
Selection of Salt-Tolerance and Ester-Producing Mutant Saccharomyces cerevisiae to Improve Flavour Formation of Soy Sauce during Co-Fermentation with Torulopsis globosa
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, and Medium
2.2. ARTP Mutagenesis and High-Salt-Temperature Based Screening
2.3. VHG Fermentation
2.4. Stress Tolerance Test
2.5. Growth Curves Test
2.6. Construction of ATF1 Gene Overexpression Strain
2.7. RT-qPCR Assay
2.8. Soy Sauce Fermentation Experiment
3. Results and Discussion
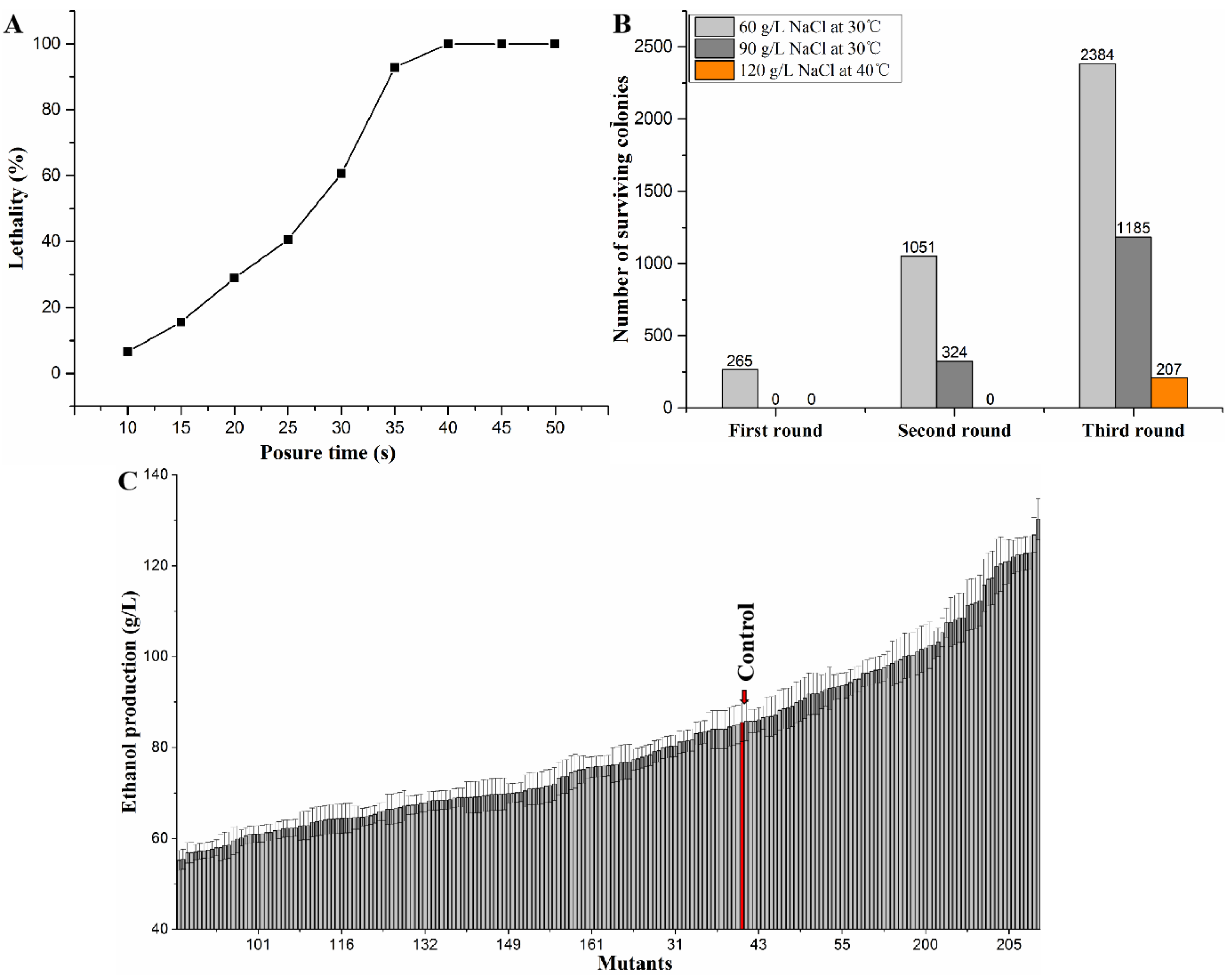
3.1. ARTP Mutagenesis and Mutant Screening
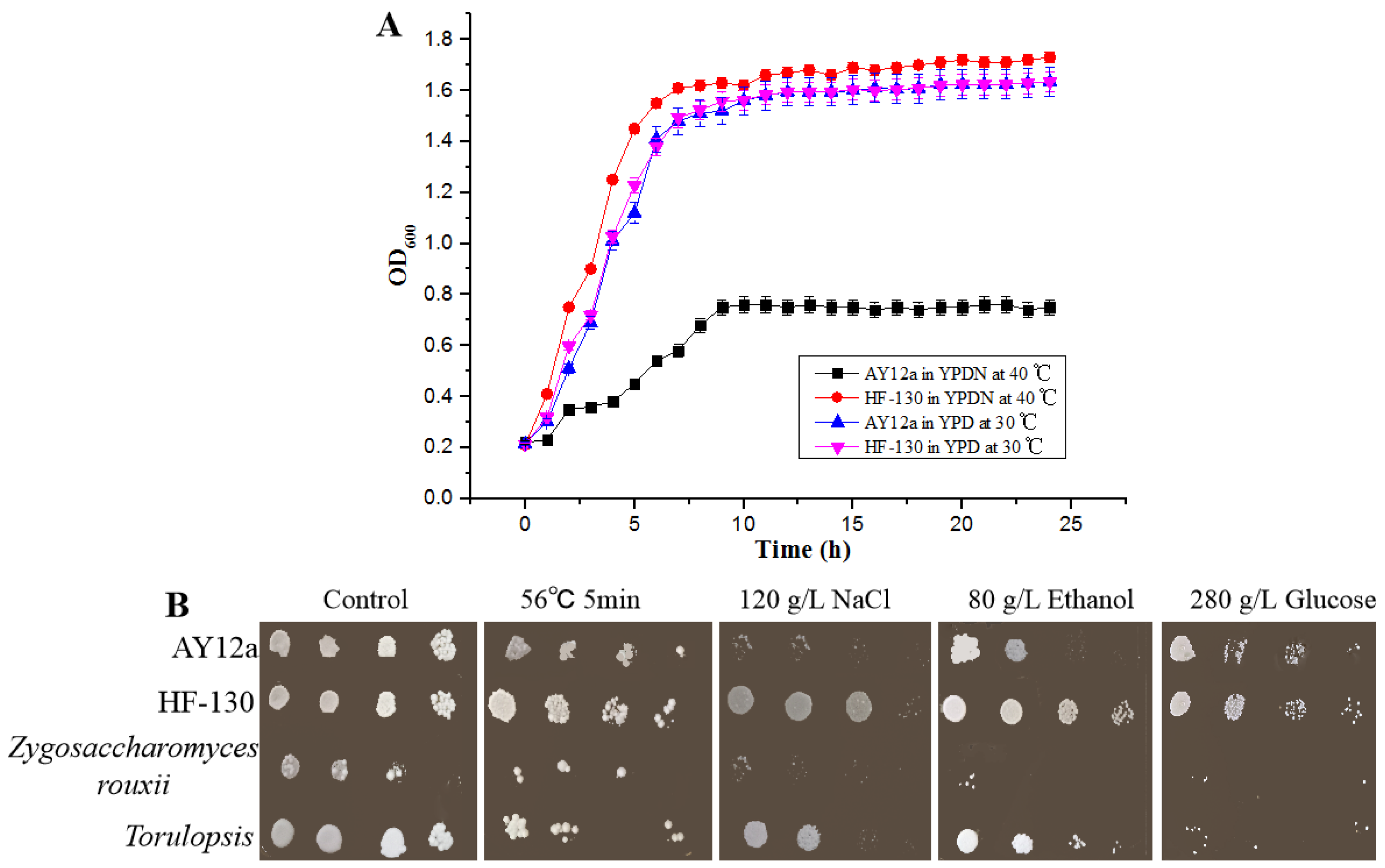
3.2. Growth, Tolerance, and Fermentation Performance of HF-130
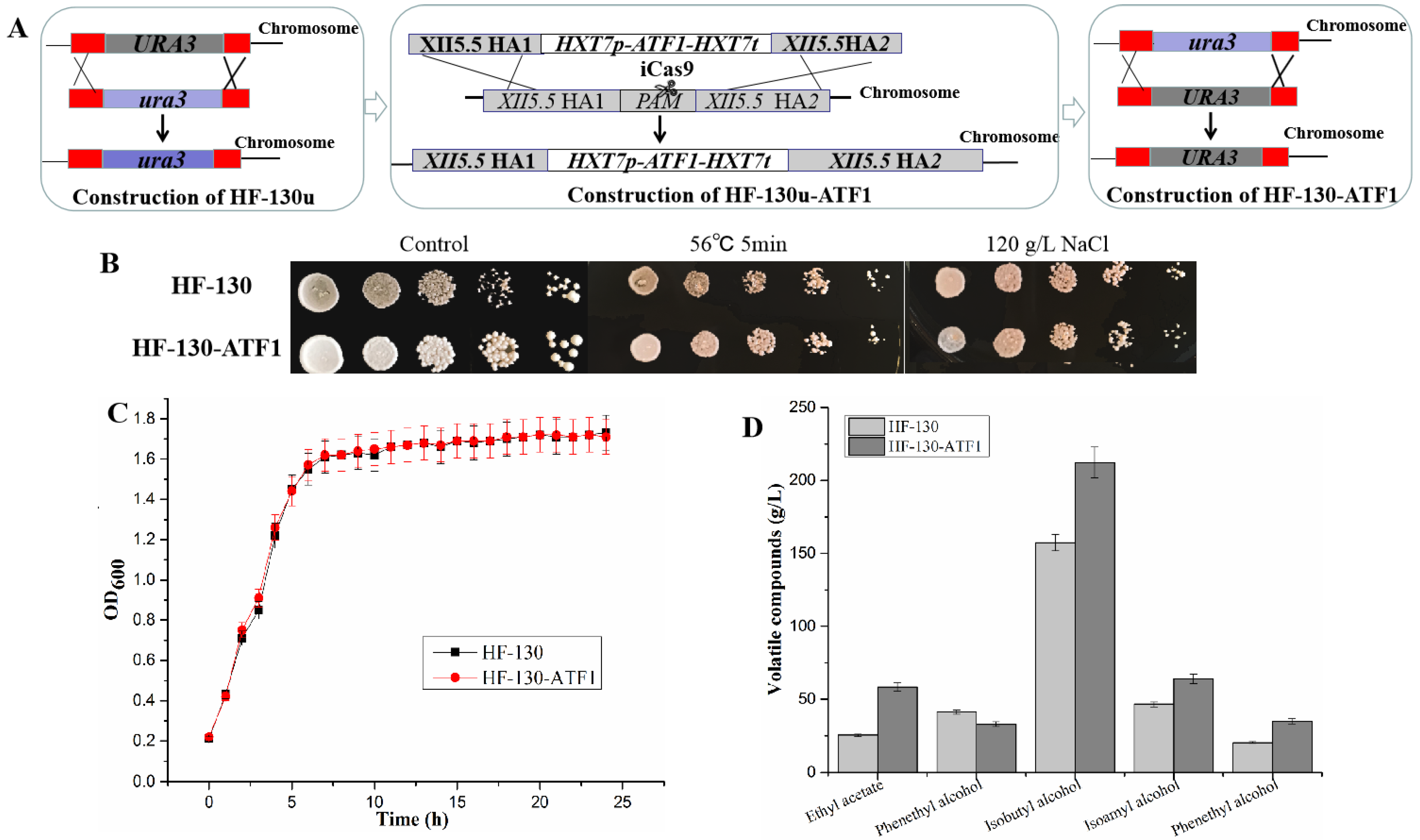
3.3. Overexpressing of ATF1 Gene in HF-130
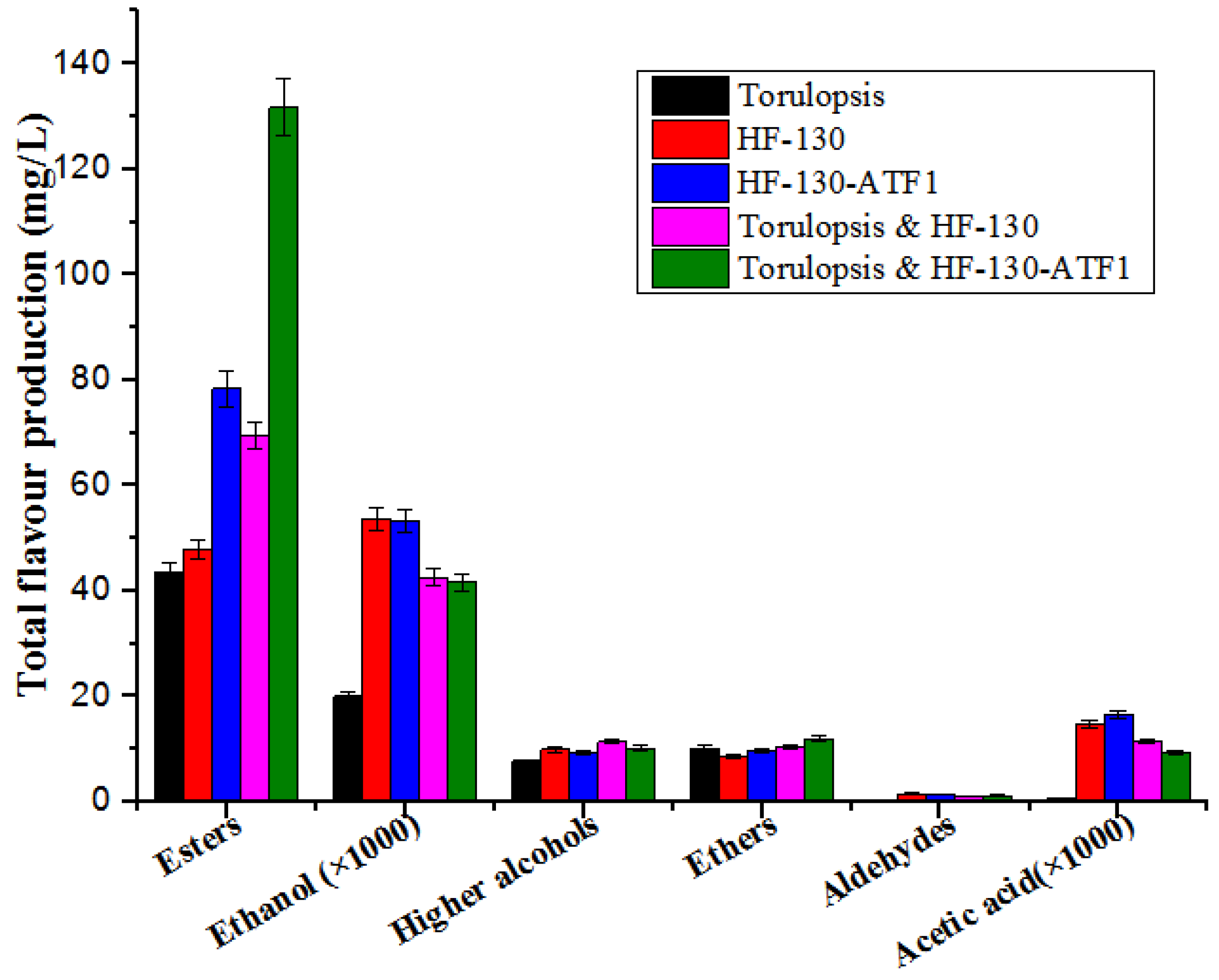
3.4. Co-Fermentation of Torulopsis and HF-130-ATF1 in Soy Sauce Fermentation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gao, X.L.; Cui, C.; Zhao, H.F.; Zhao, M.M.; Yang, L.; Ren, J.Y. Changes in volatile aroma compounds of traditional Chinese-type soy sauce during moromi fermentation and heat treatment. Food Sci. Biotechnol. 2010, 19, 889–898. [Google Scholar] [CrossRef]
- Zhang, Y.; Tao, W. Flavor and taste compounds analysis in Chinese solid fermented soy sauce. Afr. J. Biotechnol. 2009, 8, 673–681. [Google Scholar]
- Wicklow, D.T.; McAlpin, C.E.; Yeoh, Q.L. Diversity of Aspergillus oryzae genotypes (RFLP) isolated from traditional soy sauce production within Malaysia and Southeast Asia. Mycoscience 2007, 48, 373–380. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Taljaard, L.J. Industrialization of Mageu Fermentation in South Africa. In Industrialization of Indigenous Fermented Foods, Revised and Expanded; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Nunomura, N.; Sasaki, M.; Asao, Y.; Yokotsuka, T. Identification of Volatile Components in Shoyu (Soy Sauce) by Gas Chromatography-Mass Spectrometry. Agric. Biol. Chem. 1976, 40, 485–490. [Google Scholar]
- Franssen, O.; Hennink, W.E. A novel preparation method for polymeric microparticles without the use of organic solvents. Int. J. Pharm. 1998, 168, 1–7. [Google Scholar] [CrossRef]
- Feng, Z.C.; Zhang, Q.Q.; Kan, Q.H.; Wang, L.F. Determination and analysis on flavor components of weijiatian soy sauce. J. Anhui Univ. Technol. Sci. 2010, 25, 8–12. [Google Scholar]
- Singracha, P.; Niamsiri, N.; Visessanguan, W.; Lertsiri, S.; Assavanig, A. Application of lactic acid bacteria and yeasts as starter cultures for reduced-salt soy sauce (moromi) fermentation. LWT Food Sci. Technol. 2016, 78, 181–188. [Google Scholar] [CrossRef]
- Harada, R.; Yuzuki, M.; Ito, K.; Shiga, K.; Bamba, T.; Fukusaki, E. Microbe participation in aroma production during soy sauce fermentation. J. Biosci. Bioeng. 2018, 125, 688–694. [Google Scholar] [CrossRef]
- Feng, Y.; Cai, Y.; Su, G.; Zhao, H.; Wang, C.; Zhao, M. Evaluation of aroma differences between high-salt liquid-state fermentation and low-salt solid-state fermentation soy sauces from China. Food Chem. 2014, 145, 126–134. [Google Scholar] [CrossRef]
- Liu, M.-C.; Li, Z.-G.; Deng, W.; Wang, G.-M.; Yang, Y.-W. Changes in volatile compounds of pickled mustard tuber (Brassica juncea var. tsatsai) during the pickling process. Int. J. Food Sci. Technol. 2009, 44, 2278–2286. [Google Scholar] [CrossRef]
- Coutron-Gambotti, C.; Gandemer, G. Lipolysis and oxidation in subcutaneous adipose tissue during dry-cured ham processing. Food Chem. 1999, 64, 95–101. [Google Scholar] [CrossRef]
- Murooka, Y.; Yamshita, M. Traditional healthful fermented products of Japan. J. Ind. Microbiol. Biotechnol. 2008, 35, 791–798. [Google Scholar] [CrossRef]
- Makino, Y.; Takegami, I.; Matsushita, S.; Matsuo, Y.; Shirakawa, T. Production of soy-sauce by use of yeast cultured in a medium including wastewater produced by desalted soy-Sauce lees. Nippon. Shokuhin Kagaku Kogaku Kaishi 1999, 46, 664–668. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, E.J.; Shin, O.S.; Ji, W.D.; Choi, M.R.; Kim, J.K. Volatile components in the soy sauce manufactured by Bacillus species and fused Yeast. J. Microbiol. Biotechnol. 1996, 6, 194–301. [Google Scholar]
- Yan, Y.Z.; Qian, Y.L.; Ji, F.D.; Chen, J.Y.; Han, B.Z. Microbial composition during Chinese soy sauce koji-making based on culture dependent and independent methods. Food Microbiol. 2013, 34, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Suezawa, Y.; Kimura, I.; Inoue, M.; Gohda, N.; Suzuki, M. Identification and Typing of Miso and Soy Sauce Fermentation Yeasts, Candida etchellsii and C. versatilis, Based on sequence analyses of the D1D2 domain of the 26S ribosomal RNA gene, and the region of internal transcribed spacer 1, 5.8S ribosomal RNA gene. J. Agric. Chem. Soc. Japan 2006, 70, 348–354. [Google Scholar]
- Feng, J.; Zhan, X.B.; Wang, D.; Zhang, L.M.; Lin, C.C. Identification and analysis of the metabolic functions of a high-salt-tolerant halophilic aromatic yeast Candida etchellsiifor soy sauce production. World J. Microbiol. Biotechnol. 2012, 28, 1451–1458. [Google Scholar] [CrossRef] [PubMed]
- Ohata, M.; Kohama, K.; Morimitsu, Y.; Kubota, K.; Sugawara, E. The formation mechanism by yeast of 4-hydroxy-2(or 5)-ethyl-5(or 2)-methyl-3(2H)-furanone in Miso. Biosci. Biotechnol. Biochem. 2007, 71, 407–413. [Google Scholar] [CrossRef]
- Cao, X.; Song, Q.; Wang, C.; Hou, L. Genome shuffling of Hansenula anomala to improve flavour formation of soy sauce. World J. Microbiol. Biotechnol. 2012, 28, 1857–1862. [Google Scholar] [CrossRef]
- Cao, X.; Hou, L.; Lu, M.; Wang, C. Improvement of soy-sauce flavour by genome shuffling in Candida versatilis to improve salt stress resistance. Int. J. Food Sci. Technol. 2009, 45, 17–22. [Google Scholar] [CrossRef]
- Gao, X.-L.; Zhao, X.; Hu, F.; Zhang, Z.-K.; Fu, J.-Y.; Liu, Z.; Wang, B.; He, R.-H.; Ma, H.-L.; Ho, C.-T. The latest advances on soy sauce research in the past decade: Emphasis on the advances in China. Food Res. Int. 2023; accepted. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Van Laere, S.D.M.; Vanderhaegen, B.M.P.; Derdelinckx, G.; Dufour, J.-P.; Pretorius, I.S.; Winderickx, J.; Thevelein, J.M.; Delvaux, F.R. Expression levels of the yeast alcohol acetyltransferase genes ATF1, Lg-ATF1, and ATF2 control the formation of a broad range of volatile esters. Appl. Environ. Microbiol. 2003, 69, 5228–5237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.W.; Zhang, C.Y.; Dai, L.H.; Liu JD, Y.-L.; Guo, X.W.; Xiao, D.G. Effects of overexpression of the alcohol acetyltransferase- encoding gene ATF1 and disruption of the esterase-encoding gene IAH1 on the flavour profiles of Chinese yellow rice wine. Int. J. Food Sci. Technol. 2012, 47, 2590–2596. [Google Scholar] [CrossRef]
- Ku, K.L.; Chen, T.P.; Chiou, Y.Y. Apparatus used for small-scale volatile extraction from ethanol-supplemented low-salt miso and GC-MS characterization of the extracted flavors. J. Agric. Food Chem. 2000, 48, 3507. [Google Scholar] [CrossRef]
- Dong, J.; Hong, K.-Q.; Hao, A.-L.; Zhang, C.-Y.; Fu, X.-M.; Wang, P.-F.; Xiao, D.-G. Gradual enhancement of ethyl acetate production through promoter engineering in Chinese Liquor yeast strains. Biotechnol. Prog. 2018, 34, 328–336. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Wang, Z.; Zhang, Y.; Shi, S.; Nielsen, J.; Liu, Z. A gRNA-tRNA array for CRISPR-Cas9 based rapid multiplexed genome editing in Saccharomyces cerevisiae. Nat. Commun. 2019, 10, 1053. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.-L.; Liu, E.-M.; Yin, Y.-Y.; Yang, L.-X.; Huang, Q.-R.; Chen, S.; Ho, C.-T. Enhancing activities of salt-tolerant proteases secreted by Aspergillus oryzae using atmospheric and room temperature plasma mutagenesis. J. Agric. Food Chem. 2020, 68, 2757–2764. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.-Q.; Hou, X.-Y.; Hao, A.-L.; Wang, P.-F.; Fu, X.-M.; Lv, A.; Dong, J. Truncation of CYR1 promoter in industrial ethanol yeasts for improved ethanol yield in high temperature condition. Process Biochem. 2018, 65, 37–45. [Google Scholar] [CrossRef]
- Stanhill, A.; Schick, N.; Engelberg, D. The Yeast Ras/Cyclic AMP pathway induces invasive growth by suppressing the cellular stress response. Mol. Cell. Biol. 1999, 19, 7529–7538. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H.; Willems, A.R.; Woods, R.A. Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast 2010, 11, 355–360. [Google Scholar] [CrossRef]
- Sun, S.Y.; Jiang, W.G.; Zhao, Y.P. Profile of volatile compounds in 12 Chinese soy sauces produced by a high-salt-diluted state fermentation. J. Inst. Brew. 2010, 116, 316–328. [Google Scholar] [CrossRef]
- Shan, P.; Ho, C.-T.; Zhang, L.; Gao, X.; Lin, H.; Xu, T.; Wang, B.; Fu, J.; He, R.; Zhang, Y. Degradation mechanism of soybean protein B3 subunit catalyzed by prolyl endopeptidase from Aspergillus niger during Soy Sauce Fermentation. J. Agric. Food Chem. 2022, 70, 5869–5878. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, X.; Duan, X.; Kang, B.; Liu, J.; Fu, C.; Wang, C.; Li, D.; Xu, N. Effect of fermentation conditions on the formation of ammonium salt in soy sauce. LWT Food Sci. Technol. 2022, 153, 112492. [Google Scholar] [CrossRef]
- Devanthi, P.V.P.; Gkatzionis, K. Soy sauce fermentation: Microorganisms, aroma formation, and process modification. Food Res. Int. 2019, 120, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Dakal, T.C.; Solieri, L.; Giudici, P. Adaptive response and tolerance to sugar and salt stress in the food yeast Zygosaccharomyces rouxii. Int. J. Food Microbiol. 2014, 185, 140–157. [Google Scholar] [CrossRef]
- Jansen, M.; Veurink, J.H.; Euverink, G.J.W.; Dijkhuizen, L. Growth of the salt-tolerant yeast Zygosaccharomyces rouxii in microtiter plates: Effects of NaCl, pH and temperature on growth and fusel alcohol production from branched-chain amino acids. FEMS Yeast Res. 2003, 3, 313–318. [Google Scholar] [PubMed]
- Hauck, T.; Landmann, C.; Brühlmann, F.; Schwab, W. Formation of 5-methyl-4-hydroxy-3[2H]-furanone in cytosolic extracts obtained from Zygosaccharomyces rouxii. J. Agric. Food Chem. 2003, 51, 1410–1414. [Google Scholar] [CrossRef]
- Sluis CV, D.; Stoffelen CJ, P.; Castelein, S.J.; Engbers GH, M.; Schure EG, T.; Tramper, J.; Wijffels, R.H. Immobilized salt-tolerant yeasts: Application of a new polyethylene-oxide support in a continuous stirred-tank reactor for flavour production. J. Biotechnol. 2001, 88, 129–139. [Google Scholar] [CrossRef]
- Yan, M. Breeding of High Salt-Tolerant Soy Sauce Yeast and Its Application in Soy Sauce Brewing. Master’s Thesis, Hubei University of Technology, Wuhan, China, June 2014. [Google Scholar]
- Fujii, T.; Yoshimoto, H.; Nagasawa, N.; Bogaki, T.; Tamai, Y.; Hamachi, M. Nucleotide sequences of alcohol acetyltransferase genes from lager brewing yeast, Saccharomyces carlsbergensis. Yeast 1996, 12, 593–598. [Google Scholar] [CrossRef]
- Fujii, T.; Nagasawa, N.; Iwamatsu, A.; Bogaki, T.; Hamachi, M. Molecular cloning, sequence analysis, and expression of the yeast alcohol acetyltransferase gene. Appl. Environ. Microbiol. 1994, 60, 2786–2792. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source or Reference |
|---|---|---|
| AY12a | MATa | Angel Yeast Co., Ltd. (Wuhan, China) |
| HF-130 | MATa, mutant obtained via ARTP Mutagenesis | This study |
| Zygsoaccharomyces rouxii | MAT | This lab |
| Torulopsis globosa | MAT | This lab |
| AY12a-u | MATa ura3 | [26] |
| HF-130u | MATa ura3 | This study |
| HF-130-ATF1 | MATa, XII5.5::PGK1p-ATF1-PGK1t | This study |
| Plasmids | ||
| pCas9 | 2μ ampR TEF1p-iCas9 SNR52p | [27] |
| pCas9-XII5.5 | pCas9, containing 20-bp of XII5.5 gRNA sequence for gene insertion | This study |
| Primers | Sequence (5′-3′) |
|---|---|
| XII5.5-F | CCGAAGTTGATTTGCTTCAAACC |
| XII5.5-R | CCCGAAATTGTTCCTACGAGAGCACATTTCGCCCGTTCC |
| HXT7P-F | GAGCGGGAACGGGCGAAATGTGCTCTCGTAGGAACAATTTCGGGCC |
| HXT7P-R | GATTTTTCTCATCGATTTCATTCATGGATCCACTAGTTCTAGATTTTTGATTAAAATTAAAAAAAC |
| ATF1-UF | GTTTTTTTAATTTTAATCAAAAATCTAGAACTAGTGGATCCATGAATGAAATCGATGAGAAAAATC |
| ATF1-DR | TATCGAATTCCTGCAGCCCGGGCTAAGGGCCTAAAAGGAGAGCTTTG |
| HXT7T-F | GCTCTCCTTTTAGGCCCTTAGCCCGGGCTGCAGGAATTCGATATC |
| HXT7T-R | GTAGCCAAGACCAAATAATACGTCGACGCAAGAACCATAATCCTC |
| XII5.5-U | GGAGGATTATGGTTCTTGCGTCGACGTATTATTTGGTCTTGGCTACTGGC |
| XII5.5-D | CTTTCTAGACTGATAGTTGGAGGCAC |
| URA3-F | GACCATCAAAGAAGGTTAATGTGGCTGTGGTTTCAGGG |
| URA3-R | CTTGATTTGTGCCCCGTAAAATACTGTTACTTGG |
| RTATF1-F | GGGTTCAATATACAAGGCTTCG |
| RTATF1-R | GCATCGGGCTCCTCTAACTG |
| ACT1-F | TTATTGATAACGGTTCTGGTATG |
| ACT1-R | CCTTGGTGTCTTGGTCTAC |
| Parameter | Fermentation at 30 °C without NaCl | Fermentation at 40 °C with 120 g/L NaCl | ||
|---|---|---|---|---|
| AY12-G | HF-130 | AY12-G | HF-130 | |
| Ethanol production/(g/L) | 116.70 ± 0.72 | 118.40 ± 0.91 | 65.53 ± 0.64 | 130.26 ± 0.79 |
| Residual glucose/(g/L) | 24.3 ± 1.0 | 22.5 ± 1.2 | 43.0 ± 0.8 | 20.7 ± 0.8 |
| Glycerol/(g/L) | 9.91 ± 0.36 | 9.94 ± 0.49 | 7.61 ± 0.37 | 10.13 ± 0.42 |
| Organic acid b/(g/L) | 3.45 ± 0.25 | 3.56 ± 0.28 | 2.58 ± 0.20 | 3.72 ± 0.24 |
| Loss of CO2/(g/L) | 121.46 ± 0.68 | 122.79 ± 0.71 | 77.68 ± 0.59 | 128.53 ± 0.63 |
| Biomass/(g/L) | 7.89 ± 0.42 | 7.95 ± 0.40 | 5.28 ± 0.29 | 8.01± 0.32 |
| Total glucose/(g/L) | 279.42 ± 1.40 | 279.40 ± 1.56 | 279.11 ± 1.23 | 279.56 ± 1.47 |
| Cell surviving rate in 48 h/(%) | Not test | Not test | 41.1 ± 2.7 | 86.3 ± 1.2 |
| Fermentation time/(h) | 84 | 84 | 106 | 72 |
| Productivity/(g/L/h) | 1.389 ± 0.009 | 1.410 ± 0.011 | 0.679 ± 0.010 | 1.810 ± 0.009 |
| Parameter d | Torulopsis | HF-130 e | HF-130-ATF1 e | Torulopsis and HF-130 e | Torulopsis and HF-130-ATF1 e |
|---|---|---|---|---|---|
| Residual glucose/(g/L) | 19.62 ± 0.84 | 10.36 ± 0.51 | 10.26 ± 0.43 | 6.64 ± 0.21 * | 6.53 ± 0.32 * |
| Ethanol production/(g/L) | 20.11 ± 1.12 | 53.56 ± 2.15 ** | 53.25 ± 1.58 ** | 42.65 ± 1.35 ** | 41.62 ± 2.01 ** |
| Ethyl acetate/(mg/L) | 36.93 ± 0.65 | 24.65 ± 0.23 | 53.26 ± 0.41 ** | 42.78 ± 0.71 ** | 101.56 ± 1.01 ** |
| Isoamyl acetate/(mg/L) | 1.45 ± 0.08 | 1.56 ± 0.10 | 2.15 ± 0.11 | 2.57 ± 0.12 * | 4.87 ± 0.19 * |
| Ethyl propionate/(mg/L) | 0.85 ± 0.01 | 0.64 ± 0.01 | 0.81 ± 0.01 | 0.84 ± 0.01 | 1.02 ± 0.01 |
| Ethyl caprylate/(mg/L) | 0.25 ± 0.02 | 2.12 ± 0.14 ** | 2.25 ± 0.16 ** | 2.32 ± 0.21 ** | 2.69 ± 0.22 ** |
| Phenylethyl acetate/(mg/L) | 3.41 ± 0.16 | 16.54 ± 0.56 ** | 17.15 ± 0.63 ** | 17.91 ± 0.73 ** | 17.41 ± 0.59 ** |
| Ethyl caprate/(mg/L) | 0.37 ± 0.05 | 1.25 ± 0.09 ** | 1.15 ± 0.08 ** | 1.76 ± 0.10 ** | 2.04 ± 0.15 ** |
| Isobutyl acetate/(mg/L) | 0.39 ± 0.05 | 0.33 ± 0.05 | 0.45 ± 0.01 | 0.34 ± 0.02 | 0.59 ± 0.05 * |
| Dimethyl ether/(mg/L) | 10.23 ± 0.25 | 8.56 ± 0.63 | 9.66 ± 0.69 | 10.44 ± 0.11 | 11.96 ± 0.84 |
| Trimethylsilyl decyl ester/(mg/L) | /c | 0.80 ± 0.08 | 1.11 ± 0.12 | 1.07 ± 0.11 | 1.58 ± 0.09 |
| Acetic acid/(g/L) | 0.56 ± 0.09 | 14.74 ± 0.15 ** | 16.55 ± 0.21 ** | 11.56 ± 0.22 ** | 9.26 ± 0.16 ** |
| Isoamyl alcohol/(mg/L) | 1.84 ± 0.12 | 1.56 ± 0.12 | 2.61 ± 0.10 | 2.27 ± 0.15 * | 2.52 ± 0.13 * |
| phenylethyl alcohol/(mg/L) | 5.88 ± 0.51 | 6.89 ± 0.57 | 4.55 ± 0.85 | 7.6 ± 0.59 * | 6.07 ± 0.85 * |
| Phenylacetaldehyde/(mg/L) | 0.19 ± 0.08 | 1.56 ± 0.02 ** | 1.36 ± 0.08 ** | 1.05 ± 0.04 ** | 1.25 ± 0.01 ** |
| 2-Methylbutanol/(mg/L) | /c | 1.45 ± 0.05 | 2.14 ± 0.09 | 1.53 ± 0.05 | 1.59 ± 0.07 |
| 1-Octadecene/(mg/L) | 0.05 ± 0.01 | 0.06 ± 0.01 | /c | 0.05 ± 0.01 | 0.05 ± 0.01 |
| Octanoic acid/(mg/L) | /c | 2.01 ± 0.08 | 2.11 ± 0.05 | 1.92 ± 0.01 | 1.82 ± 0.04 |
| Fermentation time/(day) | 15 | 13 | 13 | 10 | 10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, K.-Q.; Fu, X.-M.; Lei, F.-F.; Chen, D.; He, D.-P. Selection of Salt-Tolerance and Ester-Producing Mutant Saccharomyces cerevisiae to Improve Flavour Formation of Soy Sauce during Co-Fermentation with Torulopsis globosa. Foods 2023, 12, 3449. https://doi.org/10.3390/foods12183449
Hong K-Q, Fu X-M, Lei F-F, Chen D, He D-P. Selection of Salt-Tolerance and Ester-Producing Mutant Saccharomyces cerevisiae to Improve Flavour Formation of Soy Sauce during Co-Fermentation with Torulopsis globosa. Foods. 2023; 12(18):3449. https://doi.org/10.3390/foods12183449
Chicago/Turabian StyleHong, Kun-Qiang, Xiao-Meng Fu, Fen-Fen Lei, Dong Chen, and Dong-Ping He. 2023. "Selection of Salt-Tolerance and Ester-Producing Mutant Saccharomyces cerevisiae to Improve Flavour Formation of Soy Sauce during Co-Fermentation with Torulopsis globosa" Foods 12, no. 18: 3449. https://doi.org/10.3390/foods12183449