
Difficulties in Establishing the Adverse Effects of β-Casomorphin-7 Released from β-Casein Variants—A Review
 , ,
, ,
Abstract
:1. Introduction
2. β-Casein Polymorphism
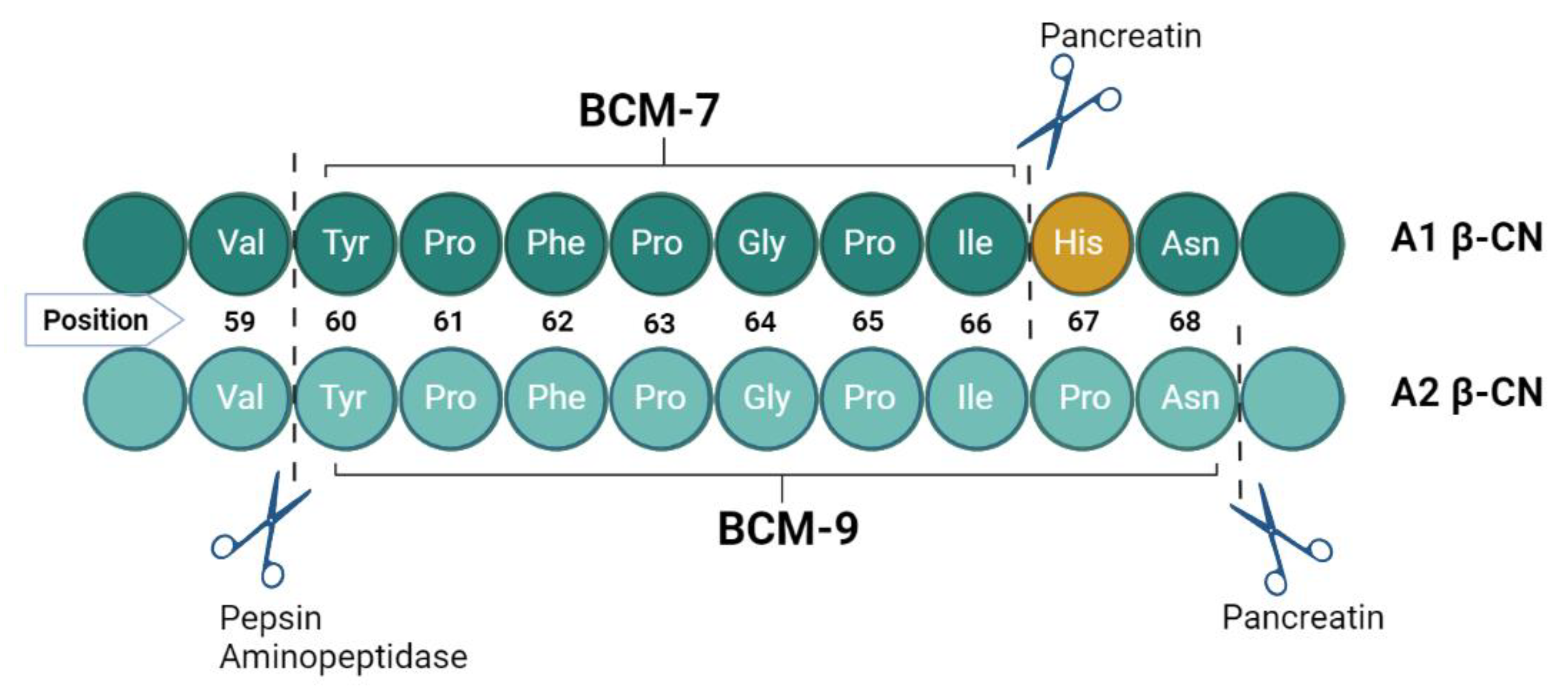
3. Digestion of β-CN and Generation of BCM-7
4. BCM-7 Threshold on Physiological Effects
5. Evidence from Animal and Human Studies on the Potential Adverse Effects of BCM-7 on the Gastrointestinal, Immunological, and Neurological Tract
Animal Studies
6. Human Studies

{kind=link}
{kind=link}
{kind=link}
| MODEL | Design | Methods | Biomarkers Evaluated | Sample | Significant Results/ Outcomes | Study | ||
|---|---|---|---|---|---|---|---|---|
| Children 3–10 y (n = 137) | 51 Neurotypical group 86 ASD group | No dietary intervention | DPP-IV and BCM-7 | Blood and urine | Level ng/mL | ASD | Control | Jarmołowska et al., 2019 [43] |
| BCM-7 blood | 42.9 ± 2.5 | 26.4 ± 1.6 | ||||||
| DPP-IV blood | 1089 ± 44 | 934 ± 52 | ||||||
| No significant difference was detected in BCM7 urinary levels | ||||||||
| Children 4–8 y (n = 20) | 10 Neurotypical group 10 ASD group | No dietary intervention | BCM-7 | Urine | Level pg/mL | ASD | Control | Sokolov et al., 2014 [65] |
| BCM-7 urine | 75 ± 10 | 58 ± 7 | ||||||
| Children and young 3–19 y (n = 404) | 296 Neurotypical group 88 ASD group | No dietary intervention | μ-opioid receptor and DPP-IV gene expression | Blood | Correlation in DPP-IV gene expression under the influence of BCM7 and hydrolyzed milk among healthy children and ASD with genotype GG. | Cieślińska et al., 2015 [66] | ||
| Chinese lactose-intolerant children 5–6 y (n = 75) | A1A2 milk group A2A2 milk group | 150 mL twice daily, 5 days of intervention; Double-blind, randomized and crossover | Gastrointestinal discomforts, Cognitive responses, BCM-7, MPO, IgE, IgG, IgG-1, IL-4 and GSH | Blood and feces |  IgE, IgG, IgG-1 and IL-4 in the A1A2 milk group GSH in the A2A2 milk group BCM-7 in the A1A2 milk group IgE, IgG, IgG-1 and IL-4 in the A1A2 milk group GSH in the A2A2 milk group BCM-7 in the A1A2 milk group frequency of stool in A2A2 milk group frequency of stool in A2A2 milk groupImproved consistency of stool in the A2A2 milk group Improvements in accuracy of the Cognitive Impairment Test in the A2A2 milk group | Sheng et al., 2019 [8] | ||
| Lactose-intolerant adults of various ethnicities (n = 25) | A2A2 group A1A2 group (25%A1/75%A2) A1A2 group (75%A1/25%A2) Lactose-free A1A2 group | 245 mL single-meal; Double-blind, randomized and crossover | Gastrointestinal symptoms over 6 h | Abdominal pain in the A2A2 group | Ramakrishnan et al., 2020 [45] | |||
| Chinese lactose-intolerant adults (n = 600) | A1A2 milk group A2A2 milk group | 300 mL single-meal; Double-blind, randomized and crossover | Gastrointestinal symptoms over 1 h, 3 h and 12 h | Feces | Abdominal pain, borborborygmus, flatulence, bloating, frequency and consistency of stool after 3 h in the A2A2 group | He et al., 2017 [69] | ||
| Australian adults (n = 37) | A1A1 milk group A2A2 milk group | 750 mL/dia, 2 weeks of intervention; Double-blind, randomized and crossover | Symptoms and transit time gastrointestinal | Feces | Results of stool consistency (Bristol Stool Scale) in the A1A1 milk group | Ho et al., 2014 [47] | ||
| Chinese lactose-intolerant adults (n = 45) | A1A2 milk group A2A2 milk group | 250 mL twice daily 2 weeks of intervention; Double-blind, randomized and crossover | Symptoms and transit time gastrointestinal, GSH, MPO, IgG, IgG, IgE, IL-4 and PCR | Gastrointestinal photograph, blood and feces | IgE, IgG, IgG-1 and IL-4 in A1A2 milk group gastrointestinal transit time A1A2 milk group A2A2 milk intake was not associated with increased gastrointestinal symptoms PCR did not differ between groups | Sun et al., 2015 [7] | ||
| New Zealander lactose-intolerant women (n= 40) | A1A2 milk group A2A2 milk group Lactose-free A1A2 group | 750 mL single-meal; Double-blind and randomized | Gastrointestinal symptoms over 3 h Bowel movements | Symptoms of nausea and fecal urgency in A2A2 and lactose-free milk group Bowel movement frequency does not differ between groups | Milan et al., 2020 [68] | |||
| Children with functional chronic constipation 1–12 y (n = 39) | A1A2 milk group A2A2 milk group | 400 mL/dia, 2 weeks of intervention | Gastrointestinal discomforts | Blood and feces | There was no effect of the casein type | Crowley et al., 2013 [70] | ||
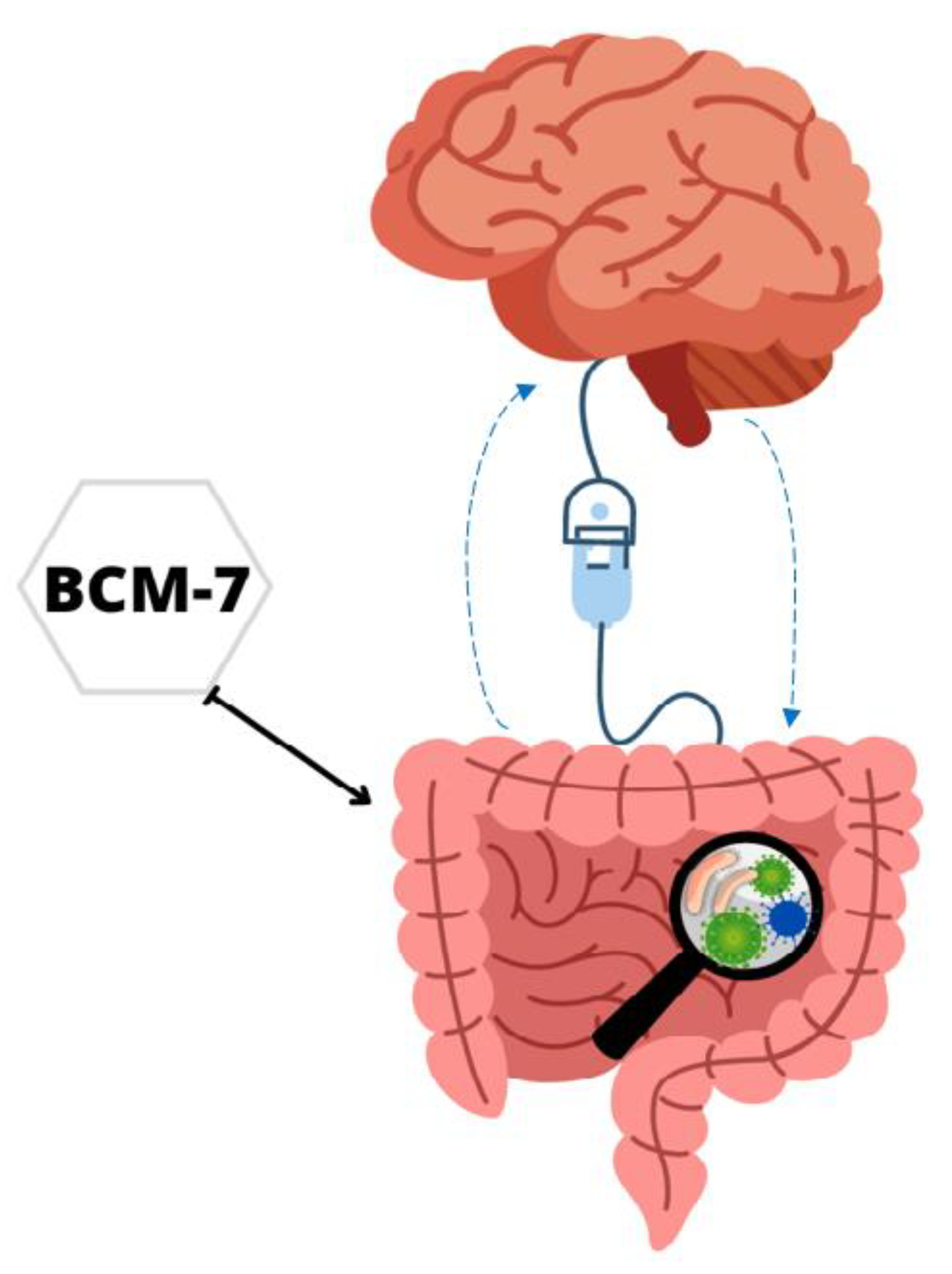
7. Gut–Brain Axis
8. Concluding Remarks and Future Directions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, N.W.; Fletcher, A.J.; Hill, J.P.; McNabb, W.C. Modeling the Contribution of Milk to Global Nutrition. Front. Nutr. 2022, 8, 716100. [Google Scholar] [CrossRef] [PubMed]
- Tulipano, G. Role of Bioactive Peptide Sequences in the Potential Impact of Dairy Protein Intake on Metabolic Health. Int. J. Mol. Sci. 2020, 21, 8881. [Google Scholar] [CrossRef] [PubMed]
- Asledottir, T.; Le, T.T.; Poulsen, N.A.; Devold, T.G.; Larsen, L.B.; Vegarud, G.E. Release of β-Casomorphin-7 from Bovine Milk of Different β-Casein Variants after Ex Vivo Gastrointestinal Digestion. Int. Dairy J. 2018, 81, 8–11. [Google Scholar] [CrossRef]
- Uzma, S.; Gill, H.; Chandrapala, J. Casein Micelles as an Emerging Delivery System for Bioactive Food Components. Foods 2021, 10, 1965. [Google Scholar] [CrossRef]
- Hannelore, D.; Vohwinkel, M.; Rehner, G. Effect of Casein and β-Casomorphins on Gastrointestinal Motility in Rats. J. Nutr. 1990, 120, 252–257. [Google Scholar] [CrossRef]
- Haq MR, U.; Kapila, R.; Saliganti, V. Consumption of β-Casomorphins-7/5 Induce Inflammatory Immune Response in Mice Gut through Th2 Pathway. J. Funct. Foods 2014, 8, 150–160. [Google Scholar] [CrossRef]
- Jianqin, S.; Leiming, X.; Lu, X.; Yelland, G.W.; Ni, J.; Clarke, A.J. Effects of Milk Containing Only A2 Beta Casein versus Milk Containing Both A1 and A2 Beta Casein Proteins on Gastrointestinal Physiology, Symptoms of Discomfort, and Cognitive Behavior of People with Self-Reported Intolerance to Traditional Cows’ Milk. Nutr. J. 2015, 15, 35. [Google Scholar] [CrossRef]
- Sheng, X.; Li, Z.; Ni, J.; Yelland, G. Effects of Conventional Milk Versus Milk Containing Only A2 β-Casein on Digestion in Chinese Children: A Randomized Study. J. Pediatr. Gastroenterol. Nutr. 2019, 69, 375–382. [Google Scholar] [CrossRef]
- Shikha, Y.; Dev, N.; Yadav, S.; Gheware, A.; Kulshreshtha, A.; Sharma, P.; Singh, V.P. Oral Feeding of Cow Milk Containing A1 Variant of β Casein Induces Pulmonary Inflammation in Male Balb/c Mice. Sci. Rep. 2020, 10, 8053. [Google Scholar] [CrossRef]
- Andrea, S.; Di Frangia, F.; Marsan, P.A.; De Noni, I.; Malacarne, M. Occurrence, Biological Properties and Potential Effects on Human Health of β-Casomorphin 7: Current Knowledge and Concerns. Crit. Rev. Food Sci. Nutr. 2020, 60, 3705–3723. [Google Scholar] [CrossRef]
- Asledottir, T.; Picariello, G.; Mamone, G.; Ferranti, P.; Røseth, A.; Devold, T.G.; Vegarud, G.E. Degradation of β-Casomorphin-7 through in Vitro Gastrointestinal and Jejunal Brush Border Membrane Digestion. J. Dairy Sci. 2019, 102, 8622–8629. [Google Scholar] [CrossRef] [PubMed]
- Barnett, M.P.; McNabb, W.C.; Roy, N.C.; Woodford, K.B.; Clarke, A.J. Dietary A1 β -Casein Affects Gastrointestinal Transit Time, Dipeptidyl Peptidase-4 Activity, and Inflammatory Status Relative to A2 β -Casein in Wistar Rats. Int. J. Food Sci. Nutr. 2014, 65, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Barłowska, J.; Szwajkowska, M.; Litwińczuk, Z.; Król, J. Nutritional Value and Technological Suitability of Milk from Various Animal Species Used for Dairy Production. Compr. Rev. Food Sci. Food Saf. 2011, 10, 291–302. [Google Scholar] [CrossRef]
- De Kruif, C.G.; Holt, C. Casein Micelle Structure, Functions and Interactions. In Advanced Dairy Chemistry—1 Proteins; Springer US: Boston, MA, USA, 2003. [Google Scholar] [CrossRef]
- Amalfitano, N.; Rosa, G.J.; Cecchinato, A.; Bittante, G. Nonlinear Modeling to Describe the Pattern of 15 Milk Protein and Nonprotein Compounds over Lactation in Dairy Cows. J. Dairy Sci. 2021, 104, 10950–10969. [Google Scholar] [CrossRef]
- Davor, D.; Noel; McCarthy, A.; Vasiljevic, T. Bovine β-Casomorphins: Friends or Foes? A Comprehensive Assessment of Evidence from in Vitro and Ex Vivo Studies. Trends Food Sci. Technol. 2021, 116, 681–700. [Google Scholar] [CrossRef]
- Monique, R.; Elnitski, L.; Miller, W.; Rosen, J.M. Multispecies Comparative Analysis of a Mammalian-Specific Genomic Domain Encoding Secretory Proteins. Genomics 2003, 82, 417–432. [Google Scholar] [CrossRef]
- Chen, Y.; Qu, S.; Huang, Z.; Ren, Y.; Wang, L.; Rankin, S.A. Analysis and Comparison of Key Proteins in Maiwa Yak and Bovine Milk Using High-Performance Liquid Chromatography Mass Spectrometry. J. Dairy Sci. 2021, 104, 8661–8672. [Google Scholar] [CrossRef]
- Oliver, C.J.; Ibeagha-Awemu, E.M.; Özbeyaz, C.; Zaragoza, P.; Williams, J.L.; Ajmone-Marsan, P.; Lenstra, J.A.; Moazami-Goudarzi, K.; Erhardt, G. Geographic Distribution of Haplotype Diversity at the Bovine Casein Locus. Genet. Sel. Evol. 2004, 36, 243. [Google Scholar] [CrossRef]
- Caroli, A.M.; Chessa, S.; Erhardt, G.J. Invited Review: Milk Protein Polymorphisms in Cattle: Effect on Animal Breeding and Human Nutrition. J. Dairy Sci. 2009, 92, 5335–5352. [Google Scholar] [CrossRef]
- Gallinat, J.L.; Qanbari, S.; Drögemüller, C.; Pimentel, E.C.G.; Thaller, G.; Tetens, J. DNA-Based Identification of Novel Bovine Casein Gene Variants. J. Dairy Sci. 2013, 96, 699–709. [Google Scholar] [CrossRef]
- Aschaffenburg, R. Section G. Genetics. Genetic Variants of Milk Proteins: Their Breed Distribution. J. Dairy Res. 1968, 35, 447–460. [Google Scholar] [CrossRef]
- Nan, G.; Uniacke-Lowe, T.; O’Regan, J.; Faulkner, H.; Alan; Kelly, L. Effect of Protein Genotypes on Physicochemical Properties and Protein Functionality of Bovine Milk: A Review. Foods 2021, 10, 2409. [Google Scholar] [CrossRef]
- Shiven, P.; Shah, T.; Sabara, P.; Bhatia, D.; Panchal, K.; Italiya, J.; Koringa, P.; Rank, D.N. Understanding Functional Implication of β-Casein Gene Variants in Four Cattle Breeds Characterized Using AmpliSeq Approach. 3 Biotech 2020, 10, 414. [Google Scholar] [CrossRef]
- Kathiravan, R.S.; Vandana, C.M.; Malarmathi, M.; Chitra, R.; Murali, N.; Arthanarieswaran, M. A Study on β-Casein Gene Polymorphism in Crossbred Cattle and Murrah / Graded Murrah Buffalo in Tamil Nadu. Indian J. Anim. Res. 2021, 57, 825–830. [Google Scholar] [CrossRef]
- Cosenza, G.; Feligini, M.; Mancusi, A.; D’Avino, A.; Coletta, A.; Di Berardino, D.; Ramunno, L. Italian Mediterranean River Buffalo CSN2 Gene Structure and Promoter Analysis. Ital. J. Anim. Sci. 2009, 8 (Suppl. S2), 57–59. [Google Scholar] [CrossRef]
- Oliveira, L.S.; Alves, J.S.; Bastos, M.S.; Costa, R.B.; de Camargo, G.M. Do Non-Bovine Domestic Animals Produce A2 Milk?: An in Silico Analysis. Anim. Biotechnol. 2021, 34, 93–95. [Google Scholar] [CrossRef]
- Jaiswal, K.; De, S.; Sarsavan, A. Detection of Single Nucleotide Polymorphism by T-ARMS PCR of Cross Bred Cattle Karan Fries for A1, A2 Beta Casein Types. Int. J. Sci. Res. Biol. Sci. 2014, 1, 18–22. [Google Scholar]
- Nguyen, H.T.; Schwendel, H.; Harland, D.; Day, L. Differences in the Yoghurt Gel Microstructure and Physicochemical Properties of Bovine Milk Containing A1A1 and A2A2 β-Casein Phenotypes. Food Res. Int. 2018, 112, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Stanisław, K.; Cieślińska, A.; Kostyra, E. Polymorphism of Bovine Beta-Casein and Its Potential Effect on Human Health. J. Appl. Genet. 2007, 48, 189–198. [Google Scholar] [CrossRef]
- Teagan, E.; Krista; Dawson, L.; Jacqueline; Keenan, I.; Andrew; Day, S. A Simple Method to Generate β-Casomorphin-7 by in Vitro Digestion of Casein from Bovine Milk. J. Funct. Foods 2021, 85, 104631. [Google Scholar] [CrossRef]
- Woodford, K.B. Casomorphins and Gliadorphins Have Diverse Systemic Effects Spanning Gut, Brain and Internal Organs. Int. J. Environ. Res. Public Health 2021, 18, 7911. [Google Scholar] [CrossRef] [PubMed]
- Bielecka, M.; Cichosz, G.; Czeczot, H. Antioxidant, Antimicrobial and Anticarcinogenic Activities of Bovine Milk Proteins and Their Hydrolysates—A Review. Int. Dairy J. 2022, 127, 10. [Google Scholar] [CrossRef]
- Blanca, H.-L.; García-Nebot, M.J.; Fernández-Tomé, S.; Amigo, L.; Recio, I. Dairy Protein Hydrolysates: Peptides for Health Benefits. Int. Dairy J. 2014, 38, 82–100. [Google Scholar] [CrossRef]
- Doan, N.D.; Johnson, S.K.; Busetti, F.; Solah, V.A. Formation and Degradation of Beta-Casomorphins in Dairy Processing. Crit. Rev. Food Sci. Nutr. 2015, 55, 1955–1967. [Google Scholar] [CrossRef]
- Sultan, S.; Huma, N.; Butt, M.S.; Aleem, M.; Abbas, M. Therapeutic Potential of Dairy Bioactive Peptides: A Contemporary Perspective. Crit. Rev. Food Sci. Nutr. 2018, 58, 105–115. [Google Scholar] [CrossRef]
- Elisa, K.; Zukerman, R.; Eshraghi, R.S.; Mittal, J.; Deth, R.C.; Castejon, A.M.; Trivedi, M.; Mittal, R.; Eshraghi, A.A. Nutritional Interventions for Autism Spectrum Disorder. Nutr. Rev. 2020, 78, 515–531. [Google Scholar] [CrossRef]
- Richard, D.; Clarke, A.; Ni, J.; Trivedi, M. Clinical Evaluation of Glutathione Concentrations after Consumption of Milk Containing Different Subtypes of β-Casein: Results from a Randomized, Cross-over Clinical Trial. Nutr. J. 2015, 15, 82. [Google Scholar] [CrossRef]
- Boutrou, R.; Gaudichon, C.; Dupont, D.; Jardin, J.; Airinei, G.; Marsset-Baglieri, A.; Benamouzig, R.; Tomé, D.; Leonil, J. Sequential Release of Milk Protein-Derived Bioactive Peptides in the Jejunum in Healthy Humans. Am. J. Clin. Nutr. 2013, 97, 1314–1323. [Google Scholar] [CrossRef]
- Sebely, P.; Woodford, K.; Kukuljan, S.; Ho, S. Milk Intolerance, Beta-Casein and Lactose. Nutrients 2015, 7, 7285–7297. [Google Scholar] [CrossRef]
- Trivedi, M.S.; Shah, J.S.; Al-Mughairy, S.; Hodgson, N.W.; Simms, B.; Trooskens, G.A.; Van Criekinge, W.; Deth, R.C. Food-Derived Opioid Peptides Inhibit Cysteine Uptake with Redox and Epigenetic Consequences. J. Nutr. Biochem. 2014, 25, 1011–1018. [Google Scholar] [CrossRef]
- Muthu, T.; Venkidasamy, B.; Thirupathi, P.; Chung, I.-M.; Subramanian, U. β-Casomorphin: A Complete Health Perspective. Food Chem. 2021, 337, 127765. [Google Scholar] [CrossRef]
- Beata, J.; Bukało, M.; Fiedorowicz, E.; Cieślińska, A.; Kordulewska, N.K.; Moszyńska, M.; Świątecki, A.; Kostyra, E. Role of Milk-Derived Opioid Peptides and Proline Dipeptidyl Peptidase-4 in Autism Spectrum Disorders. Nutrients 2019, 11, 87. [Google Scholar] [CrossRef]
- Cieślińska, A.; Kostyra, E.; Kostyra, H.; Oleński, K.; Fiedorowicz, E.; Kamiński, S. Milk from Cows of Different β-Casein Genotypes as a Source of β-Casomorphin-7. Int. J. Food Sci. Nutr. 2012, 63, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, M.; Eaton, T.K.; Sermet, O.M.; Savaiano, D.A. Milk Containing A2 β-Casein ONLY, as a Single Meal, Causes Fewer Symptoms of Lactose Intolerance than Milk Containing A1 and A2 β-Caseins in Subjects with Lactose Maldigestion and Intolerance: A Randomized, Double-Blind, Crossover Trial. Nutrients 2020, 12, 3855. [Google Scholar] [CrossRef]
- Yining, X.; Shrestha, N.; Préat, V.; Beloqui, A. An Overview of in Vitro, Ex Vivo and in Vivo Models for Studying the Transport of Drugs across Intestinal Barriers. Adv. Drug Deliv. Rev. 2021, 175, 113795. [Google Scholar] [CrossRef]
- Ho, S.; Woodford, K.; Kukuljan, S.; Pal, S. Comparative Effects of A1 versus A2 Beta-Casein on Gastrointestinal Measures: A Blinded Randomised Cross-over Pilot Study. Eur. J. Clin. Nutr. 2014, 68, 994–1000. [Google Scholar] [CrossRef]
- Nguyen, D.D.; Busetti, F.; Smolenski, G.; Johnson, S.T.; Solah, V.A. Release of beta-casomorphins during in-vitro gastrointestinal digestion of reconstituted milk after heat treatment. LWT 2021, 136, 1, 110312. [Google Scholar] [CrossRef]
- Haq, U.; Raies, M.; Kapila, R.; Kapila, S. Release of β-Casomorphin-7/5 during Simulated Gastrointestinal Digestion of Milk β-Casein Variants from Indian Crossbred Cattle (Karan Fries). Food Chem. 2015, 168, 70–79. [Google Scholar] [CrossRef]
- Cattaneo, S.; Masotti, F.; Stuknytė, M.; De Noni, I. Impact of in Vitro Static Digestion Method on the Release of β-Casomorphin-7 from Bovine Milk and Cheeses with A1 or A2 β-Casein Phenotypes. Food Chem. 2023, 404, 13. [Google Scholar] [CrossRef]
- Yin, H.; Liu, J.-J.; Yang, D.; Xu, H.Q. Yaşlı Farelerde Bağırsak Mukozası Bağışıklığına β-Kazomorfin-7’nin Etkisi. Kafkas Univ. Vet. Fak. Derg. 2019, 25, 689–696. [Google Scholar] [CrossRef]
- Tim, L.; Broeren, S.; Heck, J.; Bragt, M.; Huppertz, T. Processing Affects Beta-Casomorphin Peptide Formation during Simulated Gastrointestinal Digestion in Both A1 and A2 Milk. Int. Dairy J. 2021, 121, 105099. [Google Scholar] [CrossRef]
- Valentina, P.; Stuknytė, M.; Masotti, F.; De Noni, I.; Cattaneo, S. Model Infant Biscuits Release the Opioid-Acting Peptides Milk β-Casomorphins and Gluten Exorphins after in Vitro Gastrointestinal Digestion. Food Chem. 2021, 362, 130262. [Google Scholar] [CrossRef]
- Guantario, B.; Giribaldi, M.; Devirgiliis, C.; Finamore, A.; Colombino, E.; Capucchio, M.T.; Evangelista, R.; Motta, V.; Zinno, P.; Cirrincione, S.; et al. A Comprehensive Evaluation of the Impact of Bovine Milk Containing Different Beta-Casein Profiles on Gut Health of Ageing Mice. Nutrients 2020, 12, 2147. [Google Scholar] [CrossRef]
- Samuel, F.-T.; Martínez-Maqueda, D.; Tabernero, M.; Largo, C.; Recio, I.; Miralles, B. Effect of the Long-Term Intake of a Casein Hydrolysate on Mucin Secretion and Gene Expression in the Rat Intestine. J. Funct. Foods 2017, 33, 176–180. [Google Scholar] [CrossRef]
- Chia, J.S.; McRae, J.L.; Enjapoori, A.K.; Lefèvre, C.M.; Kukuljan, S.; Dwyer, K.M. Dietary Cows’ Milk Protein A1 Beta-Casein Increases the Incidence of T1D in NOD Mice. Nutrients 2018, 10, 1291. [Google Scholar] [CrossRef]
- Claustre, J.; Toumi, F.; Trompette, A.; Jourdan, G.; Guignard, H.; Chayvialle, J.A.; Plaisancié, P. Effects of Peptides Derived from Dietary Proteins on Mucus Secretion in Rat Jejunum. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G521–G528. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.; Sandhu, K.V.; Bastiaanssen, T.F.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Hong, Y.; Miao, J.; Ma, C.; Sun, G.; Zhang, Y. β-Casomorphin-7 Cause Decreasing in Oxidative Stress and Inhibiting NF-ΚB-INOS-NO Signal Pathway in Pancreas of Diabetes Rats. J. Food Sci. 2012, 77, C278–C282. [Google Scholar] [CrossRef]
- Han, D.N.; Zhang, D.H.; Wang, L.P.; Zhang, Y.S. Protective Effect of β-Casomorphin-7 on Cardiomyopathy of Streptozotocin-Induced Diabetic Rats via Inhibition of Hyperglycemia and Oxidative Stress. Peptides 2013, 44, 120–126. [Google Scholar] [CrossRef]
- Thakur, N.; Chauhan, G.; Mishra, B.P.; Mendiratta, S.K.; Pattanaik, A.K.; Singh, T.U.; Karikalan, M.; Meshram, S.K.; Garg, L. Comparative Evaluation of Feeding Effects of A1 and A2 Cow Milk Derived Casein Hydrolysates in Diabetic Model of Rats. J. Funct. Foods 2020, 75, 104272. [Google Scholar] [CrossRef]
- Beales, P.E.; Elliott, R.B.; Flohe, S.; Hill, J.P.; Kolb, H.; Pozzilli, P.; Wang, G.-S.; Wasmuth, H.; Scott, F. A Multi-Centre, Blinded International Trial of the Effect of A1 and A2 β-Casein Variants on Diabetes Incidence in Two Rodent Models of Spontaneous Type I Diabetes. Diabetologia 2002, 45, 1240–1246. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Cade, J.R. A Peptide Found in Schizophrenia and Autism Causes Behavioral Changes in Rats. Autism 1999, 3, 85–95. [Google Scholar] [CrossRef]
- Chin-Dusting, J.; Shennan, J.; Jones, E.; Williams, C.; Kingwell, B.; Dart, A. Effect of Dietary Supplementation with Βcasein A1 or A2 on Markers of Disease Development in Individuals at High Risk of Cardiovascular Disease. Br. J. Nutr. 2006, 95, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, O.; Kost, N.; Andreeva, O.; Korneeva, E.; Meshavkin, V.; Tarakanova, Y.; Dadayan, A.; Zolotarev, Y.; Grachev, S.; Mikheeva, I.; et al. Autistic Children Display Elevated Urine Levels of Bovine Casomorphin-7 Immunoreactivity. Peptides 2014, 56, 68–71. [Google Scholar] [CrossRef]
- Cieślińska, A.; Sienkiewicz-Szłapka, E.; Wasilewska, J.; Fiedorowicz, E.; Chwała, B.; Moszyńska-Dumara, M.; Cieśliński, T.; Bukało, M.; Kostyra, E. Influence of Candidate Polymorphisms on the Dipeptidyl Peptidase IV and μ-Opioid Receptor Genes Expression in Aspect of the β-Casomorphin-7 Modulation Functions in Autism. Peptides 2015, 65, 6–11. [Google Scholar] [CrossRef]
- El-Alameey, I.R.; Ahmed, H.H.; Eid, I.M.; El-Dory, G.H.A.D.A.; Gameel, M.A.N.A.L. Relationship of serum dipeptidyl peptidase-iv activity and anti-casein antibodies to gastrointestinal symptoms among children with autism spectrum disorder: An Egyptian study. Asian J. Pharm. Clin. Res. 2018, 11, 370–374. [Google Scholar] [CrossRef]
- Milan, A.M.; Shrestha, A.; Karlström, H.J.; Martinsson, J.A.; Nilsson, N.J.; Perry, J.K.; Day, L.; Barnett, M.P.G.; Cameron-Smith, D. Comparison of the Impact of Bovine Milk β-Casein Variants on Digestive Comfort in Females Self-Reporting Dairy Intolerance: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2020, 111, 149–160. [Google Scholar] [CrossRef]
- He, M.; Sun, J.; Jiang, Z.Q.; Yang, Y.X. Effects of Cow’s Milk Beta-Casein Variants on Symptoms of Milk Intolerance in Chinese Adults: A Multicentre, Randomised Controlled Study. Nutr. J. 2017, 16, 72. [Google Scholar] [CrossRef]
- Crowley, E.; Williams, L.; Roberts, T.; Dunstan, R.; Jones, P. Does Milk Cause Constipation? A Crossover Dietary Trial. Nutrients 2013, 5, 253–266. [Google Scholar] [CrossRef]
- Davor, D.; Noel; McCarthy, A.; Markoska, T.; Martin; Auldist, J.; Vasiljevic, T. Conformational and Physicochemical Characteristics of Bovine Skim Milk Obtained from Cows with Different Genetic Variants of β-Casein. Food Hydrocoll. 2022, 124, 107186. [Google Scholar] [CrossRef]
- Elliott, R.B.; Harris, D.P.; Hill, J.P.; Bibby, N.J.; Wasmuth, H.E. Type I (Insulin-Dependent) Diabetes Mellitus and Cow Milk: Casein Variant Consumption. Diabetologia 1999, 42, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.J.; Gregory, G.T.; Clarke, A.J. Health Implications of Milk Containing β-Casein with the A 2 Genetic Variant. Crit. Rev. Food Sci. Nutr. 2006, 46, 93–100. [Google Scholar] [CrossRef] [PubMed]
- González Olmo, B.M.; Butler, M.J.; Barrientos, R.M. Evolution of the Human Diet and Its Impact on Gut Microbiota, Immune Responses, and Brain Health. Nutrients 2021, 13, 196. [Google Scholar] [CrossRef] [PubMed]
- Boushra, D.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The Role of Short-Chain Fatty Acids in Microbiota-Gut-Brain Communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Aya, O.; Zuffa, S.; Walton, G.; Fagbodun, E.; Zanos, P.; Georgiou, P.; Kitchen, I.; Swann, J.; Bailey, A. Post-Weaning A1/A2 β-Casein Milk Intake Modulates Depressive-like Behavior, Brain μ-Opioid Receptors, and the Metabolome of Rats. IScience 2021, 24, 103048. [Google Scholar] [CrossRef]
- Kay SI, S.; Delgado, S.; Mittal, J.; Eshraghi, R.S.; Mittal, R.; Eshraghi, A.A. Beneficial Effects of Milk Having A2 β-Casein Protein: Myth or Reality? J. Nutr. 2021, 151, 1061–1072. [Google Scholar] [CrossRef]



| Source: β-CN Bovine | Peptide Sequence | Position |
|---|---|---|
| β-Casomorphin-4 | Tyr-Pro-Phe-Pro | 60–63 |
| β-Casomorphin-5 | Tyr-Pro-Phe-Pro-Gly | 60–64 |
| β-Casomorphin-6 | Tyr-Pro-Phe-Pro-Gly-Pro | 60–65 |
| β-Casomorphin-7 | Tyr-Pro-Phe-Pro-Gly-Pro-Ile | 60–66 |
| β-Casomorphin-8 | Tyr-Pro-Phe-Pro-Gly-Pro-Ile-Pro | 60–67 |
| β-Casomorphin-9 | Tyr-Pro-Phe-Pro-Gly-Pro-Ile-Pro-Asn | 60–68 |
| β-Casomorphin-10 | Tyr-Pro-Phe-Pro-Gly-Pro-Ile-Pro-Asn-Ser | 60–69 |
| β-Casomorphin-11 | Tyr-Pro-Phe-Pro-Gly-Pro-Ile-Pro-Asn-Ser-Leu | 60–70 |
| MODEL | Design/ Intervention | Dosage/ Length/ Administration | Measured Biomarkers | Samples | Significant Results/ Outcomes | Study | ||
|---|---|---|---|---|---|---|---|---|
| Male mice | BCM-7 group BCM-5 group Control group | Exposure of BCM-7 or BCM-5: 7.5 × 10−8 mol/day/animal for 15 days intubated orally Exposure with saline phosphate buffer: 200 μL for 15 days intubated orally | MPO, MCP-1, IL-4, Histamine, IgG, IgG1, IgG2a, IgE, IgA and goblet cells counting | Fluid and intestinal mucosa | Increase | BCM-7 | BCM-5 | Haq et al., 2014 [6] |
| MPO | 129.76% | 117.55% | ||||||
| MCP-1 | 33.38% | 31.73% | ||||||
| IL-4 | 175.54% | 164% | ||||||
| Histamine | 167.59% | 189.21% | ||||||
| IgE | 77.09% | 52.37% | ||||||
| IgG | 42.13% | 45.17% | ||||||
| IgG1 | 126.63% | 159.78% | ||||||
| IgG2a | 77.39% | 90.27% | ||||||
| Compared to the Control group There was no difference in IgA and goblet cells. | ||||||||
| Male mice 4 w (n = 48) | A1S group = plus saline + milk powder with A1 β-CN A2S group = plus saline + milk powder with A2 β-CN A1N group = Naloxone + powder milk with A1 β-CN A2N group = Naloxone + powder milk with or A2 β-CN | A1 skim milk powder, 475 g total to 36 h or 84 h A2 skim milk powder, 468 g total to 36 h or 84 h | Gastrointestinal transit time, MPO And DPP-IV | Urine, feces, blood and intestinal mucosa | MPO activity between 64 and 65% in all groups, except in the A2N group DPP-IV activity in 37 and 40% in A1N and A1S groups, respectively, compared to A2S group GI transit time was higher for the A1S and A1N groups | Barnett et al., 2014 [12] | ||
| Elderly mice (n = 50) | Control group young mice Control group age BCM-7 group (low, medium and high dose) | Control group young and aged mice: Exposure to saline physiological solution for 30 days through enteral administration BCM-7 for 30 days through enteral administration Low: 2 × 10−7 mol/day Medium: 1 × 10−6 mol/day High: 5 × 10−6 mol/day | IL-2, TNF-α, IgA, SOD, CAT and MDA | Intestinal mucosa | IL-2 level in the aged control group compared to the young control and low-BCM-7 groups IgA level in the aged control group compared to the young control, medium- and high-BCM-7 groups TNF-α level in the aged control group compared to the young control and low-BCM-7 groups SOD in low- and medium-BCM-7 groups MDA: All BCM-7 groups | Yin et al., 2019 [51] | ||
| Elderly mice (n = 24) | Control group A1A2 milk group A2A2 milk group | 4 weeks of intervention 120 g of lyophilized milk | MPO, DPP-IV, TNF-α, IL-6 and IgG | Feces, blood, fluid and intestinal mucosa | MPO, DPP-IV, TNF-α, IL-6 and IgG do not differ among the three groups The fecal microbiota found was different between A2A2 and A1A2 groups | Guantario et al., 2020 [54] | ||
| Male rats 8 w (n = 18) | Control group Hydrolyzed casein group | 8 weeks of intervention Hydrolyzed casein group: 0.4 g hydrolysate/kg/day/ animal | Mucin, IL-6, TNF-α, glucagon, leptin and insulin | Feces and intestin | mucin excretion after 2 w (29%) and 8 w (47%) in Hydrolyzed casein group IL-6, TNF-α, glucagon, leptin, and insulin no difference between the groups | Fernández-Tomé et al., 2017 [55] | ||
| Young mice 3–4 w | A1 milk for five generations A2 milk for five generations | The mice were fed ad libitum over a period of 30 weeks | Insulin, glucose, incidence of diabetes and immune profile | Feces, blood, lymphatic tissue and Intestinal integrity | In the F3 generation, at 30 weeks, the incidence of diabetes was doubled after the intake of beta-casein A1 relative to the intake of A2 (A1: 40% vs. A2: 20.7%) In F4 mice, subclinical insulitis and glucose metabolism bias were evident in 10-week-old mice that only received A1 milk | Chia et al., 2018 [56] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Vasconcelos, M.L.; Oliveira, L.M.F.S.; Hill, J.P.; Vidal, A.M.C. Difficulties in Establishing the Adverse Effects of β-Casomorphin-7 Released from β-Casein Variants—A Review. Foods 2023, 12, 3151. https://doi.org/10.3390/foods12173151
de Vasconcelos ML, Oliveira LMFS, Hill JP, Vidal AMC. Difficulties in Establishing the Adverse Effects of β-Casomorphin-7 Released from β-Casein Variants—A Review. Foods. 2023; 12(17):3151. https://doi.org/10.3390/foods12173151
Chicago/Turabian Stylede Vasconcelos, Marta Liliane, Luisa Maria F. S. Oliveira, Jeremy Paul Hill, and Ana Maria Centola Vidal. 2023. "Difficulties in Establishing the Adverse Effects of β-Casomorphin-7 Released from β-Casein Variants—A Review" Foods 12, no. 17: 3151. https://doi.org/10.3390/foods12173151