
Recent and Advanced DNA-Based Technologies for the Authentication of Probiotic, Protected Designation of Origin (PDO) and Protected Geographical Indication (PGI) Fermented Foods and Beverages


Abstract
:
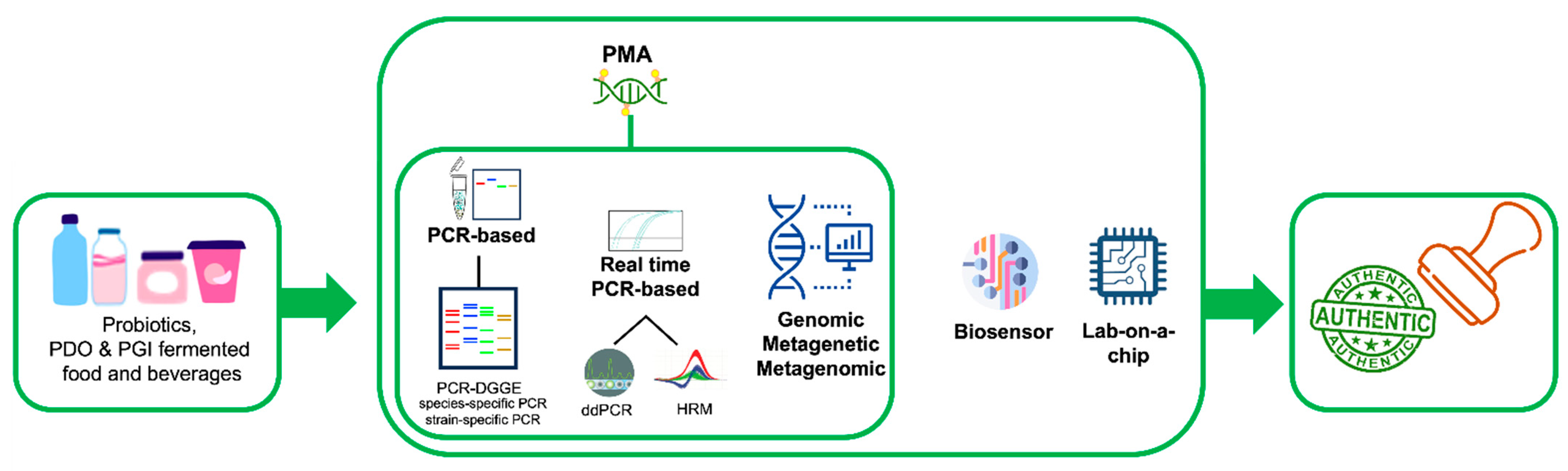
1. Introduction
2. Methods
2.1. Polymerase Chain Reaction (PCR)-Based Technologies
2.2. PCR-Based Typing Methods and Whole Genome Sequencing
2.3. Real-Time PCR-Based Methods
2.4. DNA-Based Biosensors and Lab-on-a-Chip Devices
2.5. PCR-DGGE
2.6. Metagenetics and Metagenomics
3. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Food and Drug Administration, FDA. Economically Motivated Adulteration (Food Fraud). 2023. Available online: https://www.fda.gov/food/compliance-enforcement-food/economically-motivated-adulteration-food-fraud (accessed on 1 June 2023).
- Marco, M.L.; Sanders, M.E.; Gänzle, M.; Arrieta, M.C.; Cotter, P.D.; De Vuyst, L.; Hill, C.; Holzapfel, W.; Lebeer, S.; Merenstein, D.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on fermented foods. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 196–208. [Google Scholar] [CrossRef]
- European Union (EU). Geographical Indications and Quality Schemes Explained. 2023. Available online: https://agriculture.ec.europa.eu/farming/geographical-indications-and-quality-schemes/geographical-indications-and-quality-schemes-explained_en (accessed on 1 June 2023).
- European Union. Regulation (EU) No. 1151/2012 of the European Parliament and of the Council, of 21 November 2012, on Quality Schemes for Agricultural Products and Foodstuffs. 2023. Available online: https://eur-lex.europa.eu/legal-content/it/ALL/?uri=CELEX:32012R1151 (accessed on 1 June 2023).
- Dias, C.; Mendes, L. Protected Designation of Origin (PDO), Protected Geographical Indication (PGI) and Traditional Speciality Guaranteed (TSG): A bibiliometric analysis. Food Res. Int. 2018, 103, 492–508. [Google Scholar] [CrossRef] [PubMed]
- Sckokai, P.; Soregaroli, C.; Moro, D. Estimating market power by retailers in a dynamic framework: The Italian PDO cheese market. J. Agric. Econ. 2012, 64, 33–53. [Google Scholar] [CrossRef]
- Enjoy European Quality. Salumi. 2023. Available online: https://www.enjoyeuropeanquality.it/salumi/ (accessed on 1 June 2023).
- Gori, C.; Sottini, V.A. The role of the Consortia in the Italian wine production system and the impact of EU and national legislation. Wine Econ. Policy 2014, 3, 62–67. [Google Scholar] [CrossRef]
- Danezis, G.P.; Tsagkaris, A.S.; Brusic, V.; Georgiou, C.A. Food authentication: State of the art and prospects. Curr. Opin. Food Sci. 2016, 10, 22–31. [Google Scholar] [CrossRef]
- Deng, L.; Liu, L.; Fu, T.; Li, C.; Jin, N.; Zhang, H.; Li, C.; Liu, Y.; Zhao, C. Genome sequence and evaluation of safety and probiotic potential of Lactiplantibacillus plantarum LPJZ-658. Microorganisms 2023, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- El Sheikha, A.F.; Hu, D.-M. Molecular techniques reveal more secrets of fermented foods. Crit. Rev. Food Sci. Nutr. 2018, 60, 11–32. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Fanelli, F.; de Souza, E.L. Editorial: Authenticity of Probiotic Foods and Dietary Supplements. Front. Microbiol. 2021, 12, 789049. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Fanelli, F.; Chieffi, D. Authenticity of probiotic foods and dietary supplements: A pivotal issue to address. Crit. Rev. Food Sci. Nutr. 2021, 62, 6854–6871, Erratum in Crit. Rev. Food Sci. Nutr. 2023, 63, 4210–4215. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Fanelli, F.; Chieffi, D. Authenticity of probiotic foods and supplements: Up-to-date situation and methods to assess it. In Probiotics for Human Nutrition in Health and Disease; Leite de Souza, E., de Brito Alves, J.L., Fusco, V., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 45–74. ISBN 9780323899086. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Saiki, R.K.; Scharf, S.J.; Faloona, F.; Mullis, K.B.; Horn, G.T.; Erlich, H.A.; Arnheim, N. Enzymatic amplification of β-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anemia. Science 1985, 230, 1350–1354. [Google Scholar] [CrossRef] [PubMed]
- Mullis, K.B.; Faloona, F.A. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol. 1987, 155, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Garibyan, L.; Avashia, N. Polymerase Chain Reaction. J. Investig. Dermatol. 2013, 133, 1–4. [Google Scholar] [CrossRef]
- Ansari, J.M.; Colasacco, C.; Emmanouil, E.; Kohlhepp, S.; Harriott, O. Strain-level diversity of commercial probiotic isolates of Bacillus, Lactobacillus, and Saccharomyces species illustrated by molecular identification and phenotypic profiling. PLoS ONE 2019, 14, e0213841. [Google Scholar] [CrossRef]
- Neviani, E.; Bottari, B.; Lazzi, C.; Gatti, M. New developments in the study of the microbiota of raw-milk, long-ripened cheeses by molecular methods: The case of Grana Padano and Parmigiano Reggiano. Front. Microbiol. 2013, 4, 36. [Google Scholar] [CrossRef]
- Lick, S.; Keller, M.; Bockelmann, W.; Heller, J. Rapid identification of Streptococcus thermophilus by primer-specific PCR amplification based on its lacZ gene. Syst. Appl. Microbiol. 1996, 19, 74–77. [Google Scholar] [CrossRef]
- Nikolaou, A.; Saxami, G.; Kourkoutas, Y.; Galanis, A. A new methodology for rapid detection of Lactobacillus delbrueckii subsp. bulgaricus based on multiplex PCR. J. Microbiol. Methods 2011, 84, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Quero, G.M.; Chieffi, D.; Franz, C.M. Identification of Lactobacillus brevis using a species-specific AFLP-derived marker. Int. J. Food Microbiol. 2016, 232, 90–94. [Google Scholar] [CrossRef]
- Fusco, V.; Quero, G.M.; Stea, G.; Morea, M.; Visconti, A. Novel PCR-based identification of Weissella confusa using an AFLP-derived marker. Int. J. Food Microbiol. 2011, 145, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Ward, L.J.H.; Timmins, M.J. Differentiation of Lactobacillus casei, Lactobacillus paracasei and Lactobacillus rhamnosus by polymerase chain reaction. Lett. Appl. Microbiol. 1999, 29, 90–92. [Google Scholar] [CrossRef]
- Fortina, M.G.; Ricci, G.; Mora, D.; Parini, C.; Manachini, P.L. Specific identification of Lactobacillus helveticus by PCR with pepC, pepN and htrA targeted primers. FEMS Microbiol. Lett. 2001, 198, 85–89. [Google Scholar] [CrossRef]
- Dickson, E.M.; Riggio, M.P.; Macpherson, L. A novel species-specific PCR assay for identifying Lactobacillus fermentum. J. Med Microbiol. 2005, 54, 299–303. [Google Scholar] [CrossRef]
- Torriani, S.; Felis, G.E.; Dellaglio, F. Differentiation of Lactobacillus plantarum, L. pentosus, and L. paraplantarum by recA gene sequence analysis and multiplex PCR assay with recA gene-derived primers. Appl. Environ. Microbiol. 2001, 67, 3450–3454. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Yang, S.-M.; Lim, B.; Park, S.H.; Rackerby, B.; Kim, H.-Y. Design of PCR assays to specifically detect and identify 37 Lactobacillus species in a single 96 well plate. BMC Microbiol. 2020, 20, 96. [Google Scholar] [CrossRef]
- Guglielmotti, D.M.; Pujato, S.A.; Quiberoni, A.; Suárez, V.B. Hsp60 gene as a reliable target for taxonomical identification and discrimination of Leuconostoc species of dairy origin. Int. Dairy J. 2021, 126, 105227. [Google Scholar] [CrossRef]
- Xiang, X.; Lu, J.; Xu, X.; Hou, X.; Diao, E.; Qian, S.; Song, H.; Liang, L.; He, Y.; Shang, Y. Rapid identification of novel specific molecular targets for PCR detection of four Enterococcus species. LWT 2023, 173, 114356. [Google Scholar] [CrossRef]
- Council for Responsible Nutrition and International Probiotics Association. Best Practices Guidelines for Probiotics. 2017. Available online: https://internationalprobiotics.org/resources/guidelines/2017-best-practices-guidelines/ (accessed on 1 June 2023).
- FAO/WHO (Food and Agriculture Organization of the United Nations/World Health Organization). Probiotics in Food. Health and Nutritional Properties and Guidelines for Evaluation. 2006. Available online: http://www.fao.org/3/a-a0512e.pdf (accessed on 1 June 2023).
- Fusco, V.; Quero, G.M. Culture-dependent and culture-independent nucleic-acid-based methods used in the microbial safety assessment of milk and dairy products. Compr. Rev. Food Sci. Food Saf. 2014, 13, 493–537. [Google Scholar] [CrossRef]
- Blandino, G.; Fazio, D.; Petronio, G.P.; Inturri, R.; Tempera, G.; Furneri, P.M. Labeling quality and molecular characterization studies of products containing Lactobacillus spp. strains. Int. J. Immunopathol. Pharmacol. 2015, 29, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Di Lena, M.; Quero, G.M.; Santovito, E.; Verran, J.; De Angelis, M.; Fusco, V. A selective medium for isolation and accurate enumeration of Lactobacillus casei-group lactobacilli in probiotic milks and dairy products. Int. Dairy J. 2015, 47, 27–36. [Google Scholar] [CrossRef]
- Masco, L.; Huys, G.; De Brandt, E.; Temmerman, R.; Swings, J. Culture-dependent and culture-independent qualitative analysis of probiotic products claimed to contain bifidobacteria. Int. J. Food Microbiol. 2005, 102, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Quero, G.M.; Poltronieri, P.; Morea, M.; Baruzzi, F. Autochthonous and probiotic lactic acid bacteria employed for production of “advanced traditional cheeses”. Foods 2019, 8, 412. [Google Scholar] [CrossRef]
- Quero, G.M.; Fusco, V.; Cocconcelli, P.S.; Owczarek, L.; Borcakli, M.; Fontana, C.; Skapska, S.; Jasinska, U.T.; Ozturk, T.; Morea, M. Microbiological, physico-chemical, nutritional and sensory characterization of traditional Matsoni: Selection and use of autochthonous multiple strain cultures to extend its shelf-life. Food Microbiol. 2014, 38, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Coppola, S.; Fusco, V.; Andolfi, R.; Aponte, M.; Blaiotta, G.; Ercolini, D.; Moschetti, G. Evaluating microbial diversity during the manufacture of “fior di latte di Agerola”, a traditional raw milk cheese of Naples area. J. Dairy Res. 2006, 73, 264–272. [Google Scholar] [CrossRef]
- Johnson, B.R.; Klaenhammer, T.R. Impact of genomics on the field of probiotic research: Historical perspectives to modern paradigms. Antonie van Leeuwenhoek 2014, 106, 141–156. [Google Scholar] [CrossRef]
- Peng, X.; Ed-Dra, A.; Yue, M. Whole genome sequencing for the risk assessment of probiotic lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2022, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Manetsberger, J.; Caballero Gómez, N.; Benomar, N. In silico genomic analysis of the potential probiotic Lactiplantibacillus pentosus CF2-10N reveals promising beneficial effects with health promoting properties. Front. Microbiol. 2022, 13, 989824, Erratum in Front. Microbiol. 2023, 14, 1242095. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Pérez Montoro, B.; Casado Muñoz, M.D.C.; Knapp, C.W.; Gálvez, A.; Benomar, N. In silico genomic insights into aspects of food safety and defense mechanisms of a potentially probiotic Lactobacillus pentosus MP-10 isolated from brines of naturally fermented Aloreña green table olives. PLoS ONE 2017, 12, e0176801. [Google Scholar] [CrossRef] [PubMed]
- Alayande, K.A.; Aiyegoro, O.A.; Nengwekhulu, T.M.; Katata-Seru, L.; Ateba, C.N. Integrated genome-based probiotic relevance and safety evaluation of Lactobacillus reuteri PNW1. PLoS ONE 2020, 15, e0235873. [Google Scholar] [CrossRef]
- Arellano, K.; Vazquez, J.; Park, H.; Lim, J.; Ji, Y.; Kang, H.-J.; Cho, D.; Jeong, H.W.; Holzapfel, W.H. Safety evaluation and whole-genome annotation of Lactobacillus plantarum strains from different sources with special focus on isolates from green tea. Probiotics Antimicrob. Proteins 2020, 12, 1057–1070. [Google Scholar] [CrossRef]
- Aziz, T.; Naveed, M.; Jabeen, K.; Shabbir, M.A.; Sarwar, A.; Zhennai, Y.; Alharbi, M.; Alshammari, A.; Alasmari, A.F. Integrated genome based evaluation of safety and probiotic characteristics of Lactiplantibacillus plantarum YW11 isolated from Tibetan kefir. Front. Microbiol. 2023, 14, 1157615. [Google Scholar] [CrossRef]
- Baccouri, O.; Boukerb, A.M.; Farhat, L.B.; Zébré, A.; Zimmermann, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I.; et al. Probiotic potential and safety evaluation of Enterococcus faecalis OB14 and OB15, isolated from Traditional tunisian testouri cheese and rigouta, using physiological and genomic analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef]
- Bae, W.-Y.; Lee, Y.J.; Jung, W.-H.; Shin, S.L.; Kim, T.-R.; Sohn, M. Draft genome sequence and probiotic functional property analysis of Lactobacillus gasseri LM1065 for food industry applications. Sci. Rep. 2023, 13, 12212. [Google Scholar] [CrossRef] [PubMed]
- Bang, W.Y.; Ban, O.-H.; Lee, B.S.; Oh, S.; Park, C.; Park, M.-K.; Jung, S.K.; Yang, J.; Jung, Y.H. Genomic-, phenotypic-, and toxicity-based safety assessment and probiotic potency of Bacillus coagulans IDCC 1201 isolated from green malt. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab026. [Google Scholar] [CrossRef] [PubMed]
- Boucard, A.-S.; Florent, I.; Polack, B.; Langella, P.; Bermúdez-Humarán, L.G. Genome sequence and assessment of safety and potential probiotic traits of Lactobacillus johnsonii CNCM I-4884. Microorganisms 2022, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, F.; Montemurro, M.; Verni, M.; Garbetta, A.; Bavaro, A.R.; Chieffi, D.; Cho, G.-S.; Franz, C.M.A.P.; Rizzello, C.G.; Fusco, V. Probiotic potential and safety assessment of type strains of Weissella and Periweissella species. Microbiol. Spectr. 2023, 11, e0304722. [Google Scholar] [CrossRef] [PubMed]
- Fontana, A.; Falasconi, I.; Molinari, P.; Treu, L.; Basile, A.; Vezzi, A.; Campanaro, S.; Morelli, L. Genomic comparison of Lactobacillus helveticus strains highlights probiotic potential. Front. Microbiol. 2019, 10, 1380. [Google Scholar] [CrossRef]
- Heo, S.; Kim, J.-H.; Kwak, M.-S.; Jeong, D.-W.; Sung, M.-H. Functional genomic insights into probiotic Bacillus siamensis strain B28 from traditional Korean fermented kimchi. Foods 2021, 10, 1906. [Google Scholar] [CrossRef] [PubMed]
- Hussein, W.E.; Abdelhamid, A.G.; Rocha-Mendoza, D.; García-Cano, I.; Yousef, A.E. Assessment of safety and probiotic traits of Enterococcus durans OSY-EGY, isolated from Egyptian artisanal cheese, using comparative genomics and Phenotypic analyses. Front. Microbiol. 2020, 11, 608314. [Google Scholar] [CrossRef]
- Jiang, Y.-H.; Yang, R.-S.; Lin, Y.-C.; Xin, W.-G.; Zhou, H.-Y.; Wang, F.; Zhang, Q.-L.; Lin, L.-B. Assessment of the safety and probiotic characteristics of Lactobacillus salivarius CGMCC20700 based on whole-genome sequencing and phenotypic analysis. Front. Microbiol. 2023, 14, 1120263. [Google Scholar] [CrossRef]
- Lee, Y.-R.; Bang, W.Y.; Baek, K.-R.; Kim, G.-H.; Kang, M.-J.; Yang, J.; Seo, S.-O. Safety evaluation by phenotypic and genomic characterization of four Lactobacilli strains with probiotic properties. Microorganisms 2022, 10, 2218. [Google Scholar] [CrossRef]
- Li, B.; Zhan, M.; Evivie, S.E.; Jin, D.; Zhao, L.; Chowdhury, S.; Sarker, S.K.; Huo, G.; Liu, F. Evaluating the safety of potential probiotic Enterococcus durans KLDS6.0930 using whole genome sequencing and oral toxicity study. Front. Microbiol. 2018, 9, 1943. [Google Scholar] [CrossRef]
- Mileriene, J.; Aksomaitiene, J.; Kondrotiene, K.; Asledottir, T.; Vegarud, G.E.; Serniene, L.; Malakauskas, M. Whole-genome sequence of Lactococcus lactis subsp. lactis LL16 confirms safety, probiotic potential, and reveals functional traits. Microorganisms 2023, 11, 1034. [Google Scholar] [CrossRef]
- Oh, Y.J.; Kim, S.-A.; Yang, S.H.; Kim, D.H.; Cheng, Y.-Y.; Kang, J.I.; Lee, S.Y.; Han, N.S. Integrated genome-based assessment of safety and probiotic characteristics of Lactiplantibacillus plantarum PMO 08 isolated from kimchi. PLoS ONE 2022, 17, e0273986. [Google Scholar] [CrossRef] [PubMed]
- Raethong, N.; Santivarangkna, C.; Visessanguan, W.; Santiyanont, P.; Mhuantong, W.; Chokesajjawatee, N. Whole-genome sequence analysis for evaluating the safety and probiotic potential of Lactiplantibacillus pentosus 9D3, a gamma-aminobutyric acid (GABA)-producing strain isolated from Thai pickled weed. Front. Microbiol. 2022, 13, 969548. [Google Scholar] [CrossRef]
- Salvetti, E.; Orrù, L.; Capozzi, V.; Martina, A.; Lamontanara, A.; Keller, D.; Cash, H.; Felis, G.E.; Cattivelli, L.; Torriani, S.; et al. Integrate genome-based assessment of safety for probiotic strains: Bacillus coagulans GBI-30, 6086 as a case study. Appl. Microbiol. Biotechnol. 2016, 100, 4595–4605. [Google Scholar] [CrossRef] [PubMed]
- Saroj, D.B.; Gupta, A.K. Genome based safety assessment for Bacillus coagulans strain LBSC (DSM 17654) for probiotic application. Int. J. Food Microbiol. 2020, 318, 108523. [Google Scholar] [CrossRef] [PubMed]
- Senan, S.; Prajapati, J.B.; Joshi, C.G. Feasibility of genome-wide screening for biosafety assessment of probiotics: A case study of Lactobacillus helveticus MTCC 5463. Probiotics Antimicrob. Proteins 2015, 7, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Stergiou, O.S.; Tegopoulos, K.; Kiousi, D.E.; Tsifintaris, M.; Papageorgiou, A.C.; Tassou, C.C.; Chorianopoulos, N.; Kolovos, P.; Galanis, A. Whole-genome sequencing, phylogenetic and genomic analysis of Lactiplantibacillus pentosus L33, a potential probiotic strain isolated from fermented sausages. Front. Microbiol. 2021, 12, 746659. [Google Scholar] [CrossRef] [PubMed]
- Surachat, K.; Kantachote, D.; Deachamag, P.; Wonglapsuwan, M. Genomic insight into Pediococcus acidilactici HN9, a potential probiotic strain isolated from the traditional Thai-style fermented beef Nhang. Microorganisms 2020, 9, 50. [Google Scholar] [CrossRef] [PubMed]
- Toropov, V.; Demyanova, E.; Shalaeva, O.; Sitkin, S.; Vakhitov, T. Whole-genome sequencing of Lactobacillus helveticus D75 and D76 confirms safety and probiotic potential. Microorganisms 2020, 8, 329. [Google Scholar] [CrossRef]
- Umanets, A.; Surono, I.S.; Venema, K. I am better than I look: Genome based safety assessment of the probiotic Lactiplantibacillus plantarum IS-10506. BMC Genom. 2023, 24, 518. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Y.; Liu, Y.; Zhong, J.; Zhang, D. Assessing the safety and probiotic characteristics of Lacticaseibacillus rhamnosus X253 via complete genome and phenotype analysis. Microorganisms 2023, 11, 140. [Google Scholar] [CrossRef]
- Castro-López, C.; García, H.S.; Martínez-Ávila, G.C.G.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Genomics-based approaches to identify and predict the health-promoting and safety activities of promising probiotic strains—A probiogenomics review. Trends Food Sci. Technol. 2020, 108, 148–163. [Google Scholar] [CrossRef]
- Fusco, V.; Riccardi, M.; Quero, G.M. Thin agar layer-versus most probable number-PCR to enumerate viable and stressed Escherichia coli O157:H7 and application in a traditional raw milk pasta filata cheese. Int. J. Food Microbiol. 2012, 159, 1–8. [Google Scholar] [CrossRef]
- Zand, E.; Froehling, A.; Schoenher, C.; Zunabovic-Pichler, M.; Schlueter, O.; Jaeger, H. Potential of flow cytometric approaches for rapid microbial detection and characterization in the food industry—A review. Foods 2021, 10, 3112. [Google Scholar] [CrossRef]
- Tong, S.Y.C.; Giffard, P.M. Microbiological applications of high-resolution melting analysis. J. Clin. Microbiol. 2012, 50, 3418–3421. [Google Scholar] [CrossRef]
- Wilhelm, J.; Pingoud, A. Real-time polymerase chain reaction. ChemBioChem 2003, 4, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
- Wittwer, C.T. High-resolution DNA melting analysis: Advancements and limitations. Hum. Mutat. 2009, 30, 857–859. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Chen, S.; Zheng, Y.; Zheng, X.; Lin, J.-M. Droplet-based digital PCR (ddPCR) and its applications. TrAC Trends Anal. Chem. 2023, 158, 116897. [Google Scholar] [CrossRef]
- Achilleos, C.; Berthier, F. Quantitative PCR for the specific quantification of Lactococcus lactis and Lactobacillus paracasei and its interest for Lactococcus lactis in cheese samples. Food Microbiol. 2013, 36, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Herbel, S.; Lauzat, B.; von Nickisch-Rosenegk, M.; Kuhn, M.; Murugaiyan, J.; Wieler, L.; Guenther, S. Species-specific quantification of probiotic lactobacilli in yoghurt by quantitative real-time PCR. J. Appl. Microbiol. 2013, 115, 1402–1410. [Google Scholar] [CrossRef]
- Iacumin, L.; Ginaldi, F.; Manzano, M.; Anastasi, V.; Reale, A.; Zotta, T.; Rossi, F.; Coppola, R.; Comi, G. High resolution melting analysis (HRM) as a new tool for the identification of species belonging to the Lactobacillus casei group and comparison with species-specific PCRs and multiplex PCR. Food Microbiol. 2015, 46, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kim, D.; Yang, S.-M.; Kim, H.-Y. Validation of probiotic species or subspecies identity in commercial probiotic products using high-resolution PCR method based on large-scale genomic analysis. Food Res. Int. 2022, 154, 111011. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Yang, S.-M.; Choi, C.H.; Shin, M.-K.; Kim, H.-Y. Droplet digital PCR method for the absolute quantitative detection and monitoring of Lacticaseibacillus casei. Food Microbiol. 2023, 113, 104265. [Google Scholar] [CrossRef]
- Kurbakov, K.A.; Konorov, E.A.; Minaev, M.Y.; Kuznetsova, O.A. Multiplex Real-Time PCR with HRM for Detection of Lactobacillus sakei and Lactobacillus curvatus in Food Samples. Food Technol. Biotechnol. 2019, 57, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Le Dréan, G.; Mounier, J.; Vasseur, V.; Arzur, D.; Habrylo, O.; Barbier, G. Quantification of Penicillium camemberti and P. roqueforti mycelium by real-time PCR to assess their growth dynamics during ripening cheese. Int. J. Food Microbiol. 2010, 138, 100–107. [Google Scholar] [CrossRef]
- Masco, L.; Vanhoutte, T.; Temmerman, R.; Swings, J.; Huys, G. Evaluation of real-time PCR targeting the 16S rRNA and recA genes for the enumeration of bifidobacteria in probiotic products. Int. J. Food Microbiol. 2007, 113, 351–357. [Google Scholar] [CrossRef]
- Pontonio, E.; Di Cagno, R.; Mahony, J.; Lanera, A.; De Angelis, M.; van Sinderen, D.; Gobbetti, M. Sourdough authentication: Quantitative PCR to detect the lactic acid bacterial microbiota in breads. Sci. Rep. 2017, 7, 624. [Google Scholar] [CrossRef]
- Shehata, H.R.; Newmaster, S.G. A Validated real-time PCR method for the specific identification of probiotic strain Lactobacillus rhamnosus GG (ATCC 53103). J. AOAC Int. 2020, 103, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Shehata, H.R.; Newmaster, S.G. Combined targeted and non-targeted PCR based methods reveal high levels of compliance in probiotic products sold as dietary supplements in United States and Canada. Front. Microbiol. 2020, 11, 1095. [Google Scholar] [CrossRef]
- Shehata, H.R.; Newmaster, S.G. Enumeration of probiotic strain Lacticaseibacillus rhamnosus GG (ATCC 53103) using viability real-time PCR. Probiotics Antimicrob. Proteins 2021, 13, 1611–1620. [Google Scholar] [CrossRef] [PubMed]
- Shehata, H.R.; Ragupathy, S.; Allen, S.; Leyer, G.; Newmaster, S.G. Real-time PCR assays for the specific identification of probiotic strains Lactobacillus gasseri BNR17 and Lactobacillus reuteri LRC (NCIMB 30242). Probiotics Antimicrob. Proteins 2020, 13, 837–846. [Google Scholar] [CrossRef]
- Sheu, S.-J.; Hwang, W.-Z.; Chiang, Y.-C.; Lin, W.-H.; Chen, H.-C.; Tsen, H.-Y. Use of tuf gene-based primers for the PCR detection of probiotic Bifidobacterium species and enumeration of bifidobacteria in fermented milk by cultural and quantitative real-time PCR methods. J. Food Sci. 2010, 75, M521–M527. [Google Scholar] [CrossRef] [PubMed]
- Udomsil, N.; Chen, S.; Rodtong, S.; Yongsawatdigul, J. Quantification of viable bacterial starter cultures of Virgibacillus sp. and Tetragenococcus halophilus in fish sauce fermentation by real-time quantitative PCR. Food Microbiol. 2016, 57, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Vaccalluzzo, A.; Pino, A.; Bosco, G.; Caggia, C.; Randazzo, C.L. Quantitative PCR Assay as a tool for the detection of lactobacilli in sicilian table olives produced at an industrial scale. Fermentation 2023, 9, 355. [Google Scholar] [CrossRef]
- Bilen, M.; Dufour, J.-C.; Lagier, J.-C.; Cadoret, F.; Daoud, Z.; Dubourg, G.; Raoult, D. The contribution of culturomics to the repertoire of isolated human bacterial and archaeal species. Microbiome 2018, 6, 94. [Google Scholar] [CrossRef] [PubMed]
- Barzideh, Z.; Siddiqi, M.; Mohamed, H.M.; LaPointe, G. Dynamics of starter and non-starter lactic acid bacteria populations in long-ripened cheddar cheese using propidium monoazide (PMA) treatment. Microorganisms 2022, 10, 1669. [Google Scholar] [CrossRef] [PubMed]
- Desfossés-Foucault, E.; Dussault-Lepage, V.; Le Boucher, C.; Savard, P.; LaPointe, G.; Roy, D. Assessment of probiotic viability during cheddar cheese manufacture and ripening using propidium monoazide-PCR quantification. Front. Microbiol. 2012, 3, 350. [Google Scholar] [CrossRef]
- Scariot, M.C.; Venturelli, G.L.; Prudêncio, E.S.; Arisi, A.C.M. Quantification of Lactobacillus paracasei viable cells in probiotic yoghurt by propidium monoazide combined with quantitative PCR. Int. J. Food Microbiol. 2018, 264, 1–7. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, Z.; Bao, Q.; Zhang, H. Application of propidium monoazide quantitative real-time PCR to quantify the viability of Lactobacillus delbrueckii ssp. bulgaricus. J. Dairy Sci. 2016, 99, 9570–9580. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, M.L.M.; Padilha, M.; Vieira, A.D.S.; Franco, B.D.; Martinez, R.C.R.; Saad, S.M.I. Advantageous direct quantification of viable closely related probiotics in petit-suisse cheeses under in vitro gastrointestinal conditions by propidium monoazide—qPCR. PLoS ONE 2013, 8, e82102. [Google Scholar] [CrossRef]
- Naresh, V.; Lee, N. A Review on biosensors and recent development of nanostructured materials-enabled biosensors. Sensors 2021, 21, 1109. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Ma, J.; Li, D.; Wang, R. DNA-based biosensors for the biochemical analysis: A review. Biosensors 2022, 12, 183. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.Y. A review on microscale polymerase chain reaction based methods in molecular diagnosis, and future prospects for the fabrication of fully integrated portable biomedical devices. Microchim. Acta 2018, 185, 285. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, F.; Li, Q.; Wang, L.; Fan, C. Isothermal amplification of nucleic acids. Chem. Rev. 2015, 115, 12491–12545. [Google Scholar] [CrossRef] [PubMed]
- Giuffrida, M.C.; Spoto, G. Integration of isothermal amplification methods in microfluidic devices: Recent advances. Biosens. Bioelectron. 2017, 90, 174–186. [Google Scholar] [CrossRef]
- Vidic, J.; Vizzini, P.; Manzano, M.; Kavanaugh, D.; Ramarao, N.; Zivkovic, M.; Radonic, V.; Knezevic, N.; Giouroudi, I.; Gadjanski, I. Point-of-need DNA testing for detection of foodborne pathogenic bacteria. Sensors 2019, 19, 1100. [Google Scholar] [CrossRef]
- Ercolini, D. PCR-DGGE fingerprinting: Novel strategies for detection of microbes in food. J. Microbiol. Methods 2004, 56, 297–314. [Google Scholar] [CrossRef]
- Chen, T.; Wu, Q.; Zhou, H.; Deng, K.; Wang, X.; Meng, F.; Yang, S.; Wang, X.; Shah, N.P.; Wei, H. Assessment of commercial probiotic products in China for labelling accuracy and probiotic characterisation of selected isolates. Int. J. Dairy Technol. 2016, 70, 119–126. [Google Scholar] [CrossRef]
- Elliot, E.; Teversham, K. An evaluation of nine probiotics available in South Africa, August 2003. S. Afr. Med. J. 2004, 94, 121–124. [Google Scholar]
- Fasoli, S.; Marzotto, M.; Rizzotti, L.; Rossi, F.; Dellaglio, F.; Torriani, S. Bacterial composition of commercial probiotic products as evaluated by PCR-DGGE analysis. Int. J. Food Microbiol. 2002, 82, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Patrone, V.; Molinari, P.; Morelli, L. Microbiological and molecular characterization of commercially available probiotics containing Bacillus clausii from India and Pakistan. Int. J. Food Microbiol. 2016, 237, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Theunissen, J.; Britz, T.; Torriani, S.; Witthuhn, R. Identification of probiotic microorganisms in South African products using PCR-based DGGE analysis. Int. J. Food Microbiol. 2005, 98, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Aponte, M.; Fusco, V.; Andolfi, R.; Coppola, S. Lactic acid bacteria occurring during manufacture and ripening of Provolone del Monaco cheese: Detection by different analytical approaches. Int. Dairy J. 2008, 18, 403–413. [Google Scholar] [CrossRef]
- Flórez, A.B.; Mayo, B. Microbial diversity and succession during the manufacture and ripening of traditional, Spanish, blue-veined Cabrales cheese, as determined by PCR-DGGE. Int. J. Food Microbiol. 2006, 110, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Arcuri, E.F.; El Sheikha, A.F.; Rychlik, T.; Piro-Métayer, I.; Montet, D. Determination of cheese origin by using 16S rDNA fingerprinting of bacteria communities by PCR–DGGE: Preliminary application to traditional Minas cheese. Food Control 2013, 30, 1–6. [Google Scholar] [CrossRef]
- Dufossé, L.; Donadio, C.; Valla, A.; Meile, J.-C.; Montet, D. Determination of speciality food salt origin by using 16S rDNA fingerprinting of bacterial communities by PCR–DGGE: An application on marine salts produced in solar salterns from the French Atlantic Ocean. Food Control 2013, 32, 644–649. [Google Scholar] [CrossRef]
- El Sheikha, A.F.; Bouvet, J.-M.; Montet, D. Biological bar code for determining the geographical origin of fruits using 28S rDNA fingerprinting of fungal communities by PCR-DGGE: An application to Shea tree fruits. Qual. Assur. Saf. Crop. Foods 2011, 3, 40–47. [Google Scholar] [CrossRef]
- El Sheikha, A.F.; Condur, A.; Métayer, I.; Le Nguyen, D.D.; Loiseau, G.; Montet, D. Determination of fruit origin by using 26S rDNA fingerprinting of yeast communities by PCR-DGGE: Preliminary application to Physalis fruits from Egypt. Yeast 2009, 26, 567–573. [Google Scholar] [CrossRef]
- El Sheikha, A.F.; Durand, N.; Sarter, S.; Okullo, J.B.; Montet, D. Study of the microbial discrimination of fruits by PCR-DGGE: Application to the determination of the geographical origin of Physalis fruits from Colombia, Egypt, Uganda and Madagascar. Food Control 2012, 24, 57–63. [Google Scholar] [CrossRef]
- Ercolini, D.; Frisso, G.; Mauriello, G.; Salvatore, F.; Coppola, S. Microbial diversity in Natural Whey Cultures used for the production of Caciocavallo Silano PDO cheese. Int. J. Food Microbiol. 2008, 124, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Ercolini, D.; Mauriello, G.; Blaiotta, G.; Moschetti, G.; Coppola, S. PCR-DGGE fingerprints of microbial succession during a manufacture of traditional water buffalo mozzarella cheese. J. Appl. Microbiol. 2004, 96, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Osimani, A.; Milanović, V.; Aquilanti, L.; De Filippis, F.; Stellato, G.; Di Mauro, S.; Turchetti, B.; Buzzini, P.; Ercolini, D.; et al. Bacteria and yeast microbiota in milk kefir grains from different Italian regions. Food Microbiol. 2015, 49, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Hamdouche, Y.; Guehi, T.; Durand, N.; Kedjebo, K.B.D.; Montet, D.; Meile, J.C. Dynamics of microbial ecology during cocoa fermentation and drying: Towards the identification of molecular markers. Food Control 2015, 48, 117–122. [Google Scholar] [CrossRef]
- Mauriello, G.; Moio, L.; Genovese, A.; Ercolini, D. Relationships between flavoring capabilities, bacterial composition, and geographical origin of natural whey cultures used for traditional water-buffalo mozzarella cheese manufacture. J. Dairy Sci. 2003, 86, 486–497. [Google Scholar] [CrossRef]
- Rychlik, T.; Szwengiel, A.; Bednarek, M.; Arcuri, E.; Montet, D.; Mayo, B.; Nowak, J.; Czarnecki, Z. Application of the PCR-DGGE technique to the fungal community of traditional Wielkopolska fried ripened curd cheese to determine its PGI authenticity. Food Control 2017, 73, 1074–1081. [Google Scholar] [CrossRef]
- Ercolini, D.; Moschetti, G.; Blaiotta, G.; Coppola, S. The potential of a polyphasic PCR-DGGE approach in evaluating microbial diversity of natural whey cultures for water-buffalo Mozzarella cheese production: Bias of culture-dependent and culture-independent analyses. Syst. Appl. Microbiol. 2001, 24, 610–617. [Google Scholar] [CrossRef]
- Yap, M.; Ercolini, D.; Álvarez-Ordóñez, A.; O’Toole, P.W.; O’Sullivan, O.; Cotter, P.D. Next-generation food research: Use of meta-omic approaches for characterizing microbial communities along the food chain. Annu. Rev. Food Sci. Technol. 2022, 13, 361–384. [Google Scholar] [CrossRef]
- Celano, G.; Costantino, G.; Calasso, M.; Randazzo, C.; Minervini, F. Distinctive traits of four apulian traditional Agri-food product (TAP) cheeses manufactured at the same dairy plant. Foods 2022, 11, 425. [Google Scholar] [CrossRef]
- Ullah, M.; Raza, A.; Ye, L.; Yu, Z. Viability and composition validation of commercial probiotic products by selective culturing combined with next-generation sequencing. Microorganisms 2019, 7, 188. [Google Scholar] [CrossRef]
- Shehata, H.R.; Newmaster, S.G. The power of DNA based methods in probiotic authentication. Front. Microbiol. 2023, 14, 1158440. [Google Scholar] [CrossRef] [PubMed]
- Tamang, J.P.; Das, S.; Kharnaior, P.; Pariyar, P.; Thapa, N.; Jo, S.-W.; Yim, E.-J.; Shin, D.-H. Shotgun metagenomics of Cheonggukjang, a fermented soybean food of Korea: Community structure, predictive functionalities and amino acids profile. Food Res. Int. 2021, 151, 110904. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Al-Zahrani, I.A.; Bibi, F.; El Ghany, M.A.; Azhar, E.I. New insights of bacterial communities in fermented vegetables from shotgun metagenomics and identification of antibiotic resistance genes and probiotic bacteria. Food Res. Int. 2022, 157, 111190. [Google Scholar] [CrossRef] [PubMed]
- Emerson, J.B.; Adams, R.I.; Román, C.M.B.; Brooks, B.; Coil, D.A.; Dahlhausen, K.; Ganz, H.H.; Hartmann, E.M.; Hsu, T.; Justice, N.B.; et al. Schrödinger’s microbes: Tools for distinguishing the living from the dead in microbial ecosystems. Microbiome 2017, 5, 86. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Species | Target Gene | Product Encoded by the Gene | References |
|---|---|---|---|
| Streptococcus thermophylus | lacZ | Β-galactosidase enzyme | [21] |
| Lactobacillus delbrueckii subsp. bulgaricus | tuf 16S rRNA | elongation factor Tu 16S rRNA | [22] |
| Levilactobacillus brevis | Gene encoding the aldo/keto reductase of the diketogulonate-reductase family of L. brevis | aldo/keto reductase of the diketogulonate-reductase family of L. brevis | [23] |
| Weissella confusa | lepA gene | GTPbinding protein LepA (Elongation Factor 4) | [24] |
| Lacticaseibacillus casei | 16S rRNA | Ribosomal RNA | [25] |
| Lacticaseibacillus paracasei | |||
| Lacticaseibacillus rhamnosus | |||
| Lactobacillus helveticus | pepC pepN htrA | aminopeptidases C aminopeptidase N, trypsin-like serine protease | [26] |
| Limosilactobacillus fermentum | 16S rRNA | Ribosomal RNA | [27] |
| Lactiplantibacillus plantarum Lactiplantibacillus pentosus Lactiplantibacillus paraplantarum | recA | RecA | [28] |
| 37 Lactobacillus species | 16S-23S rRNA gene | [29] | |
| Leuconostoc (Ln.) mesenteroides, Ln. pseudomesenteroides, Ln. lactis and Ln. citreum | Hsp60 | [30] | |
| Enterococcus (E.) faecalis, E. faecium, E. hirae, and E. casseliflavus | 58 specific molecular targets obtained by pan-genome analysis | [31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fusco, V.; Fanelli, F.; Chieffi, D. Recent and Advanced DNA-Based Technologies for the Authentication of Probiotic, Protected Designation of Origin (PDO) and Protected Geographical Indication (PGI) Fermented Foods and Beverages. Foods 2023, 12, 3782. https://doi.org/10.3390/foods12203782
Fusco V, Fanelli F, Chieffi D. Recent and Advanced DNA-Based Technologies for the Authentication of Probiotic, Protected Designation of Origin (PDO) and Protected Geographical Indication (PGI) Fermented Foods and Beverages. Foods. 2023; 12(20):3782. https://doi.org/10.3390/foods12203782
Chicago/Turabian StyleFusco, Vincenzina, Francesca Fanelli, and Daniele Chieffi. 2023. "Recent and Advanced DNA-Based Technologies for the Authentication of Probiotic, Protected Designation of Origin (PDO) and Protected Geographical Indication (PGI) Fermented Foods and Beverages" Foods 12, no. 20: 3782. https://doi.org/10.3390/foods12203782