
Measuring the Phytochemical Richness of Meat: Effects of Grass/Grain Finishing Systems and Grapeseed Extract Supplementation on the Fatty Acid and Phytochemical Content of Beef

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design, Animals, and Diets
2.2. Sample Collection
2.3. Proximate Analysis
2.4. Fatty Acid Analysis
2.5. Vitamin E and Mineral Analysis of Beef
2.6. Polyphenolic Profiling
2.7. Statistical Analysis
3. Results
3.1. Nutritional Composition of the Diets
3.2. Fatty Acid and Micronutrient Content of Beef
3.2.1. Fatty Acids
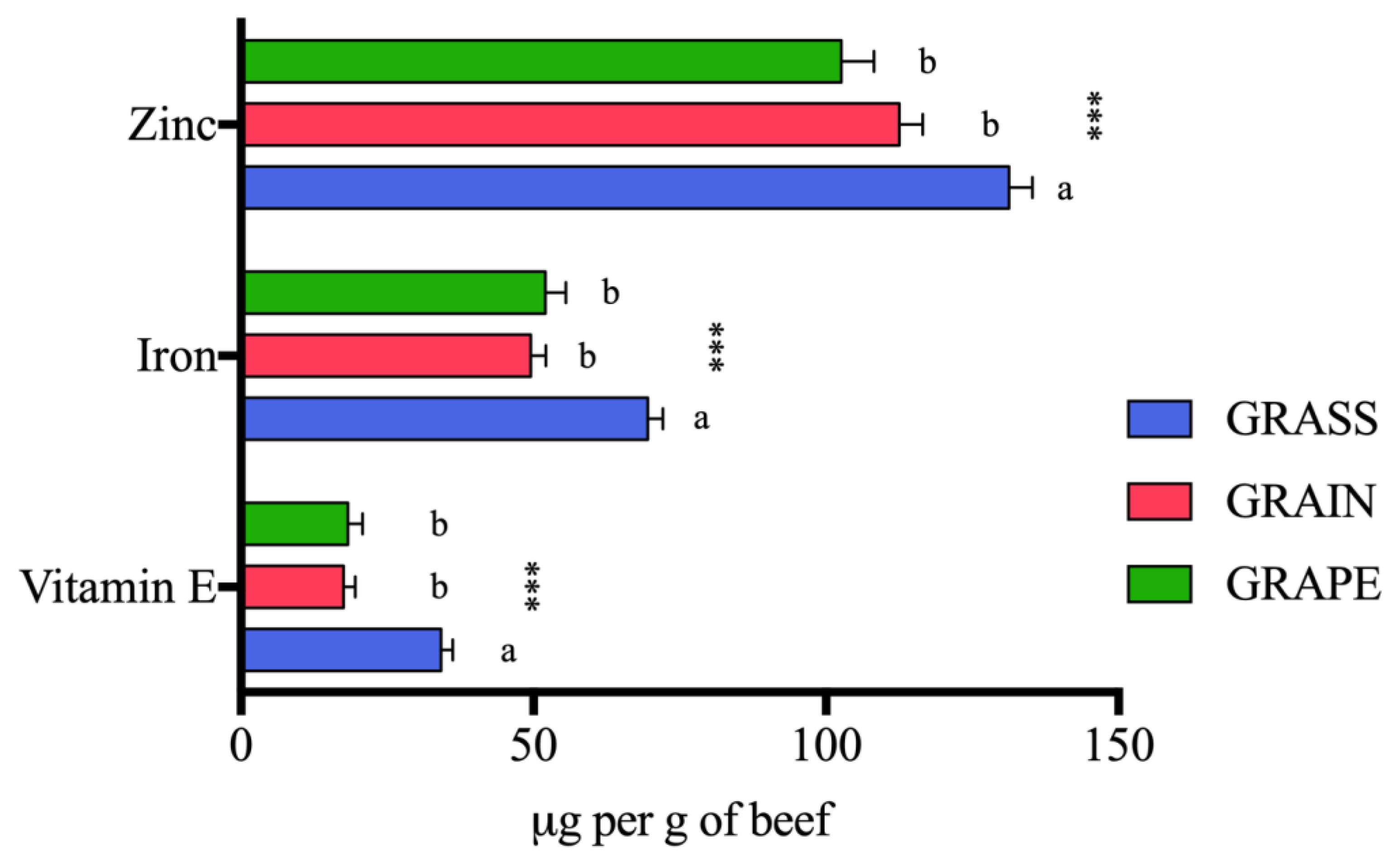
3.2.2. Micronutrients
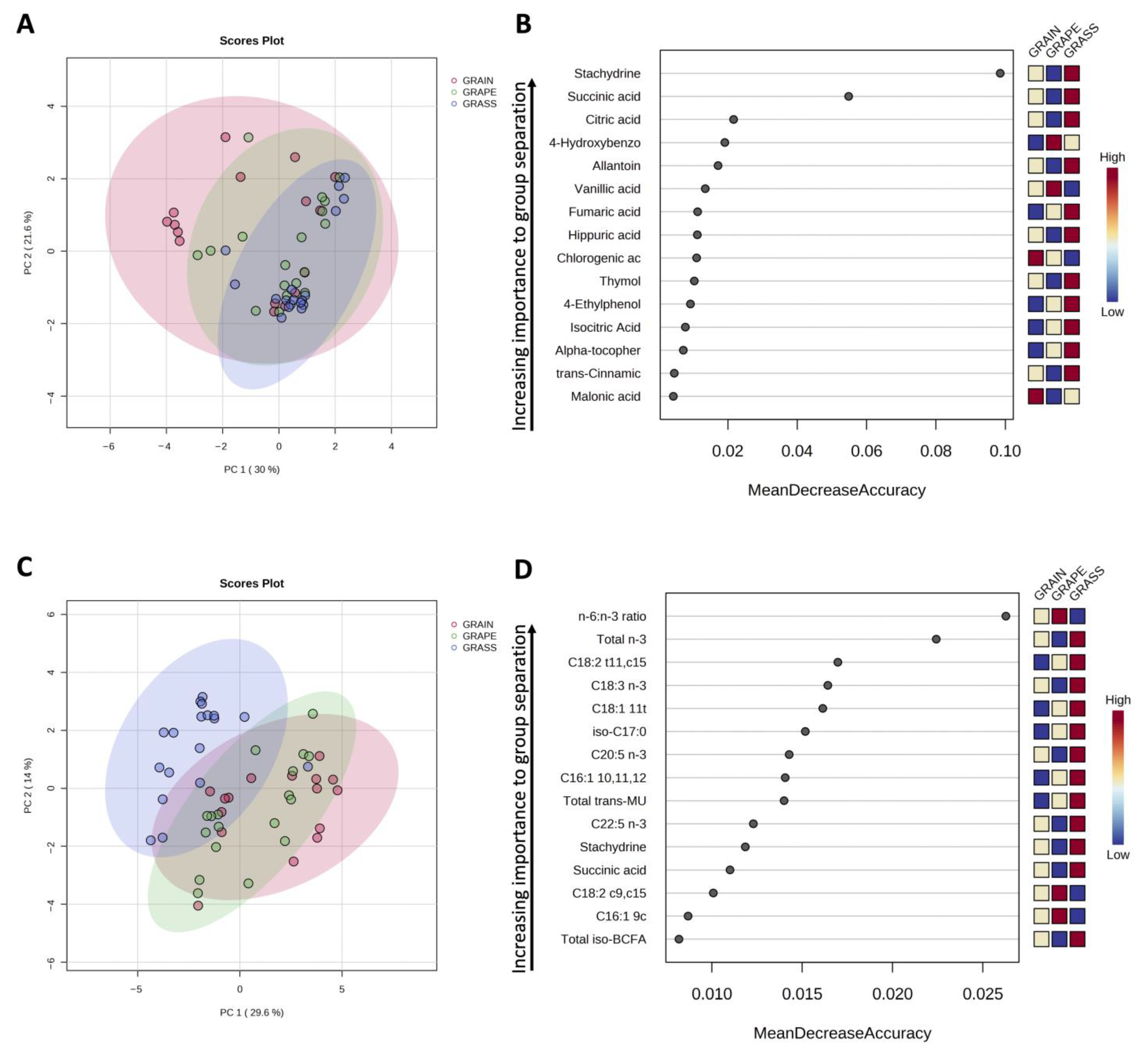
3.3. Phytochemical Profile of Beef
3.3.1. Data Visualization and Identification of Top Discriminating Compounds
3.3.2. Differences in Quantified Phytochemicals in Beef
4. Discussion
4.1. Nutritional Composition of the Diets
4.2. Beef Fatty Acids and Micronutrients
4.2.1. Fatty Acids
4.2.2. Vitamin E, Zinc, and Iron
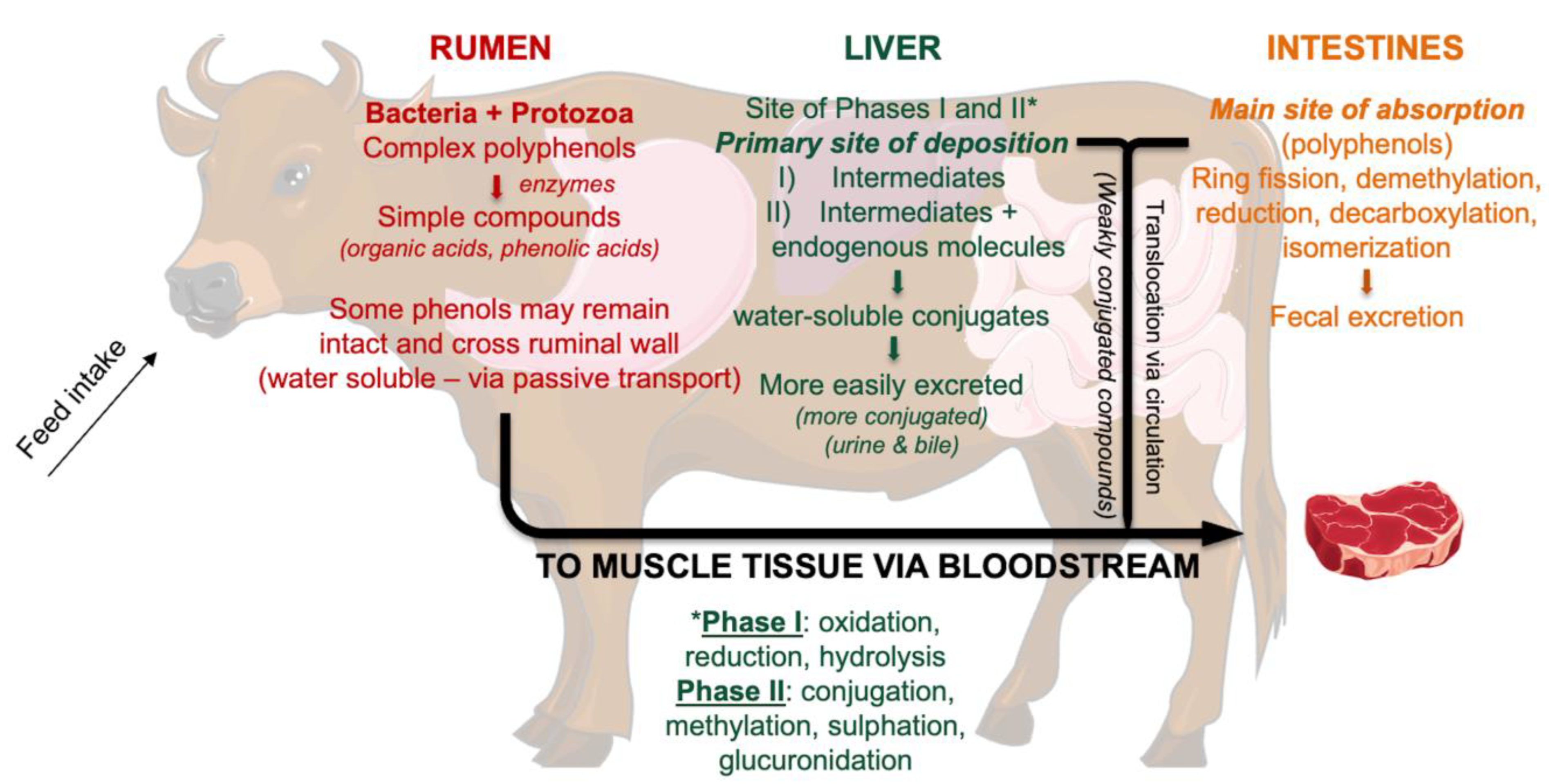
4.3. Phytochemical Profile of Beef

5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Aveyard, P.; Garnett, T.; Hall, J.W.; Key, T.J.; Lorimer, J.; Pierrehumbert, R.T.; Scarborough, P.; Springmann, M.; Jebb, S.A. Meat consumption, health, and the environment. Science 2018, 361, eaam5324. [Google Scholar] [CrossRef]
- Provenza, F.D.; Kronberg, S.L.; Gregorini, P. Is grassfed meat and dairy better for human and environmental health? Front. Nutr. 2019, 6, 26. [Google Scholar] [CrossRef]
- Xue, H.; Mainville, D.; You, W.; Nayga, R.M. Consumer preferences and willingness to pay for grass-fed beef: Empirical evidence from in-store experiments. Food Qual. Prefer. 2010, 21, 857–866. [Google Scholar] [CrossRef]
- Davis, H.; Magistrali, A.; Butler, G.; Stergiadis, S. Nutritional benefits from fatty acids in organic and grass-fed beef. Foods 2022, 11, 646. [Google Scholar] [CrossRef]
- Krusinski, L.; Maciel, I.C.F.; Sergin, S.; Goeden, T.; Schweihofer, J.P.; Singh, S.; Rowntree, J.E.; Fenton, J.I. Fatty acid and micronutrient profile of Longissimus lumborum from Red Angus and Red Angus x Akaushi cattle finished on grass or grain. Foods 2022, 11, 3451. [Google Scholar] [CrossRef]
- Van Elswyk, M.E.; McNeill, S.H. Impact of grass/forage feeding versus grain finishing on beef nutrients and sensory quality: The U.S. experience. Meat Sci. 2014, 96, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Alfaia, C.P.M.; Alves, S.P.; Martins, S.I.V.; Costa, A.S.H.; Fontes, C.M.G.A.; Lemos, J.P.C.; Bessa, R.J.B.; Prates, J.A.M. Effect of the feeding system on intramuscular fatty acids and conjugated linoleic acid isomers of beef cattle, with emphasis on their nutritional value and discriminatory ability. Food Chem. 2009, 114, 939–946. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Mann, N.J.; Sinclair, A.J. Effect of feeding systems on omega-3 fatty acids, conjugated linoleic acid and trans fatty acids in Australian beef cuts: Potential impact on human health. Asia Pac. J. Clin. Nutr. 2006, 15, 21–29. [Google Scholar] [PubMed]
- van Vliet, S.; Provenza, F.D.; Kronberg, S.L. Health-promoting phytonutrients are higher in grass-fed meat and milk. Front. Sustain. Food Syst. 2021, 4, 1–20. [Google Scholar] [CrossRef]
- Duckett, S.K.; Neel, J.P.S.; Fontenot, J.P.; Clapham, W.M. Effects of winter stocker growth rate and finishing system on: III. Tissue proximate, fatty acid, vitamin, and cholesterol content. J. Anim. Sci. 2009, 87, 2961–2970. [Google Scholar] [CrossRef] [PubMed]
- Garcia, P.T.; Pensel, N.A.; Sancho, A.M.; Latimori, N.J.; Kloster, A.M.; Amigone, M.A.; Casal, J.J. Beef lipids in relation to animal breed and nutrition in Argentina. Meat Sci. 2008, 79, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Daley, C.A.; Abbott, A.; Doyle, P.S.; Nader, G.A.; Larson, S. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutr. J. 2010, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Leheska, J.M.; Thompson, L.D.; Howe, J.C.; Hentges, E.; Boyce, J.; Brooks, J.C.; Shriver, B.; Hoover, L.; Miller, M.F. Effects of conventional and grass-feeding systems on the nutrient composition of beef. J. Anim. Sci. 2008, 86, 3575–3585. [Google Scholar] [CrossRef]
- Serra, V.; Salvatori, G.; Pastorelli, G. Dietary polyphenol supplementation in food producing animals: Effects on the quality of derived products. Animals 2021, 11, 401. [Google Scholar] [CrossRef]
- O’Connell, J.E.; Fox, P.F. Significance and applications of phenolic compounds in the production and quality of milk and dairy products: A review. Int. Dairy J. 2001, 11, 103–120. [Google Scholar] [CrossRef]
- Krusinski, L.; Sergin, S.; Jambunathan, V.; Rowntree, J.E.; Fenton, J.I. Attention to the details: How variations in U.S. grass-fed cattle-feed supplementation and finishing date influence human health. Front. Sustain. Food Syst. 2022, 6, 851494. [Google Scholar] [CrossRef]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Afrin, S.; Gasparrini, M.; Reboredo-Rodriguez, P.; Manna, P.P.; Zhang, J.; Lamas, L.B.; Flórez, S.M.; Toyos, P.A.; et al. Phenolic compounds in honey and their associated health benefits: A review. Molecules 2018, 23, 2322. [Google Scholar] [CrossRef]
- Pogorzelska-Nowicka, E.; Atanasov, A.G.; Horbańczuk, J.; Wierzbicka, A. Bioactive compounds in functional meat products. Molecules 2018, 23, 307. [Google Scholar] [CrossRef]
- Kumar, Y.; Yadav, D.N.; Ahmad, T.; Narsaiah, K. Recent trends in the use of natural antioxidants for meat and meat products. Compr. Rev. Food Sci. Food Saf. 2015, 14, 796–812. [Google Scholar] [CrossRef]
- Scalbert, A.; Manach, C.; Morand, C.; Remesy, C.; Jimenez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Braffett, B.H.; Simmens, S.J.; Young, H.A.; Ogden, C.L. Dietary polyphenol intake in US adults and 10-year trends: 2007-2016. J. Acad. Nutr. Diet. 2020, 120, 1821–1833. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, S.; Bain, J.R.; Muehlbauer, M.J.; Provenza, F.D.; Kronberg, S.L.; Pieper, C.F.; Huffman, K.M. A metabolomics comparison of plant-based meat and grass-fed meat indicates large nutritional differences despite comparable Nutrition Facts panels. Sci. Rep. 2021, 11, 13828. [Google Scholar] [CrossRef] [PubMed]
- Gatellier, P.; Mercier, Y.; Renerre, M. Effect of diet finishing mode (pasture or mixed diet) on antioxidant status of Charolais bovine meat. Meat Sci. 2004, 67, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Han, L.; Yu, Q.-L.; Li, R.-R. Color stability and antioxidant capacity of yak meat as affected by feeding with pasture or grain. Can. J. Anim. Sci. 2015, 95, 189–195. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, B.; Liu, C.; Su, R.; Hou, Y.; Yao, D.; Zhao, L.; Su, L.; Jin, Y. Meat quality, fatty acids, volatile compounds, and antioxidant properties of lambs fed pasture versus mixed diet. Food Sci. Nutr. 2019, 7, 2796–2805. [Google Scholar] [CrossRef]
- Lopez-Andres, P.; Luciano, G.; Vasta, V.; Gibson, T.M.; Scerra, M.; Biondi, L.; Priolo, A.; Mueller-Harvey, I. Antioxidant effects of ryegrass phenolics in lamb liver and plasma. Animal 2014, 8, 51–57. [Google Scholar] [CrossRef]
- Krusinski, L.; Maciel, I.C.F.; Sergin, S.; Goeden, T.; Ali, H.; Kesamneni, S.; Jambunathan, V.; Cassida, K.A.; Singh, S.; Medina-Meza, I.G.; et al. Evaluation of fatty acid and antioxidant variation in a complex pasture system as compared to standard cattle feed in the Great Lakes Region. Front. Sustain. Food Syst. 2022, 6, 945080. [Google Scholar] [CrossRef]
- Reynaud, A.; Fraisse, D.; Cornu, A.; Farruggia, A.; Pujos-Guillot, E.; Besle, J.M.; Martin, B.; Lamaison, J.L.; Paquet, D.; Doreau, M.; et al. Variation in content and composition of phenolic compounds in permanent pastures according to botanical variation. J. Agric. Food Chem. 2010, 58, 5485–5494. [Google Scholar] [CrossRef]
- Muñoz-González, I.; Chamorro, S.; Pérez-Jiménez, J.; López-Andrés, P.; Álvarez-Acero, I.; Herrero, A.M.; Nardoia, M.; Brenes, A.; Viveros, A.; Arija, I.; et al. Phenolic metabolites in plasma and thigh meat of chickens supplemented with grape byproducts. J. Agric. Food Chem. 2019, 67, 4463–4471. [Google Scholar] [CrossRef]
- Brenes, A.; Viveros, A.; Goñi, I.; Centeno, C.; Sáyago-Ayerdy, S.G.; Arija, I.; Saura-Calixto, F. Effect of grape pomace concentrate and vitamin E on digestibility of polyphenols and antioxidant activity in chickens. Poult. Sci. 2008, 87, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Krusinski, L.; Maciel, I.C.F.; Sergin, S.; Jambunathan, V.; Garg, E.; Garmyn, A.J.; Singh, S.; Bitler, C.A.; Rowntree, J.E.; Fenton, J.I. Effects of hay, baleage, and soybean hulls waste used as supplemental feeds on the nutritional profile of grass-finished beef. Foods 2022, 11, 3856. [Google Scholar] [CrossRef] [PubMed]
- Niderkorn, V.; Jayanegara, A. Opportunities offered by plant bioactive compounds to improve silage quality, animal health and product quality for sustainable ruminant production: A review. Agronomy 2021, 11, 86. [Google Scholar] [CrossRef]
- Arola-Arnal, A.; Oms-Oliu, G.; Crescenti, A.; del Bas, J.M.; Ras, M.R.; Arola, L.; Caimari, A. Distribution of grape seed flavanols and their metabolites in pregnant rats and their fetuses. Mol. Nutr. Food Res. 2013, 57, 1741–1752. [Google Scholar] [CrossRef]
- Antonini, E.; Torri, L.; Piochi, M.; Cabrino, G.; Meli, M.A.; De Bellis, R. Nutritional, antioxidant and sensory properties of functional beef burgers formulated with chia seeds and goji puree, before and after in vitro digestion. Meat Sci. 2020, 161, 108021. [Google Scholar] [CrossRef]
- Serra, A.; Blade, C.; Arola, L.; Macia, A.; Motilva, M.J. Flavanol metabolites distribute in visceral adipose depots after a long-term intake of grape seed proanthocyanidin extract in rats. Br. J. Nutr. 2013, 110, 1411–1420. [Google Scholar] [CrossRef]
- Delgadillo-Puga, C.; Cuchillo-Hilario, M.; Leon-Ortiz, L.; Ramirez-Rodriguez, A.; Cabiddu, A.; Navarro-Ocana, A.; Morales-Romero, A.M.; Medina-Campos, O.N.; Pedraza-Chaverri, J. Goats’ feeding supplementation with Acacia farnesiana pods and their relationship with milk composition: Fatty acids, polyphenols, and antioxidant activity. Animals 2019, 9, 515. [Google Scholar] [CrossRef]
- Delgadillo-Puga, C.; Cuchillo-Hilario, M. Reviewing the benefits of grazing/browsing semiarid rangeland feed resources and the transference of bioactivity and pro-healthy properties to goat milk and cheese: Obesity, insulin resistance, inflammation and hepatic steatosis prevention. Animals 2021, 11, 2942. [Google Scholar] [CrossRef]
- Besle, J.M.; Viala, D.; Martin, B.; Pradel, P.; Meunier, B.; Berdague, J.L.; Fraisse, D.; Lamaison, J.L.; Coulon, J.B. Ultraviolet-absorbing compounds in milk are related to forage polyphenols. J. Dairy Sci. 2010, 93, 2846–2856. [Google Scholar] [CrossRef]
- van Vliet, S.; Blair, A.D.; Hite, L.M.; Cloward, J.; Ward, R.E.; Kruse, C.; van Wietmarchsen, H.A.; van Eekeren, N.; Kronberg, S.L.; Provenza, F.D. Pasture-finishing of bison improves animal metabolic health and potential health-promoting compounds in meat. J. Anim. Sci. Biotechnol. 2023, 14, 49. [Google Scholar] [CrossRef]
- Maciel, I.C.F.; Schweihofer, J.P.; Fenton, J.I.; Hodbod, J.; McKendree, M.G.S.; Cassida, K.; Rowntree, J.E. Influence of beef genotypes on animal performance, carcass traits, meat quality, and sensory characteristics in grazing or feedlot-finished steers. Transl. Anim. Sci. 2021, 5, txab214. [Google Scholar] [CrossRef]
- Mertens, D.R. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beakers or crucibles: Collaborative study. J. AOAC Int. 2002, 85, 1217–1240. [Google Scholar] [PubMed]
- AOAC. Official Methods of Analysis 2000; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Hach, C.C.; Bowden, B.K.; Kopelove, A.B.; Brayton, S.V. More powerful peroxide Kjeldahl digestion method. J. Assoc. Off. Anal. Chem. 1987, 70, 783–787. [Google Scholar] [CrossRef]
- Sergin, S.; Goeden, T.; Krusinski, L.; Kesamneni, S.; Ali, H.; Bitler, C.A.; Medina-Meza, I.G.; Fenton, J.I. Fatty acid and antioxidant composition of conventional compared to pastured eggs: Characterization of conjugated linoleic acid and branched chain fatty acid isomers in eggs. Food Sci. Technol. 2021, 1, 260–267. [Google Scholar] [CrossRef]
- Bronkema, S.M.; Rowntree, J.E.; Jain, R.; Schweihofer, J.P.; Bitler, C.A.; Fenton, J.I. A nutritional survey of commercially available grass-finished beef. Meat Muscle Biol. 2019, 3, 116–126. [Google Scholar] [CrossRef]
- Jenkins, T.C. Technical note: Common analytical errors yielding inaccurate results during analysis of fatty acids in feed and digesta samples. J. Dairy Sci. 2010, 93, 1170–1174. [Google Scholar] [CrossRef] [PubMed]
- Kramer, J.K.; Hernandez, M.; Cruz-Hernandez, C.; Kraft, J.; Dugan, M.E. Combining results of two GC separations partly achieves determination of all cis and trans 16:1, 18:1, 18:2 and 18:3 except CLA isomers of milk fat as demonstrated using Ag-ion SPE fractionation. Lipids 2008, 43, 259–273. [Google Scholar] [CrossRef]
- Rettenmaier, R.; Schüep, W. Determination of vitamins A and E in liver tissue. Int. J. Vitam. Nutr. Res. 1992, 62, 312–317. [Google Scholar]
- Wahlen, R.; Evans, L.; Turner, J.; Hearn, R. The use of collision/reaction cell ICP-MS for the determination of elements in blood and serum samples. Spectroscopy 2005, 20, 84–89. Available online: https://www.spectroscopyonline.com/view/use-collisionreaction-cell-icp-ms-determination-elements-blood-and-serum-samples (accessed on 21 February 2022).
- Glasser, F.; Doreau, M.; Maxin, G.; Baumont, R. Fat and fatty acid content and composition of forages: A meta-analysis. Anim. Feed Sci. Technol. 2013, 185, 19–34. [Google Scholar] [CrossRef]
- Klopatek, S.C.; Xu, Y.; Yang, X.; Oltjen, J.W.; Vahmani, P. Effects of multiple grass- and grain-fed production systems on beef fatty acid contents and their consumer health implications. Food Sci. Technol. 2022, 2, 712–721. [Google Scholar] [CrossRef]
- Khan, N.A.; Cone, J.W.; Fievez, V.; Hendriks, W.H. Causes of variation in fatty acid content and composition in grass and maize silages. Anim. Feed Sci. Technol. 2012, 174, 36–45. [Google Scholar] [CrossRef]
- Alothman, M.; Hogan, S.A.; Hennessy, D.; Dillon, P.; Kilcawley, K.N.; O’Donovan, M.; Tobin, J.; Fenelon, M.A.; O’Callaghan, T.F. The “grass-fed” milk story: Understanding the impact of pasture feeding on the composition and quality of bovine milk. Foods 2019, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Butler, G. Manipulating dietary PUFA in animal feed: Implications for human health. Proc. Nutr. Soc. 2014, 73, 87–95. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Shingfield, K.J.; Lee, M.R.F.; Scollan, N.D. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems. Anim. Feed Sci. Technol. 2006, 131, 168–206. [Google Scholar] [CrossRef]
- Garavaglia, J.; Markoski, M.M.; Oliveira, A.; Marcadenti, A. Grape seed oil compounds: Biological and chemical actions for health. Nutr. Metab. Insights 2016, 9, 59–64. [Google Scholar] [CrossRef]
- Vinyard, J.R.; Myers, C.A.; Murdoch, G.K.; Rezamand, P.; Chibisa, G.E. Optimum grape pomace proportion in feedlot cattle diets: Ruminal fermentation, total tract nutrient digestibility, nitrogen utilization, and blood metabolites. J. Anim. Sci. 2021, 99, skab044. [Google Scholar] [CrossRef]
- Nudda, A.; Buffa, G.; Atzori, A.S.; Cappai, M.G.; Caboni, P.; Fais, G.; Pulina, G. Small amounts of agro-industrial byproducts in dairy ewes diets affects milk production traits and hematological parameters. Anim. Feed Sci. Technol. 2019, 251, 76–85. [Google Scholar] [CrossRef]
- Arend, F.A.; Murdoch, G.K.; Doumit, M.E.; Chibisa, G.E. Inclusion of grape pomace in finishing cattle diets: Carcass traits, meat quality and fatty acid composition. Animals 2022, 12, 2597. [Google Scholar] [CrossRef]
- Duckett, S.K.; Neel, J.P.S.; Lewis, R.M.; Fontenot, J.P.; Clapham, W.M. Effects of forage species or concentrate finishing on animal performance, carcass and meat quality. J. Anim. Sci. 2013, 91, 1454–1467. [Google Scholar] [CrossRef]
- Realini, C.E.; Duckett, S.K.; Brito, G.W.; Dalla Rizza, M.; De Mattos, D. Effect of pasture vs. concentrate feeding with or without antioxidants on carcass characteristics, fatty acid composition, and quality of Uruguayan beef. Meat Sci. 2004, 66, 567–577. [Google Scholar] [CrossRef]
- Duckett, S.K.; Pratt, S.L.; Pavan, E. Corn oil or corn grain supplementation to steers grazing endophyte-free tall fescue. II. Effects on subcutaneous fatty acid content and lipogenic gene expression. J. Anim. Sci. 2009, 87, 1120–1128. [Google Scholar] [CrossRef]
- Manso, T.; Gallardo, B.; Salva, A.; Guerra-Rivas, C.; Mantecon, A.R.; Lavin, P.; de la Fuente, M.A. Influence of dietary grape pomace combined with linseed oil on fatty acid profile and milk composition. J. Dairy Sci. 2016, 99, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Moate, P.J.; Williams, S.R.; Torok, V.A.; Hannah, M.C.; Ribaux, B.E.; Tavendale, M.H.; Eckard, R.J.; Jacobs, J.L.; Auldist, M.J.; Wales, W.J. Grape marc reduces methane emissions when fed to dairy cows. J. Dairy Sci. 2014, 97, 5073–5087. [Google Scholar] [CrossRef] [PubMed]
- Mapiye, C.; Vahmani, P.; Mlambo, V.; Muchenje, V.; Dzama, K.; Hoffman, L.C.; Dugan, M.E.R. The trans-octadecenoic fatty acid profile of beef: Implications for global food and nutrition security. Food Res. Int. 2015, 76, 992–1000. [Google Scholar] [CrossRef]
- Calder, P.C. Functional roles of fatty acids and their effects on human health. J. Parenter. Enteral Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Chung, K.Y.; Lunt, D.K.; Choi, C.B.; Chae, S.H.; Rhoades, R.D.; Adams, T.H.; Booren, B.; Smith, S.B. Lipid characteristics of subcutaneous adipose tissue and M. longissimus thoracis of Angus and Wagyu steers fed to US and Japanese endpoints. Meat Sci. 2006, 73, 432–441. [Google Scholar] [CrossRef]
- Smith, S.B.; Lunt, D.K.; Chung, K.Y.; Choi, C.B.; Tume, R.K.; Zembayashi, M. Adiposity, fatty acid composition, and delta-9 desaturase activity during growth in beef cattle. Anim. Sci. J. 2006, 77, 478–486. [Google Scholar] [CrossRef]
- Parolini, C. Effects of fish n-3 PUFAs on intestinal microbiota and immune system. Mar. Drugs 2019, 17, 374. [Google Scholar] [CrossRef]
- Mendivil, C.O. Dietary fish, fish nutrients, and immune function: A review. Front. Nutr. 2021, 7, 617652. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Ianni, A.; Luca, A.D.; Martino, C.; Bennato, F.; Marone, E.; Grotta, L.; Cichelli, A.; Martino, G. Dietary supplementation of dried grape pomace increases the amount of linoleic acid in beef, reduces the lipid oxidation and modifies the volatile profile. Animals 2019, 9, 578. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Evolutionary aspects of diet: The omega-6/omega-3 ratio and the brain. Mol. Neurobiol. 2011, 44, 203–215. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An increase in the omega-6/omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S. The omega-6:omega-3 ratio: A critical appraisal and possible successor. Prostaglandins Leukot. Essent. Fatty Acids 2018, 132, 34–40. [Google Scholar] [CrossRef] [PubMed]
- FAO. Fats and fatty acids in human nutrition: Report of an expert consultation. FAO Food Nutr. Pap. 2010, 91, 1–166. Available online: https://www.fao.org/3/i1953e/I1953E.pdf (accessed on 14 March 2022).
- Horcada, A.; Polvillo, O.; Gonzalez-Redondo, P.; Lopez, A.; Tejerina, D.; Garcia-Torres, S. Stability of fatty acid composition of intramuscular fat from pasture- and grain-fed young bulls during the first 7 d postmortem. Arch. Anim. Breed. 2020, 63, 45–52. [Google Scholar] [CrossRef]
- De la Fuente, J.; Diaz, M.T.; Alvarez, I.; Oliver, M.A.; Font, I.F.M.; Sanudo, C.; Campo, M.M.; Montossi, F.; Nute, G.R.; Caneque, V. Fatty acid and vitamin E composition of intramuscular fat in cattle reared in different production systems. Meat Sci. 2009, 82, 331–337. [Google Scholar] [CrossRef]
- Untea, A.E.; Varzaru, I.; Vlaicu, P.A.; Turcu, R.P.; Panaite, T.D. Studies on antioxidant activities of grape pomace using in vitro, ex vivo, and in vivo models. J. Food Meas. Charact. 2022, 17, 121–128. [Google Scholar] [CrossRef]
- Carrillo, J.A.; He, Y.; Li, Y.; Liu, J.; Erdman, R.A.; Sonstegard, T.S.; Song, J. Integrated metabolomic and transcriptome analyses reveal finishing forage affects metabolic pathways related to beef quality and animal welfare. Sci. Rep. 2016, 6, 25948. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Zhou, Y.; Wang, M.; Guo, C.; Cao, Z.; Zhang, R.; Peng, C. A review of pharmacological and pharmacokinetic properties of stachydrine. Pharmacol. Res. 2020, 155, 104755. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, G.; Ghilardelli, F.; Mosconi, M.; Masoero, F.; Gallo, A. Occurrence of polyphenols, isoflavonoids, and their metabolites in milk samples from different cow feeding regimens. Dairy 2022, 3, 314–325. [Google Scholar] [CrossRef]
- Mahato, N.; Sharma, K.; Sinha, M.; Cho, M.H. Citrus waste derived nutra-/pharmaceuticals for health benefits: Current trends and future perspectives. J. Funct. Foods 2018, 40, 307–316. [Google Scholar] [CrossRef]
- Nii, Y.; Osawa, T.; Kunii, D.; Fukuta, K.; Sakai, K.; Kondo, M.; Yamamoto, S. Effect of citrus fruit (sudachi) juice on absorption of calcium from whole small fish in healthy young men. Food Sci. Technol. Res. 2006, 12, 27–30. [Google Scholar] [CrossRef]
- Supplee, G.C.; Bellis, B. Citric acid content of milk and milk products. J. Biol. Chem. 1921, 48, 453–461. [Google Scholar] [CrossRef]
- Gurrieri, F.; Audergon, J.M.; Albagnac, G.; Reich, M. Soluble sugars and carboxylic acids in ripe apricot fruit as parameters for distinguishing different cultivars. Euphytica 2001, 117, 183–189. [Google Scholar] [CrossRef]
- Gatmaitan, A.N.; Lin, J.Q.; Zhang, J.; Eberlin, L.S. Rapid analysis and authentication of meat using the MasSpec Pen technology. J. Agric. Food Chem. 2021, 69, 3527–3536. [Google Scholar] [CrossRef]
- Gomez, J.F.M.; Consolo, N.R.B.; Antonelo, D.S.; Beline, M.; Gagaoua, M.; Higuera-Padilla, A.; Colnago, L.A.; Gerrard, D.E.; Silva, S.L. Impact of cattle feeding strategy on the beef metabolome. Metabolites 2022, 12, 640. [Google Scholar] [CrossRef]
- Apaoblaza, A.; Gerrard, S.D.; Matarneh, S.K.; Wicks, J.C.; Kirkpatrick, L.; England, E.M.; Scheffler, T.L.; Duckett, S.K.; Shi, H.; Silva, S.L.; et al. Muscle from grass- and grain-fed cattle differs energetically. Meat Sci. 2020, 161, 107996. [Google Scholar] [CrossRef]
- Sabra, A.; Netticadan, T.; Wijekoon, C. Grape bioactive molecules, and the potential health benefits in reducing the risk of heart diseases. Food Chem. X 2021, 12, 100149. [Google Scholar] [CrossRef]
- Beslo, D.; Doslic, G.; Agic, D.; Rastija, V.; Speranda, M.; Gantner, V.; Lucic, B. Polyphenols in ruminant nutrition and their effects on reproduction. Antioxidants 2022, 11, 970. [Google Scholar] [CrossRef]
- Hilario, M.C.; Puga, C.D.; Ocana, A.N.; Romo, F.P. Antioxidant activity, bioactive polyphenols in Mexican goats’ milk cheeses on summer grazing. J. Dairy Res. 2010, 77, 20–26. [Google Scholar] [CrossRef]
- Monahan, F.J.; Schmidt, O.; Moloney, A.P. Meat provenance: Authentication of geographical origin and dietary background of meat. Meat Sci. 2018, 144, 2–14. [Google Scholar] [CrossRef]
- Prache, S.; Martin, B.; Coppa, M. Review: Authentication of grass-fed meat and dairy products from cattle and sheep. Animal 2020, 14, 854–863. [Google Scholar] [CrossRef]
- Marhuenda-Munoz, M.; Laveriano-Santos, E.P.; Tresserra-Rimbau, A.; Lamuela-Raventos, R.M.; Martinez-Huelamo, M.; Vallverdu-Queralt, A. Microbial phenolic metabolites: Which molecules actually have an effect on human health? Nutrients 2019, 11, 2725. [Google Scholar] [CrossRef]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.-J.G.; Fahey, G.C.; Merchen, N.R. Effects of ruminant digestion and metabolism on phenolic monomers of forages. Br. J. Nutr. 1983, 50, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Wein, S.; Beyer, B.; Zimmermann, B.F.; Blank, R.H.; Wolffram, S. Bioavailability of quercetin from onion extracts after intraruminal application in cows. J. Agric. Food Chem. 2018, 66, 10188–10192. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| GRASS 1 | GRAIN 2 | GRAPE 3 | p-Value | |
|---|---|---|---|---|
| C10:0 | 0.03 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.216 |
| C12:0 | 0.21 ± 0.02 a | 0.03 ± 0.00 b | 0.03 ± 0.01 b | <0.001 |
| C14:0 | 0.66 ± 0.05 a | 0.07 ± 0.00 b | 0.07 ± 0.00 b | <0.001 |
| C15:0 | 0.13 ± 0.01 a | 0.03 ± 0.00 b | 0.03 ± 0.00 b | <0.001 |
| C16:0 | 14.13 ± 0.22 | 13.82 ± 0.20 | 13.35 ± 0.17 | 0.156 |
| C16:1 c9 | 0.18 ± 0.02 a | 0.09 ± 0.01 b | 0.09 ± 0.01 b | <0.010 |
| C16:1 c7 | 1.06 ± 0.04 a | 0.14 ± 0.01 b | 0.12 ± 0.02 b | <0.001 |
| C17:0 | 0.20 ± 0.01 a | 0.06 ± 0.00 b | 0.07 ± 0.00 b | <0.001 |
| C18:0 | 1.73 ± 0.07 | 1.71 ± 0.05 | 1.67 ± 0.05 | 0.898 |
| C18:1 c9 | 3.17 ± 0.26 b | 22.27 ± 0.14 a | 22.15 ± 0.31 a | <0.001 |
| C18:1 c11 | 0.52 ± 0.05 | 0.52 ± 0.01 | 0.48 ± 0.01 | 0.896 |
| C18:2 n-6 (LA) 4 | 14.09 ± 0.42 b | 56.11 ± 0.57 a | 57.03 ± 0.64 a | <0.001 |
| C18:3 n-3 (ALA) 5 | 61.60 ± 0.93 a | 4.29 ± 0.30 b | 3.95 ± 0.66 b | <0.001 |
| C20:0 | 0.62 ± 0.03 a | 0.42 ± 0.02 b | 0.41 ± 0.04 b | <0.001 |
| C22:0 | 0.75 ± 0.04 a | 0.17 ± 0.01 b | 0.17 ± 0.01 b | <0.001 |
| C24:0 | 0.74 ± 0.03 a | 0.23 ± 0.01 b | 0.22 ± 0.01 b | <0.001 |
| ∑ SFA 6 | 19.36 ± 0.38 a | 16.57 ± 0.27 b | 16.03 ± 0.24 b | <0.001 |
| ∑ OCFA 7 | 0.33 ± 0.01 a | 0.09 ± 0.00 b | 0.10 ± 0.00 b | <0.001 |
| ∑ MUFA 8 | 4.94 ± 0.25 b | 23.02 ± 0.13 a | 22.98 ± 0.30 a | <0.001 |
| ∑ PUFA 9 | 75.71 ± 0.57 a | 60.41 ± 0.32 b | 60.99 ± 0.34 b | <0.001 |
| ∑ n-6 10 | 14.09 ± 0.42 b | 56.12 ± 0.57 a | 57.04 ± 0.63 a | <0.001 |
| ∑ n-3 11 | 61.62 ± 0.93 a | 4.29 ± 0.30 b | 3.95 ± 0.66 b | <0.001 |
| n-6:n-3 ratio 12 | 0.24 ± 0.01 b | 15.12 ± 1.28 a | 18.42 ± 2.34 a | <0.001 |
| DM 13 | 21.38 ± 0.63 b | 79.80 ± 0.97 a | 82.36 ± 1.32 a | <0.001 |
| Ash * | 6.60 ± 0.17 a | 4.02 ± 0.20 b | 3.79 ± 0.34 b | <0.001 |
| CP 14* | 13.40 ± 0.47 a | 9.65 ± 0.18 b | 9.29 ± 0.32 b | <0.001 |
| NDF 15* | 51.82 ± 0.92 a | 21.02 ± 0.39 b | 20.59 ± 0.62 b | <0.001 |
| ADF 16* | 33.38 ± 0.72 a | 10.09 ± 0.26 b | 9.94 ± 0.39 b | <0.001 |
| Energy 17 | 4465.49 ± 11.98 a | 4265.90 ± 15.98 b | 4274.76 ± 28.55 b | <0.001 |
| GRASS 1 | GRAIN 2 | GRAPE 3 | p-Value | |
|---|---|---|---|---|
| ∑ SFA 4 | 767.56 ± 188.89 | 996.16 ± 194.19 | 868.22 ± 266.53 | 0.700 |
| C12:0 | 1.04 ± 0.25 | 1.09 ± 0.26 | 0.63 ± 0.35 | 0.527 |
| C14:0 | 31.29 ± 10.15 | 53.81 ± 10.43 | 49.95 ± 14.32 | 0.287 |
| C15:0 | 8.16 ± 2.23 | 3.97 ± 2.29 | 3.34 ± 3.15 | 0.330 |
| C16:0 | 424.06 ± 107.99 | 681.64 ± 111.01 | 564.21 ± 152.37 | 0.261 |
| C17:0 | 26.17 ± 8.62 | 10.18 ± 8.86 | 23.16 ± 12.16 | 0.403 |
| C18:0 | 272.22 ± 66.47 | 242.23 ± 68.33 | 224.09 ± 93.79 | 0.907 |
| ∑ MUFA 5 | 748.55 ± 194.06 | 944.75 ± 188.04 | 1076.92 ± 258.41 | 0.572 |
| C14:1 9c | 5.57 ± 3.30 b | 19.10 ± 3.39 a | 13.67 ± 4.65 a,b | <0.050 |
| C16:1 9c | 60.47 ± 54.83 | 117.74 ± 56.37 | 240.76 ± 77.37 | 0.174 |
| C16:1 9t | 3.51 ± 0.33 a | 1.08 ± 0.20 b | 1.05 ± 0.27 b | <0.001 |
| C18:1 9c | 565.28 ± 148.34 | 733.76 ± 152.50 | 732.59 ± 209.30 | 0.694 |
| C18:1 9t | 2.24 ± 0.95 | 1.44 ± 0.27 | 1.75 ± 0.36 | 0.578 |
| C18:1 11t | 36.69 ± 11.11 a | 1.54 ± 2.02 b | 1.70 ± 2.70 b | <0.001 |
| ∑ PUFA 6 | 113.84 ± 12.60 | 111.65 ± 12.95 | 95.29 ± 17.78 | 0.669 |
| ∑ n-3 7 | 46.03 ± 4.79 a | 10.13 ± 4.92 b | 7.73 ± 6.76 b | <0.001 |
| C18:3 n-3 (ALA) 8 | 24.32 ± 3.70 a | 2.32 ± 3.81 b | 2.22 ± 5.23 b | <0.001 |
| C20:3 n-3 | 0.30 ± 0.07 a | 0.03 ± 0.07 b | 0.09 ± 0.09 a,b | <0.050 |
| C20:5 n-3 (EPA) 9 | 7.41 ± 0.38 a | 1.80 ± 0.39 b | 1.10 ± 0.53 b | <0.001 |
| C22:5 n-3 (DPA) 10 | 13.39 ± 0.96 a | 5.67 ± 0.99 b | 3.96 ± 1.36 b | <0.001 |
| C22:6 n-3 (DHA) 11 | 0.61 ± 0.08 a | 0.32 ± 0.08 b | 0.35 ± 0.11 a,b | <0.050 |
| ∑ n-6 12 | 67.07 ± 9.04 b | 100.32 ± 9.29 a | 86.61 ± 12.73 a,b | <0.050 |
| C18:2 n-6 (LA) 13 | 46.95 ± 7.07 | 65.61 ± 7.27 | 57.47 ± 9.97 | 0.195 |
| C18:3 n-6 | 0.29 ± 0.05 | 0.40 ± 0.05 | 0.42 ± 0.07 | 0.243 |
| C20:2 n-6 | 0.33 ± 0.09 | 0.57 ± 0.10 | 0.55 ± 0.13 | 0.186 |
| C20:3 n-6 | 3.34 ± 0.49 b | 5.87 ± 0.50 a | 4.61 ± 0.69 a,b | <0.010 |
| C20:4 n-6 | 14.61 ± 1.51 b | 22.53 ± 1.56 a | 19.21 ± 2.14 a,b | <0.010 |
| C22:4 n-6 | 1.56 ± 0.49 b | 5.34 ± 0.51 a | 4.36 ± 0.70 a | <0.001 |
| n-6:n-3 ratio 14 | 1.65 ± 0.44 b | 8.39 ± 0.45 a | 9.82 ± 0.62 a | <0.001 |
| ∑ CLA 15 | 5.14 ± 1.20 | 2.28 ± 1.23 | 2.86 ± 1.69 | 0.243 |
| C18:2 9c,11t/9c,7t | 4.01 ± 1.05 | 1.63 ± 1.08 | 1.96 ± 1.48 | 0.269 |
| ∑ AD 16 | 18.07 ± 5.07 | 9.29 ± 5.21 | 17.01 ± 7.16 | 0.438 |
| ∑ CLnA 17 | 0.36 ± 0.05 | 0.25 ± 0.05 | 0.25 ± 0.07 | 0.292 |
| C18:3 9c,11t,15t | 0.12 ± 0.03 a | 0.02 ± 0.03 b | 0.05 ± 0.04 a,b | <0.050 |
| C18:3 9c,11t,15c | 0.24 ± 0.03 | 0.23 ± 0.03 | 0.20 ± 0.04 | 0.758 |
| ∑ FA 18 | 1686.08 ± 400.01 | 2080.23 ± 399.93 | 2090.96 ± 549.44 | 0.495 |
| GRASS 1 | GRAIN 2 | GRAPE 3 | Class Error | OOB 4 | |
|---|---|---|---|---|---|
| Phytochemicals | |||||
| GRASS | 17 | 1 | 0 | 5.60% | 17.30% |
| GRAIN | 0 | 10 | 6 | 37.50% | |
| GRAPE | 0 | 2 | 16 | 11.10% | |
| Phytochemicals and fatty acids | |||||
| GRASS | 17 | 1 | 0 | 5.60% | 15.40% |
| GRAIN | 0 | 13 | 3 | 18.80% | |
| GRAPE | 1 | 3 | 14 | 22.00% | |
| GRASS 1 | GRAIN 2 | GRAPE 3 | p-Value | |
|---|---|---|---|---|
| Stachydrine | 0.65 ± 0.04 a | 0.26 ± 0.05 b | 0.28 ± 0.06 b | <0.001 |
| 4-Ethylphenol | 3.92 ± 0.44 | 3.00 ± 0.45 | 3.83 ± 0.60 | 0.287 |
| Hippuric acid | 13.37 ± 1.20 a | 7.77 ± 1.24 b | 5.52 ± 1.66 b | <0.001 |
| Citric acid | 379.20 ± 35.20 a | 79.10 ± 36.30 b | 51.40 ± 48.70 b | <0.001 |
| Succinic acid | 36.80 ± 2.66 a | 17.80 ± 2.74 b | 13.70 ± 3.67 b | <0.001 |
| Fumaric acid | 2.68 ± 0.34 a | 0.90 ± 0.35 b | 0.62 ± 0.47 b | <0.001 |
| Caffeic acid | 0.04 ± 0.02 | 0.04 ± 0.02 | 0.01 ± 0.03 | 0.570 |
| p-Coumaric acid | 1.56 ± 0.25 b | 2.47 ± 0.25 a | 2.53 ± 0.34 a | <0.050 |
| 4-Hydroxybenzoic acid | 0.38 ± 0.22 | 0.02 ± 0.23 | 0.85 ± 0.30 | 0.058 |
| Gallic acid | 0.01 ± 0.02 | 0.00 ± 0.02 | 0.05 ± 0.03 | 0.370 |
| Ethyl gallate | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.02 ± 0.01 | 0.871 |
| Vanillic acid | 0.18 ± 0.16 | 0.28 ± 0.16 | 0.38 ± 0.22 | 0.733 |
| D-Tartaric acid | 2.29 ± 0.52 | 2.34 ± 0.53 | 1.71 ± 0.71 | 0.734 |
| Pyrocatechol sulfate | 0.52 ± 0.09 | 0.55 ± 0.10 | 0.70 ± 0.13 | 0.501 |
| Coixol | 1.07 ± 0.36 | 1.16 ± 0.37 | 1.73 ± 0.49 | 0.510 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krusinski, L.; Maciel, I.C.F.; van Vliet, S.; Ahsin, M.; Lu, G.; Rowntree, J.E.; Fenton, J.I. Measuring the Phytochemical Richness of Meat: Effects of Grass/Grain Finishing Systems and Grapeseed Extract Supplementation on the Fatty Acid and Phytochemical Content of Beef. Foods 2023, 12, 3547. https://doi.org/10.3390/foods12193547
Krusinski L, Maciel ICF, van Vliet S, Ahsin M, Lu G, Rowntree JE, Fenton JI. Measuring the Phytochemical Richness of Meat: Effects of Grass/Grain Finishing Systems and Grapeseed Extract Supplementation on the Fatty Acid and Phytochemical Content of Beef. Foods. 2023; 12(19):3547. https://doi.org/10.3390/foods12193547
Chicago/Turabian StyleKrusinski, Lucas, Isabella C. F. Maciel, Stephan van Vliet, Muhammad Ahsin, Guanqi Lu, Jason E. Rowntree, and Jenifer I. Fenton. 2023. "Measuring the Phytochemical Richness of Meat: Effects of Grass/Grain Finishing Systems and Grapeseed Extract Supplementation on the Fatty Acid and Phytochemical Content of Beef" Foods 12, no. 19: 3547. https://doi.org/10.3390/foods12193547