C. Digestive Enzymatic Profile of Dentex dentex and Response to Different Dietary Formulations. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 154, 157–164. [Google Scholar] [CrossRef]
- Jamaludin, S.L.; Rehena, J.F.; Seumahu, C.A.; Rumahlatu, D. Isolation and Identification of Protease Enzyme Producing Bacteria from Fermentation of Gonad Sea Urchin (Echinothrix calamaris). Ilmu Kelaut. Indones. J. Mar. Sci. 2019, 23, 187–198. [Google Scholar] [CrossRef]
- Tungkawachara, S.; Park, J.W.; Choi, Y.J. Biochemical Properties and Consumer Acceptance of Pacific Whiting Fish Sauce. J. Food Sci. 2003, 68, 855–860. [Google Scholar] [CrossRef]
- Xu, W.; Yu, G.; Xue, C.; Xue, Y.; Ren, Y. Biochemical Changes Associated with Fast Fermentation of Squid Processing By-Products for Low Salt Fish Sauce. Food Chem. 2008, 107, 1597–1604. [Google Scholar] [CrossRef]
- Orejana, F.M.; Liston, J. Agents of Proteolysis and Its Inhibition in Patis (Fish Sauce) Fermentation. J. Food Sci. 1982, 47, 198–203. [Google Scholar] [CrossRef]
- Beddows, C.G. Fermented Fish and Fish Products. In Microbiology of Fermented Foods; Springer: Boston, MA, USA, 1998; pp. 416–440. [Google Scholar]
- Hjalmarsson, G.H.; Park, J.W.; Kristbergsson, K. Seasonal Effects on the Physicochemical Characteristics of Fish Sauce Made from Capelin (Mallotus villosus). Food Chem. 2007, 103, 495–504. [Google Scholar] [CrossRef]
- Lopetcharat, K.; Park, J.W. Characteristics of Fish Sauce Made from Pacific Whiting and Surimi By-Products during Fermentation Stage. J. Food Sci. 2002, 67, 511–516. [Google Scholar] [CrossRef]
- Ijong, F.G.; Ohta, Y. Physicochemical and Microbiological Changes Associated with Bakasang Processing—A Traditional Indonesian Fermented Fish Sauce. J. Sci. Food Agric. 1996, 71, 69–74. [Google Scholar] [CrossRef]
- Chaveesuk, R.; Smith, J.P.; Simpson, B.K. Production of Fish Sauce and Acceleration of Sauce Fermentation Using Proteolytic Enzymes. J. Aquat. Food Prod. Technol. 1994, 2, 59–77. [Google Scholar] [CrossRef]
- Navarrete del Toro, M.A.; García-Carreño, F.L. Evaluation of the Progress of Protein Hydrolysis. Curr. Protoc. Food Anal. Chem. 2002, 10, B2.2.1–B2.2.14. [Google Scholar] [CrossRef]
- Department of Industry. Local Fish Sauce Standard Thai Industrial Standard; Department of Industry: Bangkok, Thailand, 1983. [Google Scholar]
- Auttanak, B.; Chomnawang, P.; Huaisan, K.; Sootsuwan, K.; Chomnawang, C. Influences of Salt and Koji Contents on Chemical Properties of Fish Sauce from Tilapia Frames. Aquac. Fish. 2022, 7, 67–73. [Google Scholar] [CrossRef]
- Singracha, P.; Niamsiri, N.; Visessanguan, W.; Lertsiri, S.; Assavanig, A. Application of Lactic Acid Bacteria and Yeasts as Starter Cultures for Reduced-Salt Soy Sauce (Moromi) Fermentation. LWT Food Sci. Technol. 2017, 78, 181–188. [Google Scholar] [CrossRef]
- Ding, A.; Zhu, M.; Qian, X.; Shi, L.; Huang, H.; Xiong, G.; Wang, J.; Wang, L. Effect of Fatty Acids on the Flavor Formation of Fish Sauce. Food Sci. Technol. 2020, 134, 110259. [Google Scholar] [CrossRef]
- Giri, A.; Osako, K.; Ohshima, T. Identification and Characterisation of Headspace Volatiles of Fish Miso, a Japanese Fish Meat Based Fermented Paste, with Special Emphasis on Effect of Fish Species and Meat Washing. Food Chem. 2010, 120, 621–631. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arango-Herrán, M.; Sánchez-García, F.; Palacios, V.M.; Roldán, A.M. The Impact of Sea Urchin as an Ingredient on the Physicochemical, Microbiological, and Sensory Properties of Fish Sauce Fermentation. Foods 2023, 12, 3958. https://doi.org/10.3390/foods12213958
Arango-Herrán M, Sánchez-García F, Palacios VM, Roldán AM. The Impact of Sea Urchin as an Ingredient on the Physicochemical, Microbiological, and Sensory Properties of Fish Sauce Fermentation. Foods. 2023; 12(21):3958. https://doi.org/10.3390/foods12213958
Chicago/Turabian StyleArango-Herrán, Mauricio, Fini Sánchez-García, Víctor M. Palacios, and Ana M. Roldán. 2023. "The Impact of Sea Urchin as an Ingredient on the Physicochemical, Microbiological, and Sensory Properties of Fish Sauce Fermentation" Foods 12, no. 21: 3958. https://doi.org/10.3390/foods12213958