
Effect of Thermostable Enzymes Produced by Psychrotrophic Bacteria in Raw Milk on the Quality of Ultra-High Temperature Sterilized Milk
Abstract
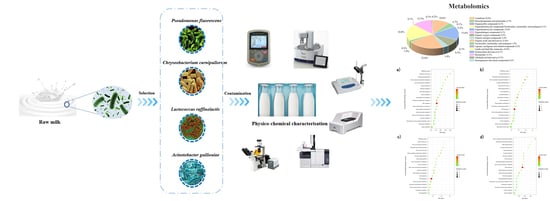
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Strain Cultivation and Screening of Enzyme-Producing Strains
2.3. Quantitative Assessment of Total Proteolytic and Lipolytic Activity
2.4. Thermostability of Proteases and Lipases
2.5. Preparation of Artificially Contaminated Milk Samples
2.6. Physicochemical Properties of Milk Samples
2.6.1. Protein and Fat Content
2.6.2. pH
2.6.3. Taste
2.6.4. Volatile Substances
2.7. Stability of UHT Milk
2.8. Mechanism of UHT Milk Quality Change
2.9. Statistical Analysis
3. Results and Discussion
3.1. Hydrolytic Evaluation of Strains and Heat Resistance of Enzymes
3.2. Physicochemical Properties of Milk Samples
3.2.1. Protein and Fat Content
3.2.2. pH
3.2.3. Taste
3.2.4. Volatile Compounds
3.3. Stability Evaluation
3.4. Metabolomic Analysis on the Changes of UHT Milk Quality
3.4.1. Multivariate Statistical Discrimination of Milk Samples
3.4.2. Metabolites Identified in Milk Samples
3.4.3. KEGG-Enriched Pathways for Differential Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gaucher, I.; Tanguy, G.; Fauquant, J.; Jardin, J.; Rousseau, F.; Robert, B.; Madec, M.-N.; Gaucheron, F. Proteolysis of casein micelles by Pseudomonas fluorescens CNRZ 798 contributes to the destabilisation of UHT milk during its storage. Dairy Sci. Technol. 2011, 91, 413–429. [Google Scholar] [CrossRef]
- Sørhaug, T.; Stepaniak, L. Psychrotrophs and their enzymes in milk and dairy products: Quality aspects. Trends Food Sci. Technol. 1997, 8, 35–41. [Google Scholar] [CrossRef]
- von Neubeck, M.; Baur, C.; Krewinkel, M.; Stoeckel, M.; Kranz, B.; Stressler, T.; Fischer, L.; Hinrichs, J.; Scherer, S.; Wenning, M. Biodiversity of refrigerated raw milk microbiota and their enzymatic spoilage potential. Int. J. Food Microbiol. 2015, 211, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Baglinière, F.; Tanguy, G.; Salgado, R.L.; Jardin, J.; Rousseau, F.; Robert, B.; Harel-Oger, M.; Vanetti, M.C.D.; Gaucheron, F. Ser2 from Serratia liquefaciens L53: A new heat stable protease able to destabilize UHT milk during its storage. Food Chem. 2017, 229, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Stead, D. Microbial lipases: Their characteristics, role in food spoilage and industrial uses. J. Dairy Res. 1986, 53, 481–505. [Google Scholar] [CrossRef]
- De Jonghe, V.; Coorevits, A.; Van Hoorde, K.; Messens, W.; Van Landschoot, A.; De Vos, P.; Heyndrickx, M. Influence of storage conditions on the growth of Pseudomonas species in refrigerated raw milk. Appl. Environ. Microbiol. 2011, 77, 460–470. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, Y.; Xi, X.; Hou, Q.; Xu, H.; Zhao, J.; Li, J.; Bian, Y.; Ma, H.; Wang, Y. Application of PacBio Single Molecule Real-Time (SMRT) sequencing in bacterial source tracking analysis during milk powder production. Food Control 2018, 93, 226–234. [Google Scholar] [CrossRef]
- Dufour, D.; Nicodème, M.; Perrin, C.; Driou, A.; Brusseaux, E.; Humbert, G.; Gaillard, J.-L.; Dary, A. Molecular typing of industrial strains of Pseudomonas spp. isolated from milk and genetical and biochemical characterization of an extracellular protease produced by one of them. Int. J. Food Microbiol. 2008, 125, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Hantsis-Zacharov, E.; Halpern, M. Culturable psychrotrophic bacterial communities in raw milk and their proteolytic and lipolytic traits. Appl. Environ. Microbiol. 2007, 73, 7162–7168. [Google Scholar] [CrossRef]
- Matéos, A.; Guyard-Nicodème, M.; Baglinière, F.; Jardin, J.; Gaucheron, F.; Dary, A.; Humbert, G.; Gaillard, J.-L. Proteolysis of milk proteins by AprX, an extracellular protease identified in Pseudomonas LBSA1 isolated from bulk raw milk, and implications for the stability of UHT milk. Int. Dairy J. 2015, 49, 78–88. [Google Scholar] [CrossRef]
- Stoeckel, M.; Lidolt, M.; Stressler, T.; Fischer, L.; Wenning, M.; Hinrichs, J. Heat stability of indigenous milk plasmin and proteases from Pseudomonas: A challenge in the production of ultra-high temperature milk products. Int. Dairy J. 2016, 61, 250–261. [Google Scholar] [CrossRef]
- Pereira, F.; Luiz, L.; Bruzaroski, S.R.; Poli-Frederico, R.; Fagnani, R.; Santana, E. The effect of cold storage, time and the population of Pseudomonas species on milk lipolysis. Grasas y Aceites 2019, 70, e300. [Google Scholar] [CrossRef]
- Bellassi, P.; Rocchetti, G.; Morelli, L.; Senizza, B.; Lucini, L.; Cappa, F. A milk foodomics investigation into the effect of Pseudomonas fluorescens growth under cold chain conditions. Foods 2021, 10, 1173. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Funct. Genom. 2002, 48, 155–171. [Google Scholar]
- Yang, X.; Guo, X.; Liu, W.; Tian, Y.; Man, C. The complex community structures and seasonal variations of psychrotrophic bacteria in raw milk in Heilongjiang Province, China. LWT Food Sci. Technol. 2020, 134, 110218. [Google Scholar] [CrossRef]
- Júnior, J.R.; De Oliveira, A.; Silva, F.d.G.; Tamanini, R.; De Oliveira, A.; Beloti, V. The main spoilage-related psychrotrophic bacteria in refrigerated raw milk. J. Dairy Sci. 2018, 101, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Sadiq, F.; Liu, T.; Li, Y.; Gu, J.; Yang, H.; He, G. Spoilage potential of psychrotrophic bacteria isolated from raw milk and the thermo-stability of their enzymes. J. Zhejiang Univ. Sci. B 2018, 19, 630–642. [Google Scholar] [CrossRef] [PubMed]
- GB 5009.5-2016; National Food Safety Standard-Determination of Protein in Foods. CFDA: Beijing, China, 2016.
- GB 5009.6-2016; National Food Safety Standard-Determination of Fat in Foods. CFDA: Beijing, China, 2016.
- Fava, L.W.; Serpa, P.; Külkamp-Guerreiro, I.; Pinto, A.T. Evaluation of viscosity and particle size distribution of fresh, chilled and frozen milk of Lacaune ewes. Small Rumin. Res. 2013, 113, 247–250. [Google Scholar] [CrossRef]
- Nrva, F.; Mda, F.; Gb, E.; Eap, C.; Trybd, F.; Nda, D. Biodiversity of culturable psychrotrophic microbiota in raw milk attributable to refrigeration conditions, seasonality and their spoilage potential. Int. Dairy J. 2016, 57, 80–90. [Google Scholar]
- Sun, T.; Wang, X.; Cong, P.; Xu, J.; Xue, C. Mass spectrometry-based lipidomics in food science and nutritional health: A comprehensive review. Compr. Rev. Food Sci. Food Saf. Food Saf. 2020, 19, 2530–2558. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.; Meng, Z.; Zhang, L.; Cui, Y.; Han, X.; Yi, H. The diversity and proteolytic properties of psychrotrophic bacteria in raw cows’ milk from North China. Int. Dairy J. 2017, 66, 34–41. [Google Scholar] [CrossRef]
- Khusniati, T.; Normalia, I. Protease Activities of Semi-purified Pseudomonas fluorescensin Protein Degradation of Pasteurized Milk at Storage. J. Biol. Res. 2011, 17, 91–95. [Google Scholar] [CrossRef]
- Lu, R.; Stevenson, C.D.; Guck, S.E.; Pillsbury, L.A.; Ismail, B.; Hayes, K.D. Effect of various heat treatments on plasminogen activation in bovine milk during refrigerated storage. Int. J. Food Sci. Technol. 2009, 44, 681–687. [Google Scholar] [CrossRef]
- Rauh, V.M.; Sundgren, A.; Bakman, M.; Ipsen, R.; Paulsson, M.; Larsen, L.B.; Hammershøj, M. Plasmin activity as a possible cause for age gelation in UHT milk produced by direct steam infusion. Int. Dairy J. 2014, 38, 199–207. [Google Scholar] [CrossRef]
- Bayaga, C.; Tanguilig, K.; Aba, R.; Pico, M.; Gabriel, A. Culturable micro-organisms in human milk were found to be associated with maternal weight, diet and age during early lactation. J. Appl. Microbiol. 2021, 131, 925–937. [Google Scholar] [CrossRef]
- Burbrink, C.N.; Hayes, K.D. Effect of thermal treatment on the activation of bovine plasminogen. Int. Dairy J. 2006, 16, 580–585. [Google Scholar] [CrossRef]
- Ismail, B.; Nielsen, S.S. Invited review: Plasmin protease in milk: Current knowledge and relevance to dairy industry. J. Dairy Sci. 2010, 93, 4999–5009. [Google Scholar] [CrossRef]
- Zhang, C.; Bijl, E.; Muis, K.; Hettinga, K. Stability of fat globules in UHT milk during proteolysis by the AprX protease from Pseudomonas fluorescens and by plasmin. J. Dairy Sci. 2020, 103, 179–190. [Google Scholar] [CrossRef]
- Baglinière, F.; Matéos, A.; Tanguy, G.; Jardin, J.; Briard-Bion, V.; Rousseau, F.; Robert, B.; Beaucher, E.; Gaillard, J.L.; Amiel, C.; et al. Proteolysis of ultra high temperature-treated casein micelles by AprX enzyme from Pseudomonas fluorescens F induces their destabilisation. Int. Dairy J. 2013, 31, 55–61. [Google Scholar] [CrossRef]
- Vacheyrou, M.; Normand, A.C.; Guyot, P.; Cassagne, C.; Piarroux, R.; Bouton, Y. Cultivable microbial communities in raw cow milk and potential transfers from stables of sixteen French farms. Int. J. Food Microbiol. 2011, 146, 253–262. [Google Scholar] [CrossRef]
- D’Incecco, P.; Brasca, M.; Rosi, V.; Morandi, S.; Ferranti, P.; Picariello, G.; Pellegrino, L. Bacterial proteolysis of casein leading to UHT milk gelation: An applicative study. Food Chem. 2019, 292, 217–226. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Beresford, T.; Ross, R.; Fitzgerald, G.; Cotter, P. Molecular approaches to analysing the microbial composition of raw milk and raw milk cheese. Int. J. Food Microbiol. 2011, 150, 81–94. [Google Scholar] [CrossRef]
- Zhang, C.; Bijl, E.; Hettinga, K. Destabilization of UHT milk by protease AprX from Pseudomonas fluorescens and plasmin. Food Chem. 2018, 263, 127–134. [Google Scholar] [CrossRef]
- Decimo, M.; Morandi, S.; Silvetti, T.; Brasca, M. Characterization of Gram-negative psychrotrophic bacteria isolated from Italian bulk tank milk. J. Food Sci. 2014, 79, M2081–M2090. [Google Scholar] [CrossRef] [PubMed]
- Stoeckel, M.; Lidolt, M.; Achberger, V.; Glück, C.; Krewinkel, M.; Stressler, T.; Von Neubeck, M.; Wenning, M.; Scherer, S.; Fischer, L. Growth of Pseudomonas weihenstephanensis, Pseudomonas proteolytica and Pseudomonas sp. in raw milk: Impact of residual heat-stable enzyme activity on stability of UHT milk during shelf-life. Int. Dairy J. 2016, 59, 20–28. [Google Scholar] [CrossRef]
- Wang, Y.; Han, X.; Chen, X.; Deng, Y. Potential harmful of extracellular proteases secreted by Pseudomonas fluorescens W3 on milk quality. J. Food Process. Preserv. 2021, 45, e15192. [Google Scholar] [CrossRef]
- Jo, Y.; Benoist, D.M.; Barbano, D.M.; Drake, M.A. Flavor and flavor chemistry differences among milks processed by high-temperature, short-time pasteurization or ultra-pasteurization. J. Dairy Sci. 2018, 101, 3812–3828. [Google Scholar] [CrossRef]
- Jensen, S.; Jansson, T.; Eggers, N.; Clausen, M.R.; Larsen, L.B.; Jensen, H.B.; Ray, C.; Sundgren, A.; Andersen, H.J.; Bertram, H.C. Storage-induced changes in the sensory characteristics and volatiles of conventional and lactose-hydrolyzed UHT processed milk. Eur. Food Res. Technol. 2015, 240, 1247–1257. [Google Scholar] [CrossRef]
- Boor, K.; Wiedmann, M.; Murphy, S.; Alcaine, S. A 100-Year Review: Microbiology and safety of milk handling. J. Dairy Sci. 2017, 100, 9933–9951. [Google Scholar] [CrossRef]
- Angelidis, A.; Tsiota, S.; Pexara, A.; Govaris, A. The microbiological quality of pasteurized milk sold by automatic vending machines. Lett. Appl. Microbiol. 2016, 62, 472–479. [Google Scholar] [CrossRef]
- Tran, P.N.; Brown, S.H.; Mitchell, T.W.; Matuschewski, K.; McMillan, P.J.; Kirk, K.; Dixon, M.W.A.; Maier, A.G. A female gametocyte-specific ABC transporter plays a role in lipid metabolism in the malaria parasite. Nat. Commun. 2014, 5, 4773. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, C.; Jiang, Q.; Yin, Y. Butyrate in Energy Metabolism: There Is Still More to Learn. Trends Endocrinol. Metab. TEM 2021, 32, 159–169. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Control Group | Adding P. fluorescens | Adding C. carnipullorum | Adding L. raffinoltis | Adding A. guillouiae | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Kind | Relative Concentration (%) | Kind | Relative Concentration (%) | Kind | Relative Concentration (%) | Kind | Relative Concentration (%) | Kind | Relative Concentration (%) | |
| Alkanes | 6 | 14.41 | 15 | 17.43 | 15 | 16.00 | 8 | 15.30 | 9 | 6.35 |
| Ketone | 4 | 77.07 | 6 | 68.11 | 8 | 69.83 | 6 | 69.30 | 7 | 89.77 |
| Aldehydes | - | - | 1 | 0.25 | 1 | 0.14 | - | - | - | - |
| Acid | - | - | 3 | 0.48 | 2 | 0.22 | 2 | 0.08 | 3 | 0.15 |
| Alcohol | - | - | 2 | 3.52 | 4 | 4.10 | - | - | 2 | 0.42 |
| Esters | - | - | - | - | 1 | 0.13 | - | - | - | - |
| Alkene | 2 | 3.02 | 1 | 3.94 | 1 | 3.31 | 3 | 4.59 | 1 | 1.10 |
| Benzene | 5 | 9.22 | 4 | 6.28 | 4 | 6.13 | 5 | 10.73 | 4 | 2.15 |
| Others | - | - | - | - | - | - | - | - | 1 | 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, X.; Cheng, J.; Qi, X.; Guan, N.; Chen, Q.; Pei, X.; Jiang, Y.; Yang, X.; Man, C. Effect of Thermostable Enzymes Produced by Psychrotrophic Bacteria in Raw Milk on the Quality of Ultra-High Temperature Sterilized Milk. Foods 2023, 12, 3752. https://doi.org/10.3390/foods12203752
Qin X, Cheng J, Qi X, Guan N, Chen Q, Pei X, Jiang Y, Yang X, Man C. Effect of Thermostable Enzymes Produced by Psychrotrophic Bacteria in Raw Milk on the Quality of Ultra-High Temperature Sterilized Milk. Foods. 2023; 12(20):3752. https://doi.org/10.3390/foods12203752
Chicago/Turabian StyleQin, Xue, Jingqi Cheng, Xuehe Qi, Ning Guan, Qing Chen, Xiaoyan Pei, Yujun Jiang, Xinyan Yang, and Chaoxin Man. 2023. "Effect of Thermostable Enzymes Produced by Psychrotrophic Bacteria in Raw Milk on the Quality of Ultra-High Temperature Sterilized Milk" Foods 12, no. 20: 3752. https://doi.org/10.3390/foods12203752