
Bioactive Functions of Lipids in the Milk Fat Globule Membrane: A Comprehensive Review


Abstract
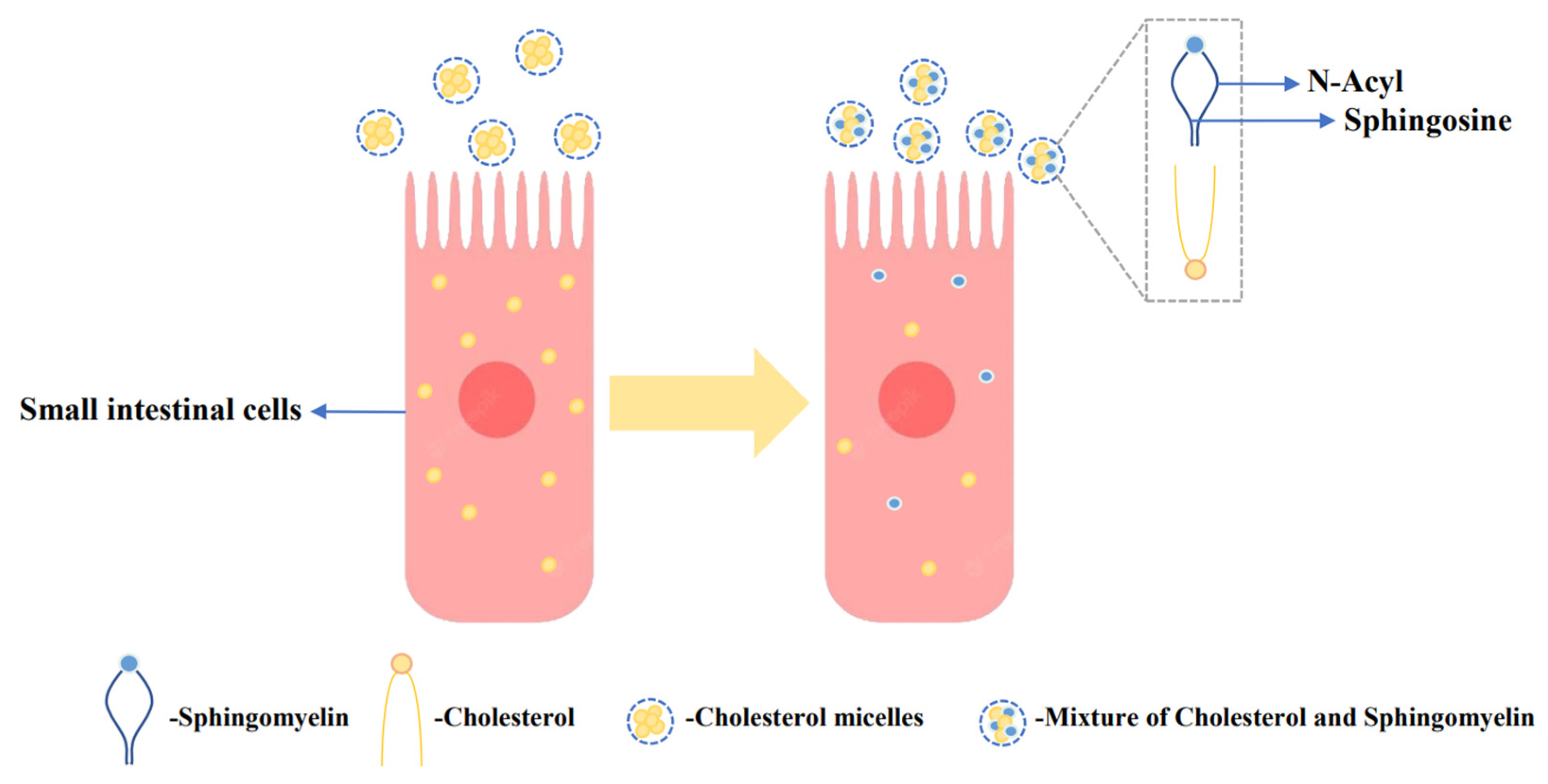
:1. Introduction

{kind=link}
{kind=link}
| Component | MFGM | Whole Milk |
|---|---|---|
| Protein | 25–60% | 1–4% |
| Cholesterol | 2% | 80% |
| PL | 15–30% | 60–70% |
| PC (% PL in MFGM) | 27.4 | 32.7 |
| PE (% PL in MFGM) | 33.0 | 28.5 |
| PS/PI (% PL in MFGM) | 17.8 | 14.1 |
| SM (% PL in MFGM) | 18.8 | 23.0 |
2. Lipids in the MFGM
2.1. Composition and Distribution of Lipids in the MFGM
2.2. Differences of Fatty Acid Composition in the Composition of Milk Fat Globules and MFGMs
3. Characterization of Lipids in the MFGM
4. Various Factors Alter the Lipid Composition of MFGMs
5. Separation of Lipids in MFGMs
6. Phospholipids
6.1. The Promoting Effect of Phospholipid Supplementation on Development
6.2. The Promoting Effect of Phospholipid Supplementation on Memory
6.3. The Promoting Effect of Phospholipid Supplementation on Exercise Performance
6.4. MFGM Phospholipid Supplementation Helps Alleviate Alzheimer’s Disease
6.5. Phospholipids Have a Regulating Effect on Gut Health
6.6. Phospholipids Regulate Cholesterol Metabolism
6.7. Anticancer Effects of Dietary Phospholipids
7. Gangliosides
7.1. Gangliosides Promote Brain Development
7.2. Inhibitory Effect of Gangliosides on Intestinal Pathogenic Microorganisms
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jukkola, A.; Rojas, O.J. Milk fat globules and associated membranes: Colloidal properties and processing effects. Adv. Colloid Interface Sci. 2017, 245, 92–101. [Google Scholar] [CrossRef]
- Wei, W.; Jin, Q.; Wang, X. Human milk fat substitutes: Past achievements and current trends. Prog. Lipid Res. 2019, 74, 69–86. [Google Scholar] [CrossRef]
- Park, Y.W. Bioactive Components in Milk and Dairy Products; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar] [CrossRef]
- Huang, Q.X.; Yang, J.; Hu, M.; Lu, W.; Zhong, K.; Wang, Y.; Yang, G.; Loor, J.J.; Han, L. Milk fat globule membrane proteins are involved in controlling the size of milk fat globules during conjugated linoleic acid–induced milk fat depression. J. Dairy Sci. 2022, 105, 9179–9190. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Jiménez-Flores, R.; Everett, D.W. Bovine milk fat globule membrane proteins are affected by centrifugal washing processes. J. Agric. Food Chem. 2013, 61, 8403–8411. [Google Scholar] [CrossRef] [PubMed]
- Dewettinck, K.; Rombaut, R.; Thienpont, N.; Le, T.T.; Messens, K.; Van Camp, J. Nutritional and technological aspects of milk fat globule membrane material. Int. Dairy J. 2008, 18, 436–457. [Google Scholar] [CrossRef]
- Lopez, C. Intracellular origin of milk fat globules, composition and structure of the milk fat globule membrane highlighting the specific role of sphingomyelin. In Advanced Dairy Chemistry, Volume 2: Lipids; Springer: Berlin/Heidelberg, Germany, 2020; pp. 107–131. [Google Scholar] [CrossRef]
- Ali, F.; Wang, Z.-X. Effect of pasteurization on the enzymatic cross-linking of milk proteins by microbial transglutaminase in view of milk fat globule membrane isolation. Food Biosci. 2021, 43, 101100. [Google Scholar] [CrossRef]
- Fontecha, J.; Brink, L.; Wu, S.; Pouliot, Y.; Visioli, F.; Jiménez-Flores, R. Sources, production, and clinical treatments of milk fat globule membrane for infant nutrition and well-being. Nutrients 2020, 12, 1607. [Google Scholar] [CrossRef]
- Sun, X.; Yu, Z.; Liang, C.; Xie, S.; Wen, J.; Wang, H.; Wang, J.; Yang, Y.; Han, R. Developmental changes in proteins of casein micelles in goat milk using data-independent acquisition-based proteomics methods during the lactation cycle. J. Dairy Sci. 2023, 106, 47–60. [Google Scholar] [CrossRef]
- Yano, M.; Haramizu, S.; Ota, N.; Minegishi, Y.; Shimotoyodome, A. Continuous supplementation of milk fat globule membrane with habitual exercise from a young age improves motor coordination and skeletal muscle function in aged mice. J. Nutr. Sci. Vitaminol. 2019, 65, 405–413. [Google Scholar] [CrossRef]
- Ibitoye, J.O.; Ly-Nguyen, B.; Le, D.N.; Dewettinck, K.; Trzcinski, A.P.; Phan, T.T.Q. Quality of set yogurts made from raw milk and processed milk supplemented with enriched milk fat globule membrane in a two-stage homogenization process. Foods 2021, 10, 1534. [Google Scholar] [CrossRef]
- Qu, X.; Hu, H.; Wang, Y.; Cao, C.; Li, H.; Liu, X.; Yu, J. Proteomics analysis of milk fat globule membrane enriched materials derived from by-products during different stages of milk-fat processing. LWT 2019, 116, 108531. [Google Scholar] [CrossRef]
- Wang, C.; Qiao, X.; Gao, Z.; Jiang, L.; Mu, Z. Advancement on Milk Fat Globule Membrane: Separation, Identification, and Functional Properties. Front. Nutr. 2021, 8, 807284. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.F.; Hogan, S.A.; Tobin, J.; Rasmussen, J.T.; Larsen, L.B.; Wiking, L. Microfiltration of raw milk for production of high-purity milk fat globule membrane material. J. Food Eng. 2020, 276, 109887. [Google Scholar] [CrossRef]
- Holzmüller, W.; Kulozik, U. Isolation of milk fat globule membrane (MFGM) material by coagulation and diafiltration of buttermilk. Int. Dairy J. 2016, 63, 88–91. [Google Scholar] [CrossRef]
- Lopez, C.; Cauty, C.; Guyomarc’h, F. Unraveling the complexity of milk fat globules to tailor bioinspired emulsions providing health benefits: The key role played by the biological membrane. Eur. J. Lipid Sci. Technol. 2019, 121, 1800201. [Google Scholar] [CrossRef]
- Claumarchirant, L.; Matencio, E.; Sanchez-Siles, L.M.; Alegría, A.; Lagarda, M.J. Sterol composition in infant formulas and estimated intake. J. Agric. Food Chem. 2015, 63, 7245–7251. [Google Scholar] [CrossRef]
- Hellhammer, J.; Waladkhani, A.R.; Hero, T.; Buss, C. Effects of milk phospholipid on memory and psychological stress response. Br. Food J. 2010, 57, 183–193. [Google Scholar] [CrossRef]
- Palmano, K.P.; MacGibbon, A.K.; Gunn, C.A.; Schollum, L.M. In vitro and in vivo anti-inflammatory activity of bovine milkfat globule (MFGM)-derived complex lipid fractions. Nutrients 2020, 12, 2089. [Google Scholar] [CrossRef]
- Hernell, O.; Timby, N.; Domellöf, M.; Lönnerdal, B. Clinical benefits of milk fat globule membranes for infants and children. J. Pediatr. 2016, 173, S60–S65. [Google Scholar] [CrossRef]
- Lee, H.; Padhi, E.; Hasegawa, Y.; Larke, J.; Parenti, M.; Wang, A.; Hernell, O.; Lönnerdal, B.; Slupsky, C. Compositional dynamics of the milk fat globule and its role in infant development. Front. Pediatr. 2018, 6, 313. [Google Scholar] [CrossRef]
- Le Huërou-Luron, I.; Lemaire, M.; Blat, S. Health benefits of dairy lipids and MFGM in infant formula. Oléagineux Corps Gras Lipides 2018, 25, D306. [Google Scholar] [CrossRef]
- Sánchez-Juanes, F.; Alonso, J.; Zancada, L.; Hueso, P. Distribution and fatty acid content of phospholipids from bovine milk and bovine milk fat globule membranes. Int. Dairy J. 2009, 19, 273–278. [Google Scholar] [CrossRef]
- Murthy, A.V.R.; Guyomarc’h, F.; Paboeuf, G.; Vie, V.; Lopez, C. Cholesterol strongly affects the organization of lipid monolayers studied as models of the milk fat globule membrane: Condensing effect and change in the lipid domain morphology. Biochim. Biophys. Acta (BBA) Biomembr. 2015, 1848, 2308–2316. [Google Scholar] [CrossRef]
- Liu, Z.; Li, C.; Pryce, J.; Rochfort, S. Comprehensive characterization of bovine milk lipids: Phospholipids, sphingolipids, glycolipids, and ceramides. J. Agric. Food Chem. 2020, 68, 6726–6738. [Google Scholar] [CrossRef]
- Calvo, M.V.; Martín-Hernández, M.C.; García-Serrano, A.; Castro-Gómez, M.P.; Alonso-Miravalles, L.; García-Martín, R.; Megino-Tello, J.; Alonso, L.; Fontecha, J. Comprehensive characterization of neutral and polar lipids of buttermilk from different sources and its milk fat globule membrane isolates. J. Food Compos. Anal. 2020, 86, 103386. [Google Scholar] [CrossRef]
- Ferreiro, T.; Gayoso, L.; Rodríguez-Otero, J. Milk phospholipids: Organic milk and milk rich in conjugated linoleic acid compared with conventional milk. J. Dairy Sci. 2015, 98, 9–14. [Google Scholar] [CrossRef]
- Raza, G.S.; Herzig, K.-H.; Leppäluoto, J. Invited review: Milk fat globule membrane—A possible panacea for neurodevelopment, infections, cardiometabolic diseases, and frailty. J. Dairy Sci. 2021, 104, 7345–7363. [Google Scholar] [CrossRef]
- Alden, K.P.; Dhondt-Cordelier, S.; McDonald, K.L.; Reape, T.J.; Ng, C.K.-Y.; McCabe, P.F.; Leaver, C.J. Sphingolipid long chain base phosphates can regulate apoptotic-like programmed cell death in plants. Biochem. Biophys. Res. Commun. 2011, 410, 574–580. [Google Scholar] [CrossRef]
- Contarini, G.; Povolo, M. Phospholipids in milk fat: Composition, biological and technological significance, and analytical strategies. Int. J. Mol. Sci. 2013, 14, 2808–2831. [Google Scholar] [CrossRef]
- Lopez, C. Milk fat globules enveloped by their biological membrane: Unique colloidal assemblies with a specific composition and structure. Curr. Opin. Colloid Interface Sci. 2011, 16, 391–404. [Google Scholar] [CrossRef]
- Sipione, S.; Monyror, J.; Galleguillos, D.; Steinberg, N.; Kadam, V. Gangliosides in the brain: Physiology, pathophysiology and therapeutic applications. Front. Neurosci. 2020, 14, 1004. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.; Madec, M.-N.; Jimenez-Flores, R. Lipid rafts in the bovine milk fat globule membrane revealed by the lateral segregation of phospholipids and heterogeneous distribution of glycoproteins. Food Chem. 2010, 120, 22–33. [Google Scholar] [CrossRef]
- Murthy, A.V.R.; Guyomarc’h, F.; Lopez, C. Cholesterol decreases the size and the mechanical resistance to rupture of sphingomyelin rich domains, in lipid bilayers studied as a model of the milk fat globule membrane. Langmuir 2016, 32, 6757–6765. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.; Cheng, K.; Perez, J. Thermotropic phase behavior of milk sphingomyelin and role of cholesterol in the formation of the liquid ordered phase examined using SR-XRD and DSC. Chem. Phys. Lipids 2018, 215, 46–55. [Google Scholar] [CrossRef]
- Et-Thakafy, O.; Guyomarc’h, F.; Lopez, C. Lipid domains in the milk fat globule membrane: Dynamics investigated in situ in milk in relation to temperature and time. Food Chem. 2017, 220, 352–361. [Google Scholar] [CrossRef]
- Tanaka, K.; Hosozawa, M.; Kudo, N.; Yoshikawa, N.; Hisata, K.; Shoji, H.; Shinohara, K.; Shimizu, T. The pilot study: Sphingomyelin-fortified milk has a positive association with the neurobehavioural development of very low birth weight infants during infancy, randomized control trial. Brain Dev. 2013, 35, 45–52. [Google Scholar] [CrossRef]
- Oshida, K.; Shimizu, T.; Takase, M.; Tamura, Y.; Shimizu, T.; Yamashiro, Y. Effects of dietary sphingomyelin on central nervous system myelination in developing rats. Pediatr. Res. 2003, 53, 589–593. [Google Scholar] [CrossRef]
- Deoni, S.; Dean III, D.; Joelson, S.; O’Regan, J.; Schneider, N. Early nutrition influences developmental myelination and cognition in infants and young children. Neuroimage 2018, 178, 649–659. [Google Scholar] [CrossRef]
- Veereman-Wauters, G.; Staelens, S.; Rombaut, R.; Dewettinck, K.; Deboutte, D.; Brummer, R.-J.; Boone, M.; Le Ruyet, P. Milk fat globule membrane (INPULSE) enriched formula milk decreases febrile episodes and may improve behavioral regulation in young children. Nutrition 2012, 28, 749–752. [Google Scholar] [CrossRef]
- Schubert, M.; Contreras, C.; Franz, N.; Hellhammer, J. Milk-based phospholipids increase morning cortisol availability and improve memory in chronically stressed men. Nutr. Res. 2011, 31, 413–420. [Google Scholar] [CrossRef]
- Liu, H.; Radlowski, E.C.; Conrad, M.S.; Li, Y.; Dilger, R.N.; Johnson, R.W. Early supplementation of phospholipids and gangliosides affects brain and cognitive development in neonatal piglets. J. Nutr. 2014, 144, 1903–1909. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaka, Y.; Soga, S.; Ota, N.; Yokoyama, K.; Yamada, Y.; Kimura, M. Light rhythmic exercise with dietary milk fat globule membrane improves physical fitness in an elderly Japanese population: A double-blind randomized placebo-controlled trial. Biosci. Biotechnol. Biochem. 2018, 82, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Haramizu, S.; Mori, T.; Yano, M.; Ota, N.; Hashizume, K.; Otsuka, A.; Hase, T.; Shimotoyodome, A. Habitual exercise plus dietary supplementation with milk fat globule membrane improves muscle function deficits via neuromuscular development in senescence-accelerated mice. Springerplus 2014, 3, 339. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Suzuki, T.; Kim, M.; Kojima, N.; Ota, N.; Shimotoyodome, A.; Hase, T.; Hosoi, E.; Yoshida, H. Effects of exercise and milk fat globule membrane (MFGM) supplementation on body composition, physical function, and hematological parameters in community-dwelling frail Japanese women: A randomized double blind, placebo-controlled, follow-up trial. PLoS ONE 2015, 10, e0116256. [Google Scholar] [CrossRef] [PubMed]
- Baliyan, S.; Calvo, M.V.; Piquera, D.; Montero, O.; Visioli, F.; Venero, C.; Fontecha, J. Milk fat globule membrane concentrate as a nutritional supplement prevents age-related cognitive decline in old rats: A lipidomic study of synaptosomes. Food Res. Int. 2023, 163, 112163. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Z.-H.; Huang, S.-L.; Yue, Z.-B.; Yin, X.-S.; Feng, Z.-Q.; Zhang, X.-G.; Song, G.-L. Whey protein powder with milk fat globule membrane attenuates Alzheimer’s disease pathology in 3×Tg-AD mice by modulating neuroinflammation through the peroxisome proliferator-activated receptor γ signaling pathway. J. Dairy Sci. 2023, 106, 5253–5265. [Google Scholar] [CrossRef]
- Billeaud, C.; Puccio, G.; Saliba, E.; Guillois, B.; Vaysse, C.; Pecquet, S.; Steenhout, P. Safety and tolerance evaluation of milk fat globule membrane-enriched infant formulas: A randomized controlled multicenter non-inferiority trial in healthy term infants. Clin. Med. Insights Pediatr. 2014, 8, 51–60. [Google Scholar] [CrossRef]
- Noh, S.K.; Koo, S.I. Milk sphingomyelin is more effective than egg sphingomyelin in inhibiting intestinal absorption of cholesterol and fat in rats. J. Nutr. 2004, 134, 2611–2616. [Google Scholar] [CrossRef]
- Conway, V.; Couture, P.; Richard, C.; Gauthier, S.; Pouliot, Y.; Lamarche, B. Impact of buttermilk consumption on plasma lipids and surrogate markers of cholesterol homeostasis in men and women. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 1255–1262. [Google Scholar] [CrossRef]
- Rosqvist, F.; Smedman, A.; Lindmark-Månsson, H.; Paulsson, M.; Petrus, P.; Straniero, S.; Rudling, M.; Dahlman, I.; Risérus, U. Potential role of milk fat globule membrane in modulating plasma lipoproteins, gene expression, and cholesterol metabolism in humans: A randomized study. Am. J. Clin. Nutr. 2015, 102, 20–30. [Google Scholar] [CrossRef]
- Vors, C.; Joumard-Cubizolles, L.; Lecomte, M.; Combe, E.; Ouchchane, L.; Drai, J.; Raynal, K.; Joffre, F.; Meiller, L.; Le Barz, M. Milk polar lipids reduce lipid cardiovascular risk factors in overweight postmenopausal women: Towards a gut sphingomyelin-cholesterol interplay. Gut 2020, 69, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Snow, D.R.; Jimenez-Flores, R.; Ward, R.E.; Cambell, J.; Young, M.J.; Nemere, I.; Hintze, K.J. Dietary milk fat globule membrane reduces the incidence of aberrant crypt foci in Fischer-344 rats. J. Agric. Food Chem. 2010, 58, 2157–2163. [Google Scholar] [CrossRef] [PubMed]
- Vickers, M.H.; Guan, J.; Gustavsson, M.; Krägeloh, C.U.; Breier, B.H.; Davison, M.; Fong, B.; Norris, C.; McJarrow, P.; Hodgkinson, S.C. Supplementation with a mixture of complex lipids derived from milk to growing rats results in improvements in parameters related to growth and cognition. Nutr. Res. 2009, 29, 426–435. [Google Scholar] [CrossRef]
- Gurnida, D.A.; Rowan, A.M.; Idjradinata, P.; Muchtadi, D.; Sekarwana, N. Association of complex lipids containing gangliosides with cognitive development of 6-month-old infants. Early Hum. Dev. 2012, 88, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, A.J.; Roig-Sagués, A.X.; Zamora, A.; Ferragut, V. 12—High-Pressure Homogenization for Structure Modification. In Innovative Food Processing Technologies; Knoerzer, K., Juliano, P., Smithers, G., Eds.; Woodhead Publishing: Sawston, UK, 2016; pp. 315–344. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, X.; Liu, Y.; Wang, L.; Li, X. Simulated in vitro infant gastrointestinal digestion of infant formulas containing different fat sources and human milk: Differences in lipid profiling and free fatty acid release. J. Agric. Food Chem. 2021, 69, 6799–6809. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.; Briard-Bion, V.; Menard, O.; Rousseau, F.; Pradel, P.; Besle, J.-M. Phospholipid, sphingolipid, and fatty acid compositions of the milk fat globule membrane are modified by diet. J. Agric. Food Chem. 2008, 56, 5226–5236. [Google Scholar] [CrossRef]
- Ménard, O.; Ahmad, S.; Rousseau, F.; Briard-Bion, V.; Gaucheron, F.; Lopez, C. Buffalo vs. cow milk fat globules: Size distribution, zeta-potential, compositions in total fatty acids and in polar lipids from the milk fat globule membrane. Food Chem. 2010, 120, 544–551. [Google Scholar] [CrossRef]
- Lopez, C.; Briard-Bion, V.; Ménard, O. Polar lipids, sphingomyelin and long-chain unsaturated fatty acids from the milk fat globule membrane are increased in milks produced by cows fed fresh pasture based diet during spring. Food Res. Int. 2014, 58, 59–68. [Google Scholar] [CrossRef]
- Fauquant, C.; Briard, V.; Leconte, N.; Michalski, M.C. Differently sized native milk fat globules separated by microfiltration: Fatty acid composition of the milk fat globule membrane and triglyceride core. Eur. J. Lipid Sci. Technol. 2005, 107, 80–86. [Google Scholar] [CrossRef]
- Fong, B.Y.; Norris, C.S.; MacGibbon, A.K. Protein and lipid composition of bovine milk-fat-globule membrane. Int. Dairy J. 2007, 17, 275–288. [Google Scholar] [CrossRef]
- Tavazzi, I.; Fontannaz, P.; Lee, L.Y.; Giuffrida, F. Quantification of glycerophospholipids and sphingomyelin in human milk and infant formula by high performance liquid chromatography coupled with mass spectrometer detector. J. Chromatogr. B 2018, 1072, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.-Q.; Guo, Z.; Huang, J.-H.; Jin, Q.-Z.; Cheong, L.-Z.; Wang, X.-G.; Xu, X.-B. Human milk fat globules from different stages of lactation: A lipid composition analysis and microstructure characterization. J. Agric. Food Chem. 2012, 60, 7158–7167. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Guo, Z.; Jin, Q.; Huang, J.; Cheong, L.; Xu, X.; Wang, X. Composition and microstructure of colostrum and mature bovine milk fat globule membrane. Food Chem. 2015, 185, 362–370. [Google Scholar] [CrossRef]
- Jiang, C.; Zhang, X.; Yu, J.; Yuan, T.; Zhao, P.; Tao, G.; Wei, W.; Wang, X. Comprehensive lipidomic analysis of milk polar lipids using ultraperformance supercritical fluid chromatography-mass spectrometry. Food Chem. 2022, 393, 133336. [Google Scholar] [CrossRef]
- George, A.D.; Gay, M.C.; Selvalatchmanan, J.; Torta, F.; Bendt, A.K.; Wenk, M.R.; Murray, K.; Wlodek, M.E.; Geddes, D.T. Healthy Breastfeeding Infants Consume Different Quantities of Milk Fat Globule Membrane Lipids. Nutrients 2021, 13, 2951. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.H.; Wei, W.; Abed, S.M.; Korma, S.A.; Mousa, A.H.; Hassan, H.M.; Jin, Q.; Wang, X. Impact of technological processes on buffalo and bovine milk fat crystallization behavior and milk fat globule membrane phospholipids profile. Lwt 2018, 90, 424–432. [Google Scholar] [CrossRef]
- Brink, L.R.; Herren, A.W.; McMillen, S.; Fraser, K.; Agnew, M.; Roy, N.; Lönnerdal, B. Omics analysis reveals variations among commercial sources of bovine milk fat globule membrane. J. Dairy Sci. 2020, 103, 3002–3016. [Google Scholar] [CrossRef]
- Yao, Y.; Zhao, G.; Xiang, J.; Zou, X.; Jin, Q.; Wang, X. Lipid composition and structural characteristics of bovine, caprine and human milk fat globules. Int. Dairy J. 2016, 56, 64–73. [Google Scholar] [CrossRef]
- Zou, X.; Huang, J.; Jin, Q.; Guo, Z.; Liu, Y.; Cheong, L.; Xu, X.; Wang, X. Lipid composition analysis of milk fats from different mammalian species: Potential for use as human milk fat substitutes. J. Agric. Food Chem. 2013, 61, 7070–7080. [Google Scholar] [CrossRef]
- Luo, J.; Huang, Z.; Liu, H.; Zhang, Y.; Ren, F. Yak milk fat globules from the Qinghai-Tibetan Plateau: Membrane lipid composition and morphological properties. Food Chem. 2018, 245, 731–737. [Google Scholar] [CrossRef]
- Agyare, A.N.; Liang, Q. Nutrition of yak milk fat–Focusing on milk fat globule membrane and fatty acids. J. Funct. Foods 2021, 83, 104404. [Google Scholar] [CrossRef]
- Thum, C.; Wall, C.; Day, L.; Szeto, I.M.; Li, F.; Yan, Y.; Barnett, M.P. Changes in human milk fat globule composition throughout lactation: A review. Front. Nutr. 2022, 9, 835856. [Google Scholar] [CrossRef] [PubMed]
- Mesilati-Stahy, R.; Argov-Argaman, N. The relationship between size and lipid composition of the bovine milk fat globule is modulated by lactation stage. Food Chem. 2014, 145, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Fauquant, C.; Briard-Bion, V.; Leconte, N.; Guichardant, M.; Michalski, M.C. Membrane phospholipids and sterols in microfiltered milk fat globules. Eur. J. Lipid Sci. Technol. 2007, 109, 1167–1173. [Google Scholar] [CrossRef]
- Lu, J.; Argov-Argaman, N.; Anggrek, J.; Boeren, S.; van Hooijdonk, T.; Vervoort, J.; Hettinga, K.A. The protein and lipid composition of the membrane of milk fat globules depends on their size. J. Dairy Sci. 2016, 99, 4726–4738. [Google Scholar] [CrossRef] [PubMed]
- Mesilati-Stahy, R.; Mida, K.; Argov-Argaman, N. Size-dependent lipid content of bovine milk fat globule and membrane phospholipids. J. Agric. Food Chem. 2011, 59, 7427–7435. [Google Scholar] [CrossRef]
- Truong, T.; Palmer, M.; Bansal, N.; Bhandari, B.; Truong, T.; Palmer, M.; Bansal, N.; Bhandari, B. Size-Dependent Variations in Fatty Acid Composition and Lipid Content of Native Milk Fat Globules. In Effect of Milk Fat Globule Size on the Physical Functionality of Dairy Products; Springer: Berlin/Heidelberg, Germany, 2016; pp. 31–34. [Google Scholar] [CrossRef]
- Lopez, C. Chapter 20—Valorization of dairy by-products for functional and nutritional applications: Recent trends toward the milk fat globule membrane. In Valorization of Agri-Food Wastes and By-Products; Bhat, R., Ed.; Academic Press: Cambridge, MA, USA, 2021; pp. 415–423. [Google Scholar] [CrossRef]
- Pimentel, L.; Gomes, A.; Pintado, M.; Rodríguez-Alcalá, L.M. Isolation and analysis of phospholipids in dairy foods. J. Anal. Methods Chem. 2016, 2016, 9827369. [Google Scholar] [CrossRef]
- Gallier, S.; Gragson, D.; Cabral, C.; Jiménez-Flores, R.; Everett, D.W. Composition and fatty acid distribution of bovine milk phospholipids from processed milk products. J. Agric. Food Chem. 2010, 58, 10503–10511. [Google Scholar] [CrossRef]
- Cheng, S.; Rathnakumar, K.; Martínez-Monteagudo, S.I. Extraction of dairy phospholipids using switchable solvents: A feasibility study. Foods 2019, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Rathnakumar, K.; Ortega-Anaya, J.; Jimenez-Flores, R.; Martínez-Monteagudo, S.I. Understanding the switchable solvent extraction of phospholipids from dairy byproducts. Food Bioprod. Process. 2021, 126, 175–183. [Google Scholar] [CrossRef]
- Costa, M.R.; Elias-Argote, X.E.; Jiménez-Flores, R.; Gigante, M.L. Use of ultrafiltration and supercritical fluid extraction to obtain a whey buttermilk powder enriched in milk fat globule membrane phospholipids. Int. Dairy J. 2010, 20, 598–602. [Google Scholar] [CrossRef]
- Price, N.; Fei, T.; Clark, S.; Wang, T. Extraction of phospholipids from a dairy by-product (whey protein phospholipid concentrate) using ethanol. J. Dairy Sci. 2018, 101, 8778–8787. [Google Scholar] [CrossRef] [PubMed]
- Brink, L.R.; Lönnerdal, B. The role of milk fat globule membranes in behavior and cognitive function using a suckling rat pup supplementation model. J. Nutr. Biochem. 2018, 58, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Moukarzel, S.; Dyer, R.A.; Garcia, C.; Wiedeman, A.M.; Boyce, G.; Weinberg, J.; Keller, B.O.; Elango, R.; Innis, S.M. Milk fat globule membrane supplementation in formula-fed rat pups improves reflex development and may alter brain lipid composition. Sci. Rep. 2018, 8, 15277. [Google Scholar] [CrossRef] [PubMed]
- Schneider, N.; Hauser, J.; Oliveira, M.; Cazaubon, E.; Mottaz, S.C.; O’Neill, B.V.; Steiner, P.; Deoni, S.C. Sphingomyelin in brain and cognitive development: Preliminary data. Eneuro 2019, 6. [Google Scholar] [CrossRef]
- Goñi, F.M. Sphingomyelin: What is it good for? Biochem. Biophys. Res. Commun. 2022, 633, 23–25. [Google Scholar] [CrossRef]
- Guan, J.; MacGibbon, A.; Fong, B.; Zhang, R.; Liu, K.; Rowan, A.; McJarrow, P. Long-term supplementation with beta serum concentrate (BSC), a complex of milk lipids, during post-natal brain development improves memory in rats. Nutrients 2015, 7, 4526–4541. [Google Scholar] [CrossRef]
- Albi, E.; Arcuri, C.; Kobayashi, T.; Tomishige, N.; Dei Cas, M.; Paroni, R.; Signorelli, P.; Cerquiglini, L.; Troiani, S.; Gizzi, C. Sphingomyelin in human breast milk might be essential for the hippocampus maturation. Front. Biosci. 2022, 27, 247. [Google Scholar] [CrossRef]
- Fraser, K.; Ryan, L.; Dilger, R.N.; Dunstan, K.; Armstrong, K.; Peters, J.; Stirrat, H.; Haggerty, N.; MacGibbon, A.K.; Dekker, J. Impacts of formula supplemented with milk fat globule membrane on the neurolipidome of brain regions of piglets. Metabolites 2022, 12, 689. [Google Scholar] [CrossRef]
- Jäger, R.; Purpura, M.; Kingsley, M. Phospholipids and sports performance. J. Int. Soc. Sports Nutr. 2007, 4, 5. [Google Scholar] [CrossRef]
- Mairbäurl, H. Red blood cells in sports: Effects of exercise and training on oxygen supply by red blood cells. Front. Physiol. 2013, 4, 332. [Google Scholar] [CrossRef]
- Rajan, K.B.; Weuve, J.; Barnes, L.L.; McAninch, E.A.; Wilson, R.S.; Evans, D.A. Population estimate of people with clinical Alzheimer’s disease and mild cognitive impairment in the United States (2020–2060). Alzheimer’s Dement. 2021, 17, 1966–1975. [Google Scholar] [CrossRef]
- Kosicek, M.; Hecimovic, S. Phospholipids and Alzheimer’s disease: Alterations, mechanisms and potential biomarkers. Int. J. Mol. Sci. 2013, 14, 1310–1322. [Google Scholar] [CrossRef]
- Schipper, L.; van Dijk, G.; Broersen, L.M.; Loos, M.; Bartke, N.; Scheurink, A.J.W.; van der Beek, E.M. A Postnatal Diet Containing Phospholipids, Processed to Yield Large, Phospholipid-Coated Lipid Droplets, Affects Specific Cognitive Behaviors in Healthy Male Mice123. J. Nutr. 2016, 146, 1155–1161. [Google Scholar] [CrossRef]
- Lee, H.; Slupsky, C.M.; Heckmann, A.B.; Christensen, B.; Peng, Y.; Li, X.; Hernell, O.; Lönnerdal, B.; Li, Z. Milk fat globule membrane as a modulator of infant metabolism and gut microbiota: A formula supplement narrowing the metabolic differences between breastfed and formula-fed infants. Mol. Nutr. Food Res. 2021, 65, 2000603. [Google Scholar] [CrossRef]
- Herrmann, F.; Nieto-Ruiz, A.; Sepúlveda-Valbuena, N.; Miranda, M.T.; Diéguez, E.; Jiménez, J.; De-Castellar, R.; García-Ricobaraza, M.; García-Santos, J.A.; Bermúdez, M.G. Infant formula enriched with milk fat globule membrane, long-chain polyunsaturated fatty acids, synbiotics, gangliosides, nucleotides and sialic acid reduces infections during the first 18 months of life: The COGNIS study. J. Funct. Foods 2021, 83, 104529. [Google Scholar] [CrossRef]
- Ramiro-Cortijo, D.; Singh, P.; Liu, Y.; Medina-Morales, E.; Yakah, W.; Freedman, S.D.; Martin, C.R. Breast milk lipids and fatty acids in regulating neonatal intestinal development and protecting against intestinal injury. Nutrients 2020, 12, 534. [Google Scholar] [CrossRef] [PubMed]
- Zavaleta, N.; Kvistgaard, A.S.; Graverholt, G.; Respicio, G.; Guija, H.; Valencia, N.; Lönnerdal, B. Efficacy of an MFGM-enriched complementary food in diarrhea, anemia, and micronutrient status in infants. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 561–568. [Google Scholar] [CrossRef]
- Ortega-Anaya, J.; Marciniak, A.; Jiménez-Flores, R. Milk fat globule membrane phospholipids modify adhesion of Lactobacillus to mucus-producing Caco-2/Goblet cells by altering the cell envelope. Food Res. Int. 2021, 146, 110471. [Google Scholar] [CrossRef] [PubMed]
- Cichosz, G.; Czeczot, H.; Bielecka, M. The anticarcinogenic potential of milk fat. Ann. Agric. Environ. Med. 2020, 27, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Hajar, R. Risk factors for coronary artery disease: Historical perspectives. Heart Views Off. J. Gulf Heart Assoc. 2017, 18, 109. [Google Scholar] [CrossRef] [PubMed]
- Bruno, R.S.; Pokala, A.; Torres-Gonzalez, M.; Blesso, C.N. Cardiometabolic health benefits of dairy-milk polar lipids. Nutr. Rev. 2021, 79 (Suppl. S2), 16–35. [Google Scholar] [CrossRef] [PubMed]
- Norris, G.H.; Milard, M.; Michalski, M.-C.; Blesso, C.N. Protective properties of milk sphingomyelin against dysfunctional lipid metabolism, gut dysbiosis, and inflammation. J. Nutr. Biochem. 2019, 73, 108224. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Chen, G.; Ma, M.; Qiu, N.; Zhu, L.; Li, J. Egg-yolk sphingomyelin and phosphatidylcholine attenuate cholesterol absorption in Caco-2 cells. Lipids 2018, 53, 217–233. [Google Scholar] [CrossRef]
- Zanabria, R.; Tellez, A.M.; Griffiths, M.; Corredig, M. Milk fat globule membrane isolate induces apoptosis in HT-29 human colon cancer cells. Food Funct. 2013, 4, 222–230. [Google Scholar] [CrossRef]
- Castro-Gomez, P.; Rodriguez-Alcala, L.M.; Monteiro, K.M.; Ruiz, A.; Carvalho, J.E.; Fontecha, J. Antiproliferative activity of buttermilk lipid fractions isolated using food grade and non-food grade solvents on human cancer cell lines. Food Chem. 2016, 212, 695–702. [Google Scholar] [CrossRef]
- Castro-Gomez, M.P.; Rodriguez-Alcala, L.M.; Calvo, M.V.; Romero, J.; Mendiola, J.A.; Ibanez, E.; Fontecha, J. Total milk fat extraction and quantification of polar and neutral lipids of cow, goat, and ewe milk by using a pressurized liquid system and chromatographic techniques. J. Dairy Sci. 2014, 97, 6719–6728. [Google Scholar] [CrossRef]
- Stoica, C.; Ferreira, A.K.; Hannan, K.; Bakovic, M. Bilayer forming phospholipids as targets for cancer therapy. Int. J. Mol. Sci. 2022, 23, 5266. [Google Scholar] [CrossRef]
- Ryan, J.M.; Rice, G.E.; Mitchell, M.D. The role of gangliosides in brain development and the potential benefits of perinatal supplementation. Nutr. Res. 2013, 33, 877–887. [Google Scholar] [CrossRef]
- Guillermo, R.B.; Yang, P.; Vickers, M.H.; McJarrow, P.; Guan, J. Supplementation with complex milk lipids during brain development promotes neuroplasticity without altering myelination or vascular density. Food Nutr. Res. 2015, 59, 25765. [Google Scholar] [CrossRef]
- Rohrhofer, J.; Zwirzitz, B.; Selberherr, E.; Untersmayr, E. The impact of dietary sphingolipids on intestinal microbiota and gastrointestinal immune homeostasis. Front. Immunol. 2021, 12, 1690. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Garrido, D.; Mills, D.A.; Barile, D. Hydrolysis of milk gangliosides by infant-gut associated bifidobacteria determined by microfluidic chips and high-resolution mass spectrometry. Electrophoresis 2014, 35, 1742–1750. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, J.; Barbera, R.; Matencio, E.; Alegría, A.; Lagarda, M. Gangliosides and sialic acid effects upon newborn pathogenic bacteria adhesion: An in vitro study. Food Chem. 2013, 136, 726–734. [Google Scholar] [CrossRef]
- Park, E.J.; Thomson, A.B.; Clandinin, M.T. Protection of intestinal occludin tight junction protein by dietary gangliosides in lipopolysaccharide-induced acute inflammation. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Schnabl, K.L.; Larsen, B.; Van Aerde, J.E.; Lees, G.; Evans, M.; Belosevic, M.; Field, C.; Thomson, A.; Clandinin, M.T. Gangliosides protect bowel in an infant model of necrotizing enterocolitis by suppressing proinflammatory signals. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 382–392. [Google Scholar] [CrossRef]


| Lipid | Participants | Dose | Time | Results | Reference |
|---|---|---|---|---|---|
| Sphingomyelin | Low-birth-weight preterm babies | 20% of total phospholipids in milk | 18 months | Supplementation of sphingomyelin in milk has a positive effect on the neurobehavioral development of low-birth-weight preterm infants. | [38] |
| Sphingomyelin | Wistar rats | 810 mg/100 g Sphingomyelin/diet | 28 days | Sphingomyelin contributes to myelination in developing rats. | [39] |
| Sphingomyelin and phosphatidylcholine | Children aged 0–5 years | 62 mg/L 85 mg/L | 90 days | Sphingomyelin and phosphatidylcholine have significant effects on neural and cognitive developments. | [40] |
| Phospholipids | Healthy preschool children aged 2.5 to 6 years | 250 mg/100 mL | 6 months | High phospholipid concentration in milk is beneficial to children’s behavior regulation and the frequency of fever is significantly reduced. | [41] |
| Phospholipids | 75 chronically stressed men aged 30 to 51 years | 250 mL fat-reduced cream powder derived from bovine milk with 0.5%, 1% PL/day | 42 days | Supplementation of PL increases the availability of cortisol in subjects and attenuates memory decline. | [42] |
| Phospholipids and gangliosides | Piglets | 0.8 or 2.5% Lacprodan PL-20 | 26 days | Supplementation with gangliosides and phospholipids improves spatial learning in piglets and affects brain development. | [43] |
| Phospholipids and exercise | Seniors aged 71–75 years | 1 g tablet containing 16% of phospholipid MFGMs per day | 8 weeks | Participants taking globular membrane tablets perform better in tapping and stepping. | [44] |
| Phospholipids, sphingolipids, and exercise | 15-week-old male SAMP1 and ICR rats | 356 ± 9 mg/day diet (contain 16.6% phospholipids) | 28 weeks | MFGM combined with exercise can improve muscle function deficits. | [45] |
| Phospholipids and exercise | Older women aged 82–84 years | 1 g milk fat globule membrane tablet per day | 12 weeks | Exercise and phospholipid supplementation may improve frailty in older adults. | [46] |
| Phospholipids | 30 fifteen-week-old Wistar rats | 0.5 g buttermilk cookie/day | 4 months | Buttermilk supplementation alters synaptic membrane lipid composition and delays cognitive decline with age. | [47] |
| Phospholipids | twenty 3 × Tg-AD mice and 10 wild-type mice | 3.4 g whey protein powder/kg/day | 3 months | Supplementation of phospholipid-rich protein powder in the diet can alleviate AD symptoms. | [48] |
| Phospholipids | Infants aged ≤14 days | 647 mg/L | 4 months | Diarrhea, vomiting, ear infections, conjunctivitis, and eczema are significantly reduced in infants fed the milk fat globule membrane phospholipid formula. | [49] |
| Sphingomyelin | Male Sprague Dawley rats | 19.5 ± 1.4% dose | 7 weeks | Compared with egg-origin sphingomyelin, milk-origin sphingomyelin has a stronger effect on inhibiting the absorption of fat and cholesterol in the rat intestinal tract. | [50] |
| Phospholipids | Men and women with serum low-density lipoprotein cholesterol (LDL-C) <5.0 mmol/L | 187.5 mg/day | 8 weeks | The intake of phospholipids reduces cholesterol levels in the body, mainly by inhibiting the absorption of cholesterol in the gut. | [51] |
| Phospholipids | Overweight men and women | 40 g/day | 8 weeks | Milk-derived phospholipids significantly reduce fasting and postprandial plasma cholesterol concentrations. Milk fat enclosed by MFGM does not impair lipoprotein profiles. | [52] |
| Phospholipids | Menopausal women | 0.3, 0.5 g/day | 4 weeks | Phospholipids may reduce specific interactions involved in cholesterol absorption in the gut. | [53] |
| Sphingomyelin | Male Fischer-344 rats | 0.11% w/w | 13 weeks | Diets containing sphingomyelin are protective against colon cancer in Fischer-344 rats. | [54] |
| Sphingomyelin and phosphatidylserine | Healthy men with an average age of 41.5 years | 13.5 g/day | 3 weeks | High doses of phospholipids can dampen the activity and reactivity of the hypothalamic–pituitary–adrenal axis (HPAA) and produce in the subject a blunted psychological stress response. | [19] |
| Ganglioside | Wistar rats | 0.2%, 1.0% CML | 80 days | Dietary gangliosides benefit cognitive development in infants. | [55] |
| Ganglioside | Infants aged 2 to 8 weeks | 11~12 μg/mL | 16 weeks | Formula with increased ganglioside content in the diet is beneficial for cognitive development in healthy infants aged 0–6 months. | [56] |
| Fatty Acid | MFG | MFGM |
|---|---|---|
| Saturated fatty acids | 66.3–73.0% | 55.2–67.0% |
| Unsaturated fatty acids | 27.0–33.7% | 33.0–44.8% |
| Omega-6 unsaturated fatty acids | ||
| C18:2 c9, t11 (CLA) | 0.42–0.92% | 6.81–7.37% |
| C18:2 c9, c12 (n-6) | 1.37–1.59% | 4.13–5.11% |
| C20:3 c8, c11, c14 (n-6) | 0.07–0.08% | 0.38–0.57% |
| C20:4 c5, c8, c11, c14 (n-6) | 0.09–0.10% | <0.04% |
| Omega-3 unsaturated fatty acids | ||
| C18:3 c9, c12, c15 (n-3) | 0.26–0.61% | 0.43–1.65% |
| C20:3 c11, c14, c17 (n-3) | 0.11–0.12% | 0.47–0.56% |
| C20:5 c5, c8, c11, c14, c17 (n-3; EPA) | 0.03–0.04% | 0.13–0.86% |
| C22:5 c4, c7, c10, c13, c16, c19 (n-3; DPA) | 0.06–0.10% | 0.32–0.56% |
| C22:6 c4, c7, c10, c13, c16, c19 (n-3; DHA) | 0.00% | 0.01–0.48% |
| Polar Lipids (%) | Bovine | Goat | Human | Sheep | Yak |
|---|---|---|---|---|---|
| PI + PS | 16.29–18.96 | 3.00–23.40 | 20.81–22.21 | 6.60–16.9 | 15.56 |
| PC | 25.74–33.12 | 27.00–32.00 | 24.39–25.08 | 24.50–30.50 | 23.18 |
| PE | 23.42–33.76 | 20.00–42.00 | 12.48–25.33 | 30.50–43.00 | 28.20 |
| SM | 24.87–25.40 | 16.00–30.00 | 29.28–40.18 | 22.30–28.20 | 33.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, J.; Chen, M.; Li, N.; Han, R.; Yang, Y.; Zheng, N.; Zhao, S.; Zhang, Y. Bioactive Functions of Lipids in the Milk Fat Globule Membrane: A Comprehensive Review. Foods 2023, 12, 3755. https://doi.org/10.3390/foods12203755
Pan J, Chen M, Li N, Han R, Yang Y, Zheng N, Zhao S, Zhang Y. Bioactive Functions of Lipids in the Milk Fat Globule Membrane: A Comprehensive Review. Foods. 2023; 12(20):3755. https://doi.org/10.3390/foods12203755
Chicago/Turabian StylePan, Junyu, Meiqing Chen, Ning Li, Rongwei Han, Yongxin Yang, Nan Zheng, Shengguo Zhao, and Yangdong Zhang. 2023. "Bioactive Functions of Lipids in the Milk Fat Globule Membrane: A Comprehensive Review" Foods 12, no. 20: 3755. https://doi.org/10.3390/foods12203755