
Genetically Modified Rice Is Associated with Hunger, Health, and Climate Resilience

Abstract
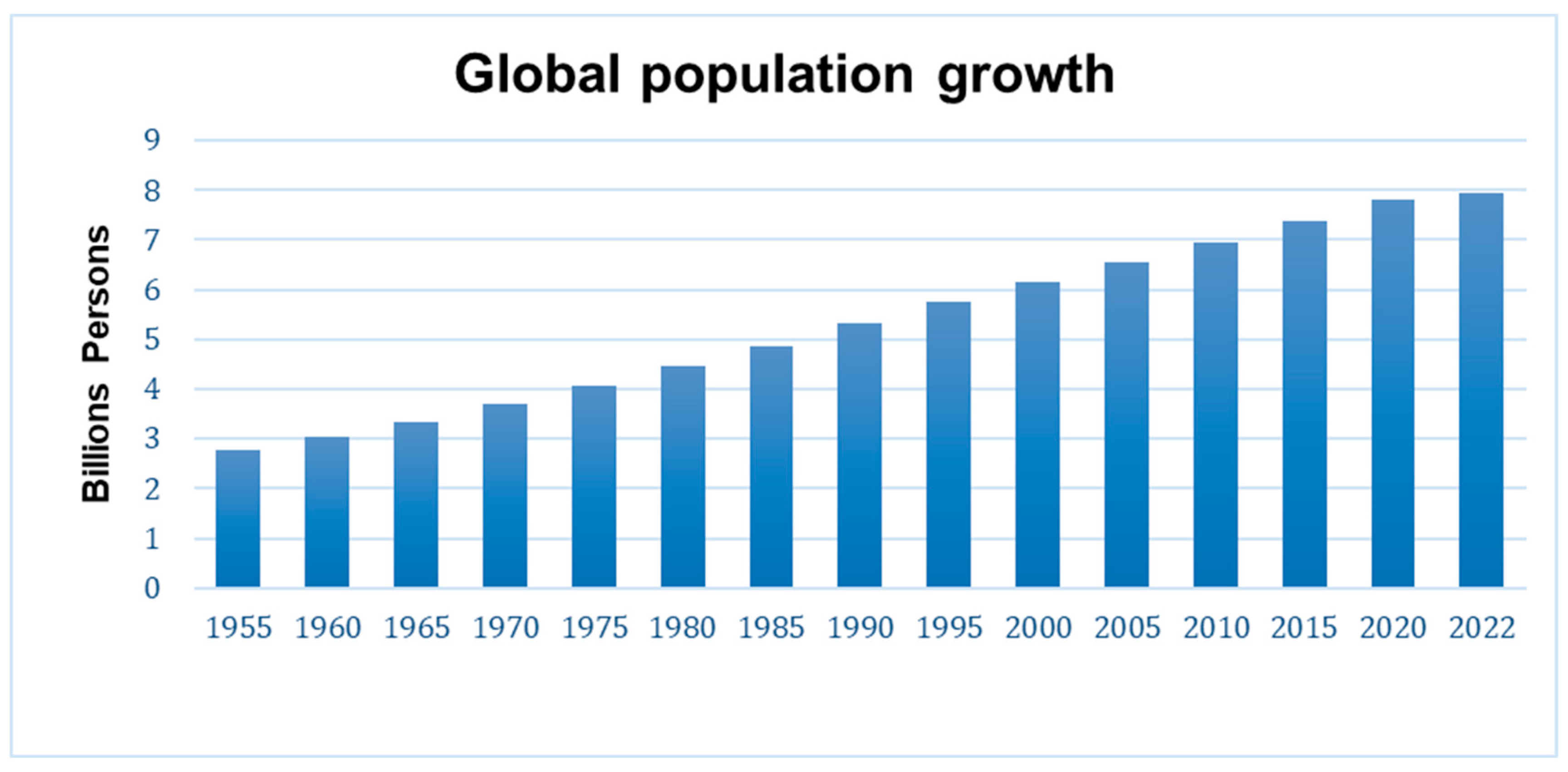
:1. Introduction


2. A Potential Solution to Hunger
2.1. Increasing the Grain Size of Rice
2.2. Increasing the Yield of Rice
2.3. Enhancing the Nutrient Content of Rice Grains
Enhancing the Nutritive Value of Rice Grains
3. Potential Health Benefits
3.1. Bioactive Compounds
3.2. Antioxidant Activity
3.3. Anti-Diabetic Activity
3.4. Anti-Inflammatory Activity
4. Addressing Climate Change
5. Implications, Limitations, and Future Research
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nair, K.P. Chapter four—Utilizing crop wild relatives to combat global warming. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 153, pp. 175–258. [Google Scholar]
- Hasan, K.; Tanaka, T.S.T.; Alam, M.; Ali, R.; Saha, C.K. Impact of Modern Rice Harvesting Practices over Traditional Ones. Rev. Agric. Sci. 2020, 8, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Krishna, K.R. Agroecosystems: Soils, Climate, Crops, Nutrient Dynamics and Productivity; CRC Press: Boca Raton, FL, USA, 2013; ISBN 978-1-4822-0342-4. [Google Scholar]
- Wijerathna-Yapa, A.; Pathirana, R. Sustainable Agro-Food Systems for Addressing Climate Change and Food Security. Agriculture 2022, 12, 1554. [Google Scholar] [CrossRef]
- Belahsen, R. Nutrition Transition and Food Sustainability. Proc. Nutr. Soc. 2014, 73, 385–388. [Google Scholar] [CrossRef]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The Impact of Climate Change on Agricultural Insect Pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Berry, H.L.; Ebi, K.; Bambrick, H.; Hu, W.; Green, D.; Hanna, E.; Wang, Z.; Butler, C.D. Climate Change, Food, Water and Population Health in China. Bull. World Health Organ. 2016, 94, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Klümper, W.; Qaim, M. A Meta-Analysis of the Impacts of Genetically Modified Crops. PLoS ONE 2014, 9, e111629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GM Approval Database|ISAAA.Org. Available online: https://www.isaaa.org/gmapprovaldatabase/ (accessed on 21 June 2023).
- Fraiture, M.-A.; Roosens, N.H.C.; Taverniers, I.; de Loose, M.; Deforce, D.; Herman, P. Biotech Rice: Current Developments and Future Detection Challenges in Food and Feed Chain. Trends Food Sci. Technol. 2016, 52, 66–79. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Shelton, A.; Ye, G. Insect-Resistant Genetically Modified Rice in China: From Research to Commercialization. Annu. Rev. Entomol. 2011, 56, 81–101. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Gambhir, G.; Dass, A.; Tripathi, A.K.; Singh, A.; Jha, A.K.; Yadava, P.; Choudhary, M.; Rakshit, S. Genetically Modified Crops: Current Status and Future Prospects. Planta 2020, 251, 91. [Google Scholar] [CrossRef]
- Liu, C.; Mao, B.; Yuan, D.; Chu, C.; Duan, M. Salt Tolerance in Rice: Physiological Responses and Molecular Mechanisms. Crop. J. 2022, 10, 13–25. [Google Scholar] [CrossRef]
- Endo, M.; Toki, S. Genome Editing in Rice. Rice 2020, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Zafar, K.; Sedeek, K.E.M.; Rao, G.S.; Khan, M.Z.; Amin, I.; Kamel, R.; Mukhtar, Z.; Zafar, M.; Mansoor, S.; Mahfouz, M.M. Genome Editing Technologies for Rice Improvement: Progress, Prospects, and Safety Concerns. Front. Genome Ed. 2020, 2, 5. [Google Scholar] [CrossRef]
- Gmr. Available online: http://igc.int/en/gmr_summary.aspx (accessed on 5 July 2023).
- World Population Dashboard. Available online: https://www.unfpa.org/data/world-population-dashboard (accessed on 9 April 2023).
- Atukunda, P.; Eide, W.B.; Kardel, K.R.; Iversen, P.O.; Westerberg, A.C. Unlocking the Potential for Achievement of the UN Sustainable Development Goal 2—‘Zero Hunger’—In Africa: Targets, Strategies, Synergies and Challenges. Food Nutr. Res. 2021, 65, 7686. [Google Scholar] [CrossRef]
- Song, X.J.; Kuroha, T.; Ayano, M.; Furuta, T.; Nagai, K.; Komeda, N.; Segami, S.; Miura, K.; Ogawa, D.; Kamura, T.; et al. Rare Allele of a Previously Unidentified Histone H4 Acetyltransferase Enhances Grain Weight, Yield, and Plant Biomass in Rice. Proc. Natl. Acad. Sci. USA 2015, 112, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Ren, Y.; Liu, L.; Wang, F.; Zhang, H.; Tian, P.; Pan, T.; Wang, Y.; Jing, R.; Liu, T.; et al. Ubiquitin Specific Protease 15 Has an Important Role in Regulating Grain Width and Size in Rice. Plant Physiol. 2019, 180, 381–391. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.L.; Dong, N.Q.; Guo, T.; Ye, W.W.; Shan, J.X.; Lin, H.X. A quantitative trait locus GW6 controls rice grain size and yield through the gibberellin pathway. Plant J. 2020, 103, 1174–1188. [Google Scholar] [CrossRef] [PubMed]
- Oliver, M.J. Why We Need GMO Crops in Agriculture. Mo Med. 2014, 111, 492–507. [Google Scholar] [PubMed]
- Chaudhari, P.R.; Tamrakar, N.; Singh, L.; Tandon, A.; Sharma, D. Rice Nutritional and Medicinal Properties: A Review Article. J. Pharmacogn. Phytochem. 2018, 7, 150–156. [Google Scholar]
- Jin, Y.; Drabik, D.; Heerink, N.; Wesseler, J. The Cost of Postponement of Bt Rice Commercialization in China. Front. Plant Sci. 2019, 10, 1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cromwell, G.L.; Henry, B.J.; Scott, A.L.; Gerngross, M.F.; Dusek, D.L.; Fletcher, D.W. Glufosinate Herbicide-Tolerant (LibertyLink) Rice vs. Conventional Rice in Diets for Growing-Finishing Swine. J. Anim. Sci. 2005, 83, 1068–1074. [Google Scholar] [CrossRef]
- Jin, M.; Chen, L.; Deng, X.W.; Tang, X. Development of Herbicide Resistance Genes and Their Application in Rice. Crop. J. 2022, 10, 26–35. [Google Scholar] [CrossRef]
- Muduli, L.; Pradhan, S.K.; Mishra, A.; Bastia, D.N.; Samal, K.C.; Agrawal, P.K.; Dash, M. Understanding Brown Planthopper Resistance in Rice: Genetics, Biochemical and Molecular Breeding Approaches. Rice Sci. 2021, 28, 532–546. [Google Scholar] [CrossRef]
- Wang, H.; Liao, Z.; Gao, Y.; Zhang, L.; Lei, W.; Huang, H.; Lei, S.; Jiang, M.; Chen, S.; Shi, L. Transposon Polymorphism and Its Potential Impacts on Brown Planthopper (Nilaparvata lugens Stål) Resistance in Rice (Oryza sativa L.). Agronomy 2023, 13, 1699. [Google Scholar] [CrossRef]
- Younas, M.U.; Wang, G.; Du, H.; Zhang, Y.; Ahmad, I.; Rajput, N.; Li, M.; Feng, Z.; Hu, K.; Khan, N.U.; et al. Approaches to Reduce Rice Blast Disease Using Knowledge from Host Resistance and Pathogen Pathogenicity. Int. J. Mol. Sci. 2023, 24, 4985. [Google Scholar] [CrossRef] [PubMed]
- Devanna, B.N.; Jain, P.; Solanke, A.U.; Das, A.; Thakur, S.; Singh, P.K.; Kumari, M.; Dubey, H.; Jaswal, R.; Pawar, D.; et al. Understanding the Dynamics of Blast Resistance in Rice-Magnaporthe Oryzae Interactions. J. Fungi 2022, 8, 584. [Google Scholar] [CrossRef]
- Yu, K.; Liu, Z.; Gui, H.; Geng, L.; Wei, J.; Liang, D.; Lv, J.; Xu, J.; Chen, X. Highly Efficient Generation of Bacterial Leaf Blight-Resistant and Transgene-Free Rice Using a Genome Editing and Multiplexed Selection System. BMC Plant Biol. 2021, 21, 197. [Google Scholar] [CrossRef]
- Jing, X.; Song, X.; Cai, S.; Wang, P.; Lu, G.; Yu, L.; Zhang, C.; Wu, Z. Overexpression of OsHAK5 Potassium Transporter Enhances Virus Resistance in Rice (Oryza sativa). Mol. Plant Pathol. 2022, 23, 1107–1121. [Google Scholar] [CrossRef]
- Toda, Y.; Hayashi, Y.; Suzuki, T.; Zeng, H.; Xiao, L.; Xiao, X.; Xu, J.; Guo, S.; Yan, F. Plasma Membrane H+-ATPase Overexpression Increases Rice Yield via Simultaneous Enhancement of Nutrient Uptake and Photosynthesis. Nat. Commun. 2021, 12, 735. [Google Scholar] [CrossRef]
- Kumari, V.V.; Banerjee, P.; Verma, V.C.; Sukumaran, S.; Chandran, M.A.S.; Gopinath, K.A.; Venkatesh, G.; Yadav, S.K.; Singh, V.K.; Awasthi, N.K. Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops. Int. J. Mol. Sci. 2022, 23, 8519. [Google Scholar] [CrossRef]
- Koza, N.A.; Adedayo, A.A.; Babalola, O.O.; Kappo, A.P. Microorganisms in Plant Growth and Development: Roles in Abiotic Stress Tolerance and Secondary Metabolites Secretion. Microorganisms 2022, 10, 1528. [Google Scholar] [CrossRef]
- Michalak, A.; Wdowikowska, A.; Janicka, M. Plant Plasma Membrane Proton Pump: One Protein with Multiple Functions. Cells 2022, 11, 4052. [Google Scholar] [CrossRef] [PubMed]
- Yuyu, C.; Aike, Z.; Pao, X.; Xiaoxia, W.; Yongrun, C.; Beifang, W.; Yue, Z.; Liaqat, S.; Shihua, C.; Liyong, C. Effects of GS3 and GL3.1 for Grain Size Editing by CRISPR/Cas9 in Rice. Rice Sci. 2020, 27, 405–413. [Google Scholar] [CrossRef]
- Li, Q.; Lu, L.; Liu, H.; Bai, X.; Zhou, X.; Wu, B.; Yuan, M.; Yang, L.; Xing, Y. A Minor QTL, SG3, Encoding an R2R3-MYB Protein, Negatively Controls Grain Length in Rice. Theor. Appl. Genet. 2020, 133, 2387–2399. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, K.; Qian, Q.; Liu, Q.; Li, Q.; Pan, Y.; Ye, Y.; Liu, X.; Wang, J.; Zhang, J.; et al. Non-Canonical Regulation of SPL Transcription Factors by a Human OTUB1-like Deubiquitinase Defines a New Plant Type Rice Associated with Higher Grain Yield. Cell Res. 2017, 27, 1142–1156. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Feng, F.; Zhang, Y.; Elesawi, I.E.; Xu, K.; Li, T.; Mei, H.; Liu, H.; Gao, N.; Chen, C.; et al. A Novel Rice Grain Size Gene OsSNB Was Identified by Genome-Wide Association Study in Natural Population. PLoS Genet. 2019, 15, e1008191. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Yuan, X.; Wang, Y.; Yang, Y.; Zhang, M.; Yu, H.; Xu, Q.; Wang, S.; Niu, X.; Wei, X. Validation of a QTL for Grain Size and Weight Using an Introgression Line from a Cross between Oryza Sativa and Oryza Minuta. Rice 2021, 14, 43. [Google Scholar] [CrossRef]
- Shim, K.-C.; Kim, S.H.; Jeon, Y.-A.; Lee, H.-S.; Adeva, C.; Kang, J.-W.; Kim, H.-J.; Tai, T.H.; Ahn, S.-N. A RING-Type E3 Ubiquitin Ligase, OsGW2, Controls Chlorophyll Content and Dark-Induced Senescence in Rice. Int. J. Mol. Sci. 2020, 21, 1704. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Zhang, Y.; Wang, C. Molecular Functions of Genes Related to Grain Shape in Rice. Breed. Sci. 2015, 65, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wei, Y.; Li, J.; Yang, F.; Chen, Y.; Chen, Y.; Guo, S.; Sha, A. Identification of QTL TGW12 Responsible for Grain Weight in Rice Based on Recombinant Inbred Line Population Crossed by Wild Rice (Oryza minuta) Introgression Line K1561 and Indica Rice G1025. BMC Genet. 2020, 21, 10. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Li, T.; Hao, C.; Wang, Y.; Chen, X.; Zhang, X. TaGS5-3A, a Grain Size Gene Selected during Wheat Improvement for Larger Kernel and Yield. Plant Biotechnol. J. 2016, 14, 1269–1280. [Google Scholar] [CrossRef]
- Li, S.; Gao, F.; Xie, K.; Zeng, X.; Cao, Y.; Zeng, J.; He, Z.; Ren, Y.; Li, W.; Deng, Q.; et al. The OsmiR396c-OsGRF4-OsGIF1 Regulatory Module Determines Grain Size and Yield in Rice. Plant Biotechnol. J. 2016, 14, 2134–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.-J.; Zhang, H.-P.; Cao, J.-J.; Zhu, X.-F.; Wang, S.-X.; Jiang, H.; Wu, Z.Y.; Lu, J.; Chang, C.; Sun, G.-L.; et al. Characterization of an IAA-Glucose Hydrolase Gene TaTGW6 Associated with Grain Weight in Common Wheat (Triticum aestivum L.). Mol. Breed. 2016, 36, 25. [Google Scholar] [CrossRef]
- Ye, X.; Al-Babili, S.; Kloti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the Provitamin A (β-Carotene) Biosynthetic Pathway into (Carotenoid-Free) Rice Endosperm. Science 2000, 287, 303. [Google Scholar] [CrossRef] [Green Version]
- Römer, S.; Fraser, P.D.; Kiano, J.W.; Shipton, C.A.; Misawa, N.; Schuch, W.; Bramley, P.M. Elevation of the Provitamin A Content of Transgenic Tomato Plants. Nat. Biotechnol. 2000, 18, 666–669. [Google Scholar] [CrossRef]
- Saini, P.; Singh, C.; Kumar, P.; Bishnoi, S.; Francies, R. Breeding for Nutritional Quality Improvement in Field Crops; MD Publishers New Delhi: New Delhi, Indina, 2020; pp. 200–264. ISBN 978-93-89235-25-8. [Google Scholar]
- Grune, T.; Lietz, G.; Palou, A.; Ross, A.C.; Stahl, W.; Tang, G.; Thurnham, D.; Yin, S.; Biesalski, H.K. β-Carotene Is an Important Vitamin A Source for Humans123. J. Nutr. 2010, 140, 2268S–2285S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, G.; Qin, J.; Dolnikowski, G.G.; Russell, R.M.; Grusak, M.A. Golden Rice Is an Effective Source of Vitamin A23. Am. J. Clin. Nutr. 2009, 89, 1776–1783. [Google Scholar] [CrossRef] [Green Version]
- Fiaz, S.; Ahmad, S.; Noor, M.A.; Wang, X.; Younas, A.; Riaz, A.; Riaz, A.; Ali, F. Applications of the CRISPR/Cas9 System for Rice Grain Quality Improvement: Perspectives and Opportunities. Int. J. Mol. Sci. 2019, 20, 888. [Google Scholar] [CrossRef] [Green Version]
- Han, F.; Han, F.; Wang, Y.; Fan, L.; Song, G.; Chen, X.; Jiang, P.; Miao, H.; Han, Y. Digestible Indispensable Amino Acid Scores of Nine Cooked Cereal Grains. Br. J. Nutr. 2019, 121, 30–41. [Google Scholar] [CrossRef]
- FAO. Dietary Protein Quality Evaluation in Human Nutrition: Report of an FAO Expert Consultation, 31 March-2 April, 2011, Auckland, New Zealand; Food and Agriculture Organization of the United Nations, Ed.; FAO Food and Nutrition Paper; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; ISBN 978-92-5-107417-6. [Google Scholar]
- Liu, X.; Zhang, C.; Wang, X.; Liu, Q.; Yuan, D.; Pan, G.; Sun, S.S.M.; Tu, J. Development of High-Lysine Rice via Endosperm-Specific Expression of a Foreign LYSINE RICH PROTEIN Gene. BMC Plant Biol. 2016, 16, 147. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Zhao, S.; Zhang, Y.; Yin, H.; Peng, X.; Cheng, Z.; Yang, Z.; Zheng, J. Production of Marker-Free Transgenic Rice (Oryza sativa L.) with Improved Nutritive Quality Expressing AmA1. Iran. J. Biotechnol. 2017, 15, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Cai, H.; Xiao, J.; Li, X.; Zhang, Q.; Lian, X. Over-Expression of Aspartate Aminotransferase Genes in Rice Resulted in Altered Nitrogen Metabolism and Increased Amino Acid Content in Seeds. Theor. Appl. Genet. 2009, 118, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.-Y.; Ma, A.; Xie, L.; Ramachandran, S. Improving Protein Content and Quality by Over-Expressing Artificially Synthetic Fusion Proteins with High Lysine and Threonine Constituent in Rice Plants. Sci. Rep. 2016, 6, 34427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Zhang, C.; Chan, M.; Zhao, D.; Chen, J.; Wang, Q.; Li, Q.; Yu, H.; Gu, M.; Sun, S.S.; et al. Biofortification of Rice with the Essential Amino Acid Lysine: Molecular Characterization, Nutritional Evaluation, and Field Performance. J. Exp. Bot. 2016, 67, 4285–4296. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, Y.; Bhullar, N.K. Molecular processes in iron and zinc homeostasis and their modulation for biofortification in rice. J. Integr. Plant Biol. 2018, 60, 1181–1198. [Google Scholar] [CrossRef]
- Ludwig, Y.; Slamet-Loedin, I.H. Genetic Biofortification to Enrich Rice and Wheat Grain Iron: From Genes to Product. Front. Plant Sci. 2019, 10, 833. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.A.T.; Kyriacou, B.; Callahan, D.L.; Carruthers, L.; Stangoulis, J.; Lombi, E.; Tester, M. Constitutive Overexpression of the OsNAS Gene Family Reveals Single-Gene Strategies for Effective Iron- and Zinc-Biofortification of Rice Endosperm. PLoS ONE 2011, 6, e24476. [Google Scholar] [CrossRef] [Green Version]
- Shahbaz, U.; Yu, X.-B.; Akhtar, W.; Ndagijimana, R.; Rauf, H. Golden Rice to Eradicate the Vitamin A Deficiency in the Developing Countries. Eur. J. Nutr. Food Saf. 2020, 12, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Verma, D.K.; Srivastav, P.P. Bioactive Compounds of Rice (Oryza sativa L.): Review on Paradigm and Its Potential Benefit in Human Health. Trends Food Sci. Technol. 2020, 97, 355–365. [Google Scholar] [CrossRef]
- Xu, K.; Zhang, B.; Liu, Y.; Mi, B.; Wang, Y.; Shen, Y.; Shi, G.; Dang, S.; Liu, X.; Yan, H. Staple Food Preference and Obesity Phenotypes: The Regional Ethnic Cohort Study in Northwest China. Nutrients 2022, 14, 5243. [Google Scholar] [CrossRef]
- Juliano, B.O. Rice: Role in diet. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; pp. 641–645. ISBN 978-0-12-384953-3. [Google Scholar]
- Su, X.; Xu, J.; Rhodes, D.; Shen, Y.; Song, W.; Katz, B.; Tomich, J.; Wang, W. Identification and Quantification of Anthocyanins in Transgenic Purple Tomato. Food Chem. 2016, 202, 184–188. [Google Scholar] [CrossRef] [Green Version]
- Ayella, A.K.; Trick, H.N.; Wang, W. Enhancing Lignan Biosynthesis by Over-Expressing Pinoresinol Lariciresinol Reductase in Transgenic Wheat. Mol. Nutr. Food Res. 2007, 51, 1518–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravichanthiran, K.; Ma, Z.F.; Zhang, H.; Cao, Y.; Wang, C.W.; Muhammad, S.; Aglago, E.K.; Zhang, Y.; Jin, Y.; Pan, B. Phytochemical Profile of Brown Rice and Its Nutrigenomic Implications. Antioxidants 2018, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Hosseini Dastgerdi, A.; Sharifi, M.; Soltani, N. GABA Administration Improves Liver Function and Insulin Resistance in Offspring of Type 2 Diabetic Rats. Sci. Rep. 2021, 11, 23155. [Google Scholar] [CrossRef] [PubMed]
- Imam, M.U.; Azmi, N.H.; Bhanger, M.I.; Ismail, N.; Ismail, M. Antidiabetic Properties of Germinated Brown Rice: A Systematic Review. Evid.-Based Complement. Altern. Med. 2012, 2012, 816501. [Google Scholar] [CrossRef]
- Kumar, A.; Agarwal, D.K.; Kumar, S.; Reddy, Y.M.; Chintagunta, A.D.; Saritha, K.V.; Pal, G.; Kumar, S.P.J. Nutraceuticals Derived from Seed Storage Proteins: Implications for Health Wellness. Biocatal. Agric. Biotechnol. 2019, 17, 710–719. [Google Scholar] [CrossRef]
- Morita, H.; Uno, Y.; Umemoto, T.; Sugiyama, C.; Matsumoto, M.; Wada, Y.; Ishizuka, T. Effect of gamma-aminobutyric acid-rich germinated brown rice on indexes of life-style related diseases. Nihon Ronen Igakkai Zasshi Jpn. J. Geriatr. 2004, 41, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazemzadeh, M.; Safavi, S.M.; Nematollahi, S.; Nourieh, Z. Effect of Brown Rice Consumption on Inflammatory Marker and Cardiovascular Risk Factors among Overweight and Obese Non-Menopausal Female Adults. Int. J. Prev. Med. 2014, 5, 478–488. [Google Scholar]
- Ronchetti, D.; Borghi, V.; Gaitan, G.; Herrero, J.F.; Impagnatiello, F. NCX 2057, a Novel NO-Releasing Derivative of Ferulic Acid, Suppresses Inflammatory and Nociceptive Responses in in Vitro and in Vivo Models. Br. J. Pharmacol. 2009, 158, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Hafeez, U.; Ali, M.; Hassan, S.M.; Akram, M.A.; Zafar, A. Advances in Breeding and Engineering Climate-Resilient Crops: A Comprehensive Review. Int. J. Res. Adv. Agric. Sci. 2023, 2, 85–99. [Google Scholar]
- Haider, Z.; Ahmad, I.; Zia, S.; Gan, Y. Recent Developments in Rice Molecular Breeding for Tolerance to Heavy Metal Toxicity. Agriculture 2023, 13, 944. [Google Scholar] [CrossRef]
- Giri, J.; Parida, S.K.; Raghuvanshi, S.; Tyagi, A.K. Emerging Molecular Strategies for Improving Rice Drought Tolerance. Curr. Genom. 2021, 22, 16–25. [Google Scholar] [CrossRef]
- Guo, L.-M.; Li, J.; He, J.; Liu, H.; Zhang, H.-M. A Class I Cytosolic HSP20 of Rice Enhances Heat and Salt Tolerance in Different Organisms. Sci. Rep. 2020, 10, 1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, C.; Ye, H.; Fan, M.; Pu, T.; Yan, J. The Rice Transcription Factors OsICE Confer Enhanced Cold Tolerance in Transgenic Arabidopsis. Plant Signal. Behav. 2017, 12, e1316442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perata, P. The Rice SUB1A Gene: Making Adaptation to Submergence and Post-Submergence Possible. Plant Cell Environ. 2018, 41, 717–720. [Google Scholar] [CrossRef]
- Blancquaert, D.; Daele, J.V.; Strobbe, S.; Kiekens, F.; Storozhenko, S.; Steur, H.D.; Gellynck, X.; Lambert, W.; Stove, C.; Straeten, D.V.D. Improving Folate (Vitamin B9) Stability in Biofortified Rice through Metabolic Engineering. Nat. Biotechnol. 2015, 33, 1076–1081. [Google Scholar] [CrossRef]
- Connorton, J.M.; Balk, J. Iron Biofortification of Staple Crops: Lessons and Challenges in Plant Genetics. Plant Cell Physiol. 2019, 60, 1447–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, M.; Turaidar, V.; Anantapur, K.K.N.R.; Bharani, S.S.; Dalawai, N. Enhancement of iron and zinc in rice grain through biofortification approach. Int. J. Curr. Microbiol. App. Sci. 2018, 7, 628–637. [Google Scholar]
- Lee, S.; An, G. Over-Expression of OsIRT1 Leads to Increased Iron and Zinc Accumulations in Rice. Plant Cell Environ. 2009, 32, 408–416. [Google Scholar] [CrossRef]
- Tozawa, Y.; Hasegawa, H.; Terakawa, T.; Wakasa, K. Characterization of Rice Anthranilate Synthase α-Subunit Genes OASA1 and OASA2. Tryptophan Accumulation in Transgenic Rice Expressing a Feedback-Insensitive Mutant of OASA1. Plant Physiol. 2001, 126, 1493–1506. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xia, Y.; Dong, Y.; Xie, T.; Sun, W.; Yu, S. Natural Variation of Fatty Acid Desaturase Gene Affects Linolenic Acid Content and Starch Pasting Viscosity in Rice Grains. Int. J. Mol. Sci. 2022, 23, 12055. [Google Scholar] [CrossRef]
- Ogo, Y.; Mori, T.; Nakabayashi, R.; Saito, K.; Takaiwa, F. Transgenic Rice Seed Expressing Flavonoid Biosynthetic Genes Accumulate Glycosylated and/or Acylated Flavonoids in Protein Bodies. J. Exp. Bot. 2016, 67, 95–106. [Google Scholar] [CrossRef] [PubMed]
- FAO. World Food and Agriculture—Statistical Yearbook 2022; FAO Statistical Yearbook—World Food and Agriculture; FAO: Rome, Italy, 2022; ISBN 978-92-5-136930-2. [Google Scholar]
- Itoh, M.; Sudo, S.; Mori, S.; Saito, H.; Yoshida, T.; Shiratori, Y.; Suga, S.; Yoshikawa, N.; Suzue, Y.; Mizukami, H.; et al. Mitigation of Methane Emissions from Paddy Fields by Prolonging Midseason Drainage. Agric. Ecosyst. Environ. 2011, 141, 359–372. [Google Scholar] [CrossRef]
- Su, J.; Hu, C.; Yan, X.; Jin, Y.; Chen, Z.; Guan, Q.; Wang, Y.; Zhong, D.; Jansson, C.; Wang, F.; et al. Expression of Barley SUSIBA2 Transcription Factor Yields High-Starch Low-Methane Rice. Nature 2015, 523, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Linquist, B.; van Groenigen, K.J.; Adviento-Borbe, M.A.; Pittelkow, C.; van Kessel, C. An Agronomic Assessment of Greenhouse Gas Emissions from Major Cereal Crops. Glob. Chang. Biol. 2012, 18, 194–209. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biotic Stress | Causes of the Stress | Transgenic Approach | References |
|---|---|---|---|
| Insect pests | Brown Planthopper (BPH) | Focusing on the identification of candidate insect resistance genes to provide initial materials for the breeding program of BPH-resistant rice varieties. | [27,28] |
| Bacterium Bacillus thuringiensis | By transferring genes from the soil bacterium Bacillus thuringiensis into the rice genome, genetically modified BT rice is resistant against lepidopteran pest damage and thus less required the amount of insecticide use. | [24] | |
| Fungus | Rice blast | Several qualitative resistance and quantitative resistance genes as well as several avirulence genes from the pathogen have been characterized. However, the population of rice blast fungus is constantly changing. Therefore, it is a challenge to control rice blast by developing an efficient surveillance system to monitor emerging new virulent strains. | [29,30] |
| Bacteria | Bacterial leaf blight | To improve rice resistance to bacteria, mutation of a key bacterial protein-binding site in the UPT box of Xa13 to abolish PXO99-induced Xa13 expression. | [31] |
| Viruses | Rice grassy stunt virus (RGSV) | Many studies have revealed a correlation between intracellular potassium levels and the incidence of virus-caused diseases. The induction of host resistance against RGSV infection in High-Affinity K+ Transporter 5 transgenic overexpression rice plants may involve the accumulation of reactive oxygen species (ROS). This is because RGSV infection, along with OsHAK5 overexpression, leads to an increase in ROS accumulation in rice leaves. | [32] |
| Herbicide toxicity | Phosphinothricin or Glufosinate | Phosphinothricin acetyl transferase (PAT) is a protein that plays a role in controlling resistance by detoxifying the Liberty herbicide molecule (glufosinate-ammonium). The production of the PAT enzyme enables the breakdown of Liberty before it can bind to the enzyme glutamine synthetase. Glutamine synthetase is responsible for facilitating the production of specific amino acids and recycling ammonium in plants. Hence, Liberty resistance in rice from the presence of a transgene introduces an additional enzymatic function. | [22,23] |
| DIAA Reference Ratio | Brown Rice | Polished Rice | Buckwheat | Oats | Adlay | Whole Wheat |
|---|---|---|---|---|---|---|
| Histidine | 0.99 | 0.77 | 0.89 | 0.86 | 0.8 | 0.97 |
| Threonine | 0.89 | 0.64 | 0.89 | 0.79 | 0.73 | 079 |
| Valine | 1.04 | 0.93 | 0.87 | 1.01 | 1.05 | 0.88 |
| Lysine | 0.42 | 0.37 | 0.7 | 0.43 | 0.13 | 0.2 |
| Isoleucine | 0.9 | 0.82 | 0.87 | 1.02 | 1.05 | 0.98 |
| Leucine | 0.97 | 0.84 | 0.82 | 1.07 | 2.01 | 0.96 |
| Tryptophan | 2.23 | 0.84 | 1.05 | 1.23 | 0.91 | 0.95 |
| Sulfur AA | 0.43 | 0.58 | 0.68 | 0.68 | 0.78 | 0.64 |
| Aromatic AA | 1.39 | 1.28 | 1.17 | 1.52 | 1.59 | 1.34 |
| DIAAS (%) | 42 (Lys) | 37 (Lys) | 68 (SAA) | 43 (Lys) | 13 (Lys) | 20 (Lys) |
| Crop | Target Tissue | Element | Baseline Concentration in Popular Cultivars (µg/g) | Target Concentration (µg/g) |
|---|---|---|---|---|
| Rice | Polished grains | Fe | 2 | 15 |
| Zn | 16 | 28 |
| Components | Amounts (per 100 g) | |
|---|---|---|
| Brown Rice | Milled Rice | |
| Carbohydrate (g) | 73–87 | 77–87 |
| Protein (g) | 7.1–8.3 | 6.3–7.1 |
| Fiber (g) | 2.9–4.4 | 0.7–2.7 |
| Fat (g) | 1.6–2.8 | 0.3–0.6 |
| Calcium (mg) | 10–50 | 10–30 |
| Phosphorus (mg) | 0.17–0.43 | 0.08–0.15 |
| Iron (mg) | 1.4–5.2 | 0.3–0.8 |
| Zinc (mg) | 1.9–2.8 | 0.8–2.3 |
| α-tocopherol (mg) | 0.8–2.5 | 0.1–0.3 |
| Phytic acid P (mg) | 0.13–2.7 | 0.02–0.07 |
| Thiamin (mg) | 0.4–0.6 | 0.07–0.17 |
| Riboflavin (mg) | 0.04–0.14 | 0.02–0.06 |
| Niacin (mg) | 3.5–6.2 | 1.3–2.5 |
| Pantothenic acid (mg) | 1.4–1.6 | 0.8–1.3 |
| Vitamin B6 (mg) | 0.5–0.7 | 0.1–0.4 |
| Folate (μg) | 16–20 | 4–9 |
| Rice Variety | Breeding Approach | Reference |
|---|---|---|
| Heavy metal-tolerant rice | Recent advancements in genome engineering and editing techniques have been successfully applied to rice to enhance metal tolerance and reduce the accumulation of heavy metals in rice. | [78] |
| Drought-tolerant rice | Marker-assisted selection for drought QTLs. | [79] |
| Heat- and salt-tolerant rice | Transgenic rice plants constitutively overexpressing OsHSP20. | [80] |
| Cold-tolerant rice | Introduced or enhanced the expression of CBF/DREB Genes. Manipulating the expression of ICE1 and ICE2 Genes. | [81] |
| Flood-tolerant rice | Submergence 1 (SUB1) gene has been identified and used in GM rice. | [82] |
| Biofortification | Breeding Approach | References |
|---|---|---|
| Beta-carotene Phytoene (precursor of beta-carotene) | Golden rice and its nutritional value; beta-carotene metabolism; genetic engineering provitamin | [50,52] |
| Folate (vitamin B9) | Folate fortification and stability; metabolic engineering | [83] |
| Iron | Nicotianamine aminotransferase genes; transgenic; multigene introduction; ferritin gene; endosperm biofortification | [84,85] |
| Zinc | Over-expression of OsIRT1; involvement of genes in phytosiderophore synthesis | [86] |
| High amino acid and protein content | Accumulation of glycinin with the glutelins; dihydrodipicolinate synthase gene; tryptophan accumulation; over-expression of aspartate aminotransferase genes | [60,87] |
| Alpha-linolenic acid | Microsomal omega-3 fatty acid desaturase gene | [88] |
| Flavonoids and antioxidants | Flavonoids synthesis in the endosperm; transgenic | [89] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, K.; Wang, X.; Wang, W. Genetically Modified Rice Is Associated with Hunger, Health, and Climate Resilience. Foods 2023, 12, 2776. https://doi.org/10.3390/foods12142776
Kobayashi K, Wang X, Wang W. Genetically Modified Rice Is Associated with Hunger, Health, and Climate Resilience. Foods. 2023; 12(14):2776. https://doi.org/10.3390/foods12142776
Chicago/Turabian StyleKobayashi, Kaori, Xiaohui Wang, and Weiqun Wang. 2023. "Genetically Modified Rice Is Associated with Hunger, Health, and Climate Resilience" Foods 12, no. 14: 2776. https://doi.org/10.3390/foods12142776