
Caffeine Synthesis and Its Mechanism and Application by Microbial Degradation, A Review
Abstract
:1. Introduction
2. Caffeine Biosynthesis in Plants and Its Ecological Functions
2.1. Pathways for Caffeine Synthesis in Plants
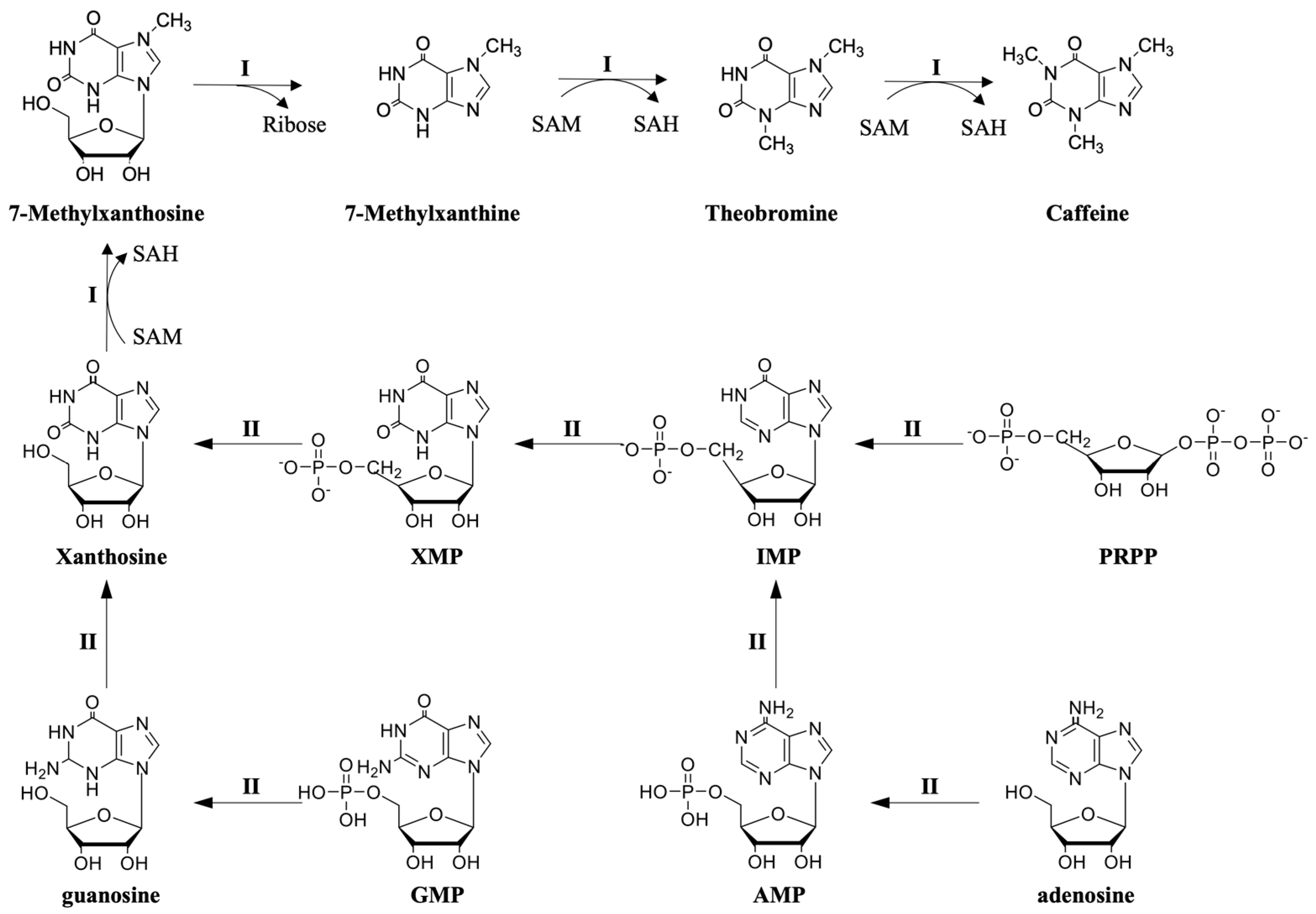
2.1.1. Caffeine Biosynthesis Pathways from Xanthosine in Plants
2.1.2. Pathways Supplying Xanthosine for Caffeine Biosynthesis in Plants
- The de novo route. The de novo route is the most significant synthesis pathway of xanthosine in caffeine production. This pathway starts from 5-phosphoribose-1-pyrophosphate (PRPP), which is transformed into inosine 5′-monophosphate (IMP) via a series of enzyme-catalyzed reactions and ultimately converted into xanthosine (the key enzymes involved in the conversion from IMP to xanthosine are IMP dehydrogenase and 5′-nucleotidase) [21].
- The AMP route. Xanthine nucleosides can be synthesized from adenine nucleosides in the following way: AMP → IMP → XMP → xanthosine (the key enzymes involved in the conversion from AMP to xanthosine are AMP deaminase, IMP dehydrogenase, and 5′-nucleotidase) [25].
- The SAM cycle route. The synthesis of xanthosine can also occur through S-adenosylhomocysteine (SAH) in the S-adenosylmethionine (SAM) cycle [26]. The pathway from SAH to xanthine nucleosides involves the following steps: SAH → adenosine → adenine → AMP → IMP → XMP → xanthosine (the key enzymes involved in the conversion from adenosine to xanthosine are adenosine nucleosidase, adenine phosphoribosyltransferase, AMP deaminase, IMP dehydrogenase, and 5′-nucleotidase).
2.2. Ecological Function of Caffeine
3. Caffeine Metabolism in Bacteria
3.1. N-Demethylation Pathway
3.2. C-8 Oxidation Pathway
4. Characteristics of Structures and Sequences from the Enzymes in the Bacterial Degradation Pathway of Caffeine
4.1. Structural Features of N-Demethylases
4.2. Gene Sequence Characteristics of N-Demethylases
4.3. Structure and Gene Sequence Characteristics of C-8 Dehydrogenase
5. Caffeine Metabolism in Fungi
6. Application of Caffeine Degradation by Microorganisms
6.1. Reuse of Coffee Wastes
6.2. Purifying Caffeine Wastewater
6.3. Production of Methylxanthine
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Runge, F. Neueste Phytochemische Entdeckungen Zur Begründung Einer Wissenschaftlichen Phytochemie. In Phytochemische Entdeckungen; Reimer: Berlin, Germany, 1820; p. 204. [Google Scholar]
- Von, G.F. Vermischte Notizen. 1. Kaffeestoff Und Salzgehalt Des Quassia Extrakts. Ann. Chem. 1820, 4, 240. [Google Scholar]
- Fredholm, B.B.; Bättig, K.; Holmén, J.; Nehlig, A.; Zvartau, E.E. Actions of Caffeine in the Brain with Special Reference to Factors That Contribute to Its Widespread Use. Pharmacol. Rev. 1999, 51, 83–133. [Google Scholar] [PubMed]
- Meeusen, R.; Roelands, B.; Spriet, L.L. Caffeine, Exercise and the Brain. Nestle Nutr. Inst. Workshop Ser. 2013, 76, 1–12. [Google Scholar] [PubMed] [Green Version]
- Marx, B.; Scuvée, É.; Scuvée-Moreau, J.; Seutin, V.; Jouret, F. Mechanisms of caffeine-induced diuresis. Med. Sci. 2016, 32, 485–490. [Google Scholar]
- Cheng, Y.-C.; Ding, Y.-M.; Hueng, D.-Y.; Chen, J.-Y.; Chen, Y. Caffeine Suppresses the Progression of Human Glioblastoma via Cathepsin B and MAPK Signaling Pathway. J. Nutr. Biochem. 2016, 33, 63–72. [Google Scholar] [CrossRef]
- Tej, G.N.V.C.; Nayak, P.K. Mechanistic Considerations in Chemotherapeutic Activity of Caffeine. Biomed. Pharmacother. 2018, 105, 312–319. [Google Scholar] [CrossRef]
- Bartolomeu, A.R.; Romualdo, G.R.; Lisón, C.G.; Besharat, Z.M.; Corrales, J.A.M.; Chaves, M.Á.G.; Barbisan, L.F. Caffeine and Chlorogenic Acid Combination Attenuate Early-Stage Chemically Induced Colon Carcinogenesis in Mice: Involvement of OncomiR MiR-21a-5p. Int. J. Mol. Sci. 2022, 23, 6292. [Google Scholar] [CrossRef]
- Sherman, S.M.; Buckley, T.P.; Baena, E.; Ryan, L. Caffeine Enhances Memory Performance in Young Adults during Their Non-Optimal Time of Day. Front. Psychol. 2016, 7, 1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nehlig, A. Is Caffeine A Cognitive Enhancer? J. Alzheimers Dis. 2010, 20, S85–S94. [Google Scholar] [CrossRef]
- Ngueta, G. Caffeine and Caffeine Metabolites in Relation to Hypertension in U.S. Adults. Eur. J. Clin. Nutr. 2020, 74, 77–86. [Google Scholar] [CrossRef]
- Kolb, H.; Martin, S.; Kempf, K. Coffee and Lower Risk of Type 2 Diabetes: Arguments for a Causal Relationship. Nutrients 2021, 13, 1144. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.M.M.; Lima, D.R.A. Coffee Consumption, Obesity and Type 2 Diabetes: A Mini-Review. Eur. J. Clin. Nutr. 2016, 55, 1345–1358. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Zhang, W.; Zhang, Y.; Zhang, M.; Zhang, Y.; Niu, X.; Zhao, Q.; Liu, Z.; Li, Y.; Diao, A. Caffeine Promotes the Expression of Telomerase Reverse Transcriptase to Regulate Cellular Senescence and Aging. Food Funct. 2021, 12, 2914–2924. [Google Scholar] [CrossRef]
- Fisone, G.; Borgkvist, A.; Usiello, A. Caffeine as a Psychomotor Stimulant: Mechanism of Action. Cell. Mol. Life Sci. 2004, 61, 857–872. [Google Scholar] [CrossRef]
- Derry, C.J.; Derry, S.; Moore, R.A. Caffeine as an Analgesic Adjuvant for Acute Pain in Adults. Cochrane Database Syst. Rev. 2012, 3, CD009281. [Google Scholar]
- Fujimaki, M.; Saiki, S.; Li, Y.; Kaga, N.; Taka, H.; Hatano, T.; Ishikawa, K.-I.; Oji, Y.; Mori, A.; Okuzumi, A.; et al. Serum Caffeine and Metabolites Are Reliable Biomarkers of Early Parkinson Disease. Neurology 2018, 90, e404–e411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggiero, M.; Calvello, R.; Porro, C.; Messina, G.; Cianciulli, A.; Panaro, M.A. Neurodegenerative Diseases: Can Caffeine Be a Powerful Ally to Weaken Neuroinflammation? Int. J. Mol. Sci. 2022, 23, 12958. [Google Scholar] [CrossRef] [PubMed]
- Socała, K.; Szopa, A.; Serefko, A.; Poleszak, E.; Wlaź, P. Neuroprotective Effects of Coffee Bioactive Compounds: A Review. Int. J. Mol. Sci. 2021, 22, 107. [Google Scholar] [CrossRef]
- Herman, A.; Herman, A.P. Caffeine’s Mechanisms of Action and Its Cosmetic Use. Skin Pharmacol. Phys. 2013, 26, 8–14. [Google Scholar] [CrossRef]
- Ashihara, H.; Mizuno, K.; Yokota, T.; Crozier, A. Xanthine Alkaloids: Occurrence, Biosynthesis, and Function in Plants. In Progress in the Chemistry of Organic Natural Products 105; Kinghorn, A.D., Falk, H., Gibbons, S., Kobayashi, J., Eds.; Progress in the Chemistry of Organic Natural Products; Springer International Publishing: Cham, Switzerland, 2017; Volume 105, pp. 1–88. ISBN 978-3-319-49711-2. [Google Scholar]
- Ashihara, H.; Sano, H.; Crozier, A. Caffeine and Related Purine Alkaloids: Biosynthesis, Catabolism, Function and Genetic Engineering. Phytochemistry 2008, 69, 841–856. [Google Scholar] [CrossRef]
- Ashihara, H.; Monteiro, A.M.; Gillies, F.M.; Crozier, A. Biosynthesis of Caffeine in Leaves of Coffee. Plant Physiol. 1996, 111, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T. The Participation of S-Adenosylmethionine in the Biosynthesis of Caffeine in the Tea Plant. FEBS Lett. 1972, 24, 18–20. [Google Scholar] [CrossRef] [Green Version]
- Ashihara, H. Purine Metabolism and the Biosynthesis of Caffeine in Maté Leaves. Phytochemistry 1993, 33, 1427–1430. [Google Scholar] [CrossRef]
- Koshiishi, C.; Kato, A.; Yama, S.; Crozier, A.; Ashihara, H. A New Caffeine Biosynthetic Pathway in Tea Leaves: Utilisation of Adenosine Released from the S-Adenosyl-L-Methionine Cycle. FEBS Lett. 2001, 499, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Ashihara, H.; Takasawa, Y.; Suzuki, T. Metabolic Fate of Guanosine in Higher Plants. Physiol. Plant. 1997, 100, 909–916. [Google Scholar] [CrossRef]
- Negishi, O.; Ozawa, T.; Imagawa, H. Guanosine Deaminase and Guanine Deaminase from Tea Leaves. Biosci. Biotechnol. Biochem. 1994, 58, 1277–1281. [Google Scholar] [CrossRef]
- Anaya, A.L. Metabolism and Ecology of Purine Alkaloids. Front. Biosci.-Landmark 2006, 11, 2354–2370. [Google Scholar] [CrossRef] [Green Version]
- Epifanio, J. Ecology of the Cafetalero Agroecosystem. Ph.D. Thesis, Universidad Nacional Autonoma de Mexico, Mexico City, DF, Mexico, 1820. [Google Scholar]
- Friedman, J.; Waller, G.R. Caffeine Hazards and Their Prevention in Germinating Seeds of Coffee (Coffea arabica L.). J. Chem. Ecol. 1983, 9, 1099–1106. [Google Scholar] [CrossRef]
- Nathanson, J.A. Caffeine and Related Methylxanthines: Possible Naturally Occurring Pesticides. Science 1984, 226, 184–187. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-S.; Sano, H. Pathogen Resistance of Transgenic Tobacco Plants Producing Caffeine. Phytochemistry 2008, 69, 882–888. [Google Scholar] [CrossRef]
- Hollingsworth, R.G.; Armstrong, J.W.; Campbell, E. Caffeine as a Repellent for Slugs and Snails. Nature 2002, 417, 915–916. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, A.; Sano, C.M.; Yazaki, K.; Sano, H. Caffeine Fostering of Mycoparasitic Fungi against Phytopathogens. Plant Singal. Behav. 2016, 11, e1113362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, F.E.; Emche, S.; Shao, J.; Simpkins, A.; Summers, R.M.; Mock, M.B.; Ebert, D.; Infante, F.; Aoki, S.; Maul, J.E. Cultivation and Genome Sequencing of Bacteria Isolated From the Coffee Berry Borer (Hypothenemus hampei), With Emphasis on the Role of Caffeine Degradation. Front. Microbiol. 2021, 12, 644768. [Google Scholar] [CrossRef] [PubMed]
- Thathola, P.; Agnihotri, V.; Pandey, A. Microbial Degradation of Caffeine Using Himalayan Psychrotolerant Pseudomonas sp.GBPI_Hb5 (MCC 3295). Curr. Microbiol. 2021, 78, 3924–3935. [Google Scholar] [CrossRef]
- Mazzafera, P.; Olsson, O.; Sandberg, G. Degradation of Caffeine and Related Methylxanthines by Serratia Marcescens Isolated from Soil under Coffee Cultivation. Microb. Ecol. 1996, 31, 199–207. [Google Scholar] [CrossRef]
- Dash, S.S.; Gummadi, S.N. Inducible Nature of the Enzymes Involved in Catabolism of Caffeine and Related Methylxanthines. J. Basic Microbiol. 2008, 48, 227–233. [Google Scholar] [CrossRef]
- Asano, Y.; Komeda, T.; Yamada, H. Enzymes Involved in Theobromine Production from Caffeine by Pseudomonas putida No. 352. Biosci. Biotechnol. Biochem. 1994, 58, 2303–2304. [Google Scholar] [CrossRef]
- Glück, M.; Lingens, F. Heteroxanthinedemethylase, a New Enzyme in the Degradation of Caffeine by Pseudomonas putida. Appl. Micobiol. Biot. 1988, 28, 59–62. [Google Scholar]
- Dash, S.S.; Gummadi, S.N. Degradation Kinetics of Caffeine and Related Methylxanthines by Induced Cells of Pseudomonas sp. Curr. Microbiol. 2007, 55, 56–60. [Google Scholar] [CrossRef]
- Yu, C.L.; Louie, T.M.; Summers, R.; Kale, Y.; Gopishetty, S.; Subramanian, M. Two Distinct Pathways for Metabolism of Theophylline and Caffeine Are Coexpressed in Pseudomonas putida CBB5. J. Bacteriol. 2009, 191, 4624–4632. [Google Scholar] [CrossRef] [Green Version]
- Summers, R.M.; Louie, T.M.; Yu, C.-L.; Gakhar, L.; Louie, K.C.; Subramanian, M. Novel, Highly Specific N -Demethylases Enable Bacteria to Live on Caffeine and Related Purine Alkaloids. J. Bacteriol. 2012, 194, 2041–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouf, M.A.; Lomprey, R.F. Degradation of Uric Acid by Certain Aerobic Bacteria. J. Bacteriol. 1968, 96, 617–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madyastha, K.M.; Sridhar, G.R. A Novel Pathway for the Metabolism of Caffeine by a Mixed Culture Consortium. Biochem. Biophys. Res. Commun. 1998, 249, 178–181. [Google Scholar] [CrossRef]
- Madyastha, K.M.; Sridhar, G.R.; Vadiraja, B.B.; Madhavi, Y.S. Purification and Partial Characterization of Caffeine Oxidase—A Novel Enzyme from a Mixed Culture Consortium. Biochem. Biophys. Res. Commun. 1999, 263, 460–464. [Google Scholar] [CrossRef]
- Yu, C.L.; Kale, Y.; Gopishetty, S.; Louie, T.M.; Subramanian, M. A Novel Caffeine Dehydrogenase in Pseudomonas sp. Strain CBB1 Oxidizes Caffeine to Trimethyluric Acid. J. Bacteriol. 2008, 190, 772–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohapatra, B.R.; Harris, N.; Nordin, R.; Mazumder, A. Purification and Characterization of a Novel Caffeine Oxidase from Alcaligenes Species. J. Bacteriol. 2006, 125, 319–327. [Google Scholar] [CrossRef]
- Mohanty, S.K.; Yu, C.-L.; Das, S.; Louie, T.M.; Gakhar, L.; Subramanian, M. Delineation of the Caffeine C-8 Oxidation Pathway in Pseudomonas sp. Strain CBB1 via Characterization of a New Trimethyluric Acid Monooxygenase and Genes Involved in Trimethyluric Acid Metabolism. J. Bacteriol. 2012, 194, 3872–3882. [Google Scholar] [CrossRef] [Green Version]
- Beltrán, J.G.; Leask, R.L.; Brown, W.A. Activity and Stability of Caffeine Demethylases Found in Pseudomonas putida IF-3. Biochem. Eng. J. 2006, 31, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Summers, R.M.; Louie, T.M.; Yu, C.L.; Subramanian, M. Characterization of a Broad-Specificity Non-Haem Iron N-Demethylase from Pseudomonas putida CBB5 Capable of Utilizing Several Purine Alkaloids as Sole Carbon and Nitrogen Source. Microbiology 2011, 157, 583–592. [Google Scholar] [CrossRef] [Green Version]
- Perry, C.; de los Santos, E.L.C.; Alkhalaf, L.M.; Challis, G.L. Rieske Non-Heme Iron-Dependent Oxygenases Catalyse Diverse Reactions in Natural Product Biosynthesis. Nat. Prod. Rep. 2018, 35, 622–632. [Google Scholar] [CrossRef]
- Wackett, L.P. Mechanism and Applications of Rieske Non-Heme Iron Dioxygenases. Enzyme Microb. Technol. 2002, 31, 577–587. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, B.H.; Brooks, S.; Kang, S.Y.; Summers, R.M.; Song, H.K. Structural and Mechanistic Insights into Caffeine Degradation by the Bacterial N-Demethylase Complex. J. Mol. Biol. 2019, 431, 3647–3661. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.-X.; Wu, X.-H.; Wu, H.-S.; Dong, Z.-B.; Ye, J.-H.; Zheng, X.-Q.; Liang, Y.-R.; Lu, J. Different Catabolism Pathways Triggered by Various Methylxanthines in Caffeine-Tolerant Bacterium Pseudomonas putida CT25 Isolated from Tea Garden Soil. J. Microbiol. Biotechnol. 2018, 28, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Summers, R.M.; Seffernick, J.L.; Quandt, E.M.; Yu, C.L.; Barrick, J.E.; Subramanian, M.V. Caffeine Junkie: An Unprecedented Glutathione S-Transferase-Dependent Oxygenase Required for Caffeine Degradation by Pseudomonas putida CBB5. J. Bacteriol. 2013, 195, 3933–3939. [Google Scholar] [CrossRef] [Green Version]
- Retnadhas, S.; Gummadi, S.N. Identification and Characterization of Oxidoreductase Component (NdmD) of Methylxanthine Oxygenase System in Pseudomonas sp. NCIM 5235. Appl. Microbiol. Biotechnol. 2018, 102, 7913–7926. [Google Scholar] [CrossRef]
- Sun, D.; Yang, X.; Zeng, C.; Li, B.; Wang, Y.; Zhang, C.; Hu, W.; Li, X.; Gao, Z. Novel Caffeine Degradation Gene Cluster Is Mega-Plasmid Encoded in Paraburkholderia caffeinilytica CF1. Appl. Microbiol. Biotechnol. 2020, 104, 3025–3036. [Google Scholar] [CrossRef] [PubMed]
- Brand, D.; Pandey, A.; Roussos, S.; Soccol, C.R. Biological Detoxification of Coffee Husk by Filamentous Fungi Using a Solid State Fermentation System. Enzyme Microb. Technol. 2000, 27, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Schwimmer, S.; Kurtzman, R.H.; Heftmann, E. Caffeine Metabolism by Penicillium roqueforti. Arch. Biochem. Biophys. 1971, 147, 109–113. [Google Scholar] [CrossRef]
- Nanjundaiah, S.; Mutturi, S.; Bhatt, P. Modeling of Caffeine Degradation Kinetics during Cultivation of Fusarium solani Using Sucrose as Co-Substrate. Biochem. Eng. J. 2017, 125, 73–80. [Google Scholar] [CrossRef]
- Nayak, V.; Pai, P.V.; Pai, A.; Pai, S.; Sushma, Y.D.; Rao, C.V. A Comparative Study of Caffeine Degradation by Four Different Fungi. Bioremediat. J. 2013, 17, 79–85. [Google Scholar] [CrossRef]
- Roussos, S.; de los Angeles Aquiáhuatl, M.; del Refugio Trejo-Hernández, M.; Gaime Perraud, I.; Favela, E.; Ramakrishna, M.; Raimbault, M.; Viniegra-González, G. Biotechnological Management of Coffee Pulp—Isolation, Screening, Characterization, Selection of Caffeine-Degrading Fungi and Natural Microflora Present in Coffee Pulp and Husk. Appl. Micobiol. Biotechnol. 1995, 42, 756–762. [Google Scholar] [CrossRef]
- Hakil, M.; Denis, S.; Viniegra-González, G.; Augur, C. Degradation and Product Analysis of Caffeine and Related Dimethylxanthines by Filamentous Fungi. Enzyme Microb. Technol. 1998, 22, 355–359. [Google Scholar] [CrossRef]
- Denis, S.; Augur, C.; Marin, B.; Roussos, S. A New HPLC Analytical Method to Study Fungal Caffeine Metabolism. Biotechnol. Tech. 1998, 12, 359–362. [Google Scholar] [CrossRef]
- Tagliari, C.V.; Sanson, R.K.; Zanette, A.; Franco, T.T.; Soccol, C.R. Caffeine Degradation by Rhizopus Delemar in Packed Bed Column Bioreactor Using Coffee Husk as Substrate. Braz. J. Microbiol. 2003, 34, 102–104. [Google Scholar] [CrossRef] [Green Version]
- De Mejia, E.G.; Ramirez-Mares, M.V. Impact of Caffeine and Coffee on Our Health. Trends Endocrinol. Metab. 2014, 25, 489–492. [Google Scholar] [CrossRef]
- Jahrami, H.; Al-Mutarid, M.; Penson, P.E.; Al-Islam Faris, M.; Saif, Z.; Hammad, L. Intake of Caffeine and Its Association with Physical and Mental Health Status among University Students in Bahrain. Foods 2020, 9, 473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, N.K.; Honig, S.; Cronstein, B.N.; Pillinger, M.H. The Effects of Caffeine on Bone Mineral Density and Fracture Risk. Osteoporos. Int. 2022, 33, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Salari-Moghaddam, A.; Saneei, P.; Askari, M.; Larijani, B.; Azadbakht, L.; Esmaillzadeh, A. Maternal Caffeine Consumption during Pregnancy and Risk of Low Birth Weight: A Dose–Response Meta-Analysis of Cohort Studies. Crit. Rev. Food Sci. 2021, 63, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Shilo, L.; Sabbah, H.; Hadari, R.; Kovatz, S.; Weinberg, U.; Dolev, S.; Dagan, Y.; Shenkman, L. The Effects of Coffee Consumption on Sleep and Melatonin Secretion. Sleep Med. 2002, 3, 271–273. [Google Scholar] [CrossRef]
- Roehrs, T.; Roth, T. Caffeine: Sleep and Daytime Sleepiness. Sleep Med. Rev. 2008, 12, 153–162. [Google Scholar] [CrossRef]
- Daly, J.W.; Fredholm, B.B. Caffeine-an Atypical Drug of Dependence. Drug Alcohol Depend. 1998, 51, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, N.; Ueki, H. Clinical Importance of Caffeine Dependence and Abuse. Psychiat. Clin. Neurosci. 2007, 61, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Schuh, K.J.; Griffiths, R.R. Caffeine Reinforcement: The Role of Withdrawal. Psychopharmacology 1997, 130, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Nehlig, A. Are We Dependent upon Coffee and Caffeine? A Review on Human and Animal Data. Neurosci. Biobehav. Rev. 1999, 23, 563–576. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, M.L.; dos Santos, D.Y.A.C.; Chow, F.; Pompêo, M.L.M. Caffeine as a Contaminant of Periphyton: Ecological Changes and Impacts on Primary Producers. Ecotoxicology 2021, 30, 599–609. [Google Scholar] [CrossRef]
- Vieira, L.R.; Soares, A.M.V.M.; Freitas, R. Caffeine as a Contaminant of Concern: A Review on Concentrations and Impacts in Marine Coastal Systems. Chemosphere 2022, 286, 131675. [Google Scholar] [CrossRef]
- Dafouz, R.; Cáceres, N.; Rodríguez-Gil, J.L.; Mastroianni, N.; López de Alda, M.; Barceló, D.; de Miguel, Á.G.; Valcárcel, Y. Does the Presence of Caffeine in the Marine Environment Represent an Environmental Risk? A Regional and Global Study. Sci. Total Environ. 2018, 615, 632–642. [Google Scholar] [CrossRef]
- Bachmann, S.A.L.; Calvete, T.; Féris, L.A. Caffeine Removal from Aqueous Media by Adsorption: An Overview of Adsorbents Evolution and the Kinetic, Equilibrium and Thermodynamic Studies. Sci. Total Environ. 2021, 767, 144229. [Google Scholar] [CrossRef]
- Romano, R.; De Luca, L.; Basile, G.; Nitride, C.; Pizzolongo, F.; Masi, P. The Use of Carbon Dioxide as a Green Approach to Recover Bioactive Compounds from Spent Coffee Grounds. Foods 2023, 12, 1958. [Google Scholar] [CrossRef]
- Kobori, K.; Maruta, Y.; Mineo, S.; Shigematsu, T.; Hirayama, M. Polyphenol-Retaining Decaffeinated Cocoa Powder Obtained by Supercritical Carbon Dioxide Extraction and Its Antioxidant Activity. Foods 2013, 2, 462–477. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Niu, G.; Liu, H. Microwave-Assisted Extraction of Tea Polyphenols and Tea Caffeine from Green Tea Leaves. Chem. Eng. Process. 2003, 42, 129–133. [Google Scholar] [CrossRef]
- Lou, Z.; Er, C.; Li, J.; Wang, H.; Zhu, S.; Sun, J. Removal of Caffeine from Green Tea by Microwave-Enhanced Vacuum Ice Water Extraction. Anal. Chim. Acta 2012, 716, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Serna-Jiménez, J.A.; Torres-Valenzuela, L.S.; Sanín Villarreal, A.; Roldan, C.; Martín, M.A.; Siles, J.A.; Chica, A.F. Advanced Extraction of Caffeine and Polyphenols from Coffee Pulp: Comparison of Conventional and Ultrasound-Assisted Methods. LWT 2023, 177, 114571. [Google Scholar] [CrossRef]
- Sheng, C.; Nnanna, A.G.A.; Liu, Y.; Vargo, J.D. Removal of Trace Pharmaceuticals from Water Using Coagulation and Powdered Activated Carbon as Pretreatment to Ultrafiltration Membrane System. Sci. Total Environ. 2016, 550, 1075–1083. [Google Scholar] [CrossRef]
- Kwon, Y.; Lee, D.G. Removal of Contaminants of Emerging Concern (CECs) Using a Membrane Bioreactor (MBR): A Short Review. Glob. NEST J. 2019, 21, 337–346. [Google Scholar]
- Yu, Z.; Deng, H.; Qu, H.; Zhang, B.; Lei, G.; Chen, J.; Feng, X.; Wu, D.; Huang, Y.; Ji, Z. Penicillium simplicissimum Possessing High Potential to Develop Decaffeinated Qingzhuan Tea. LWT 2022, 165, 113606. [Google Scholar] [CrossRef]
- Jiang, Y.; Lu, Y.; Huang, Y.; Chen, S.; Ji, Z. Bacillus amyloliquefaciens HZ-12 Heterologously Expressing NdmABCDE with Higher Ability of Caffeine Degradation. LWT 2019, 109, 387–394. [Google Scholar] [CrossRef]
- Gokulakrishnan, S.; Chandraraj, K.; Gummadi, S.N. Microbial and Enzymatic Methods for the Removal of Caffeine. Enzyme Microb. Technol. 2005, 37, 225–232. [Google Scholar] [CrossRef]
- Arya, S.S.; Venkatram, R.; More, P.R.; Vijayan, P. The Wastes of Coffee Bean Processing for Utilization in Food: A Review. J. Food Sci. Technol. 2022, 59, 429–444. [Google Scholar] [CrossRef]
- Peña-Lucio, E.M.; Londoño-Hernández, L.; Ascacio-Valdes, J.A.; Chavéz-González, M.L.; Bankole, O.E.; Aguilar, C.N. Use of Coffee Pulp and Sorghum Mixtures in the Production of N-Demethylases by Solid-State Fermentation. Bioresour. Technol. 2020, 305, 123112. [Google Scholar] [CrossRef]
- Mazzafera, P. Degradation of Caffeine by Microorganisms and Potential Use of Decaffeinated Coffee Husk and Pulp in Animal Feeding. Sci. Agric. 2002, 59, 815–821. [Google Scholar] [CrossRef] [Green Version]
- Carrasco-Cabrera, C.P.; Bell, T.L.; Kertesz, M.A. Caffeine Metabolism during Cultivation of Oyster Mushroom (Pleurotus ostreatus) with Spent Coffee Grounds. Appl. Microbiol. Biotechnol. 2019, 103, 5831–5841. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Pandey, A.; Mohan, R.; Soccol, C.R. Use of Various Coffee Industry Residues for the Cultivation of Pleurotus Ostreatus in Solid State Fermentation. Acta Biotechnol. 2000, 20, 41–52. [Google Scholar] [CrossRef]
- Leifa, F.; Pandey, A.; Soccol, C.R. Production of Flammulina velutipes on Coffee Husk and Coffee Spent-Ground. Braz. Arch. Biol. Technol. 2001, 44, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; He, B.; Wang, J.; Liu, J.; Hu, X. Risks of Caffeine Residues in the Environment: Necessity for a Targeted Ecopharmacovigilance Program. Chemosphere 2020, 243, 125343. [Google Scholar] [CrossRef]
- Ogunseitan, O.A. Removal of Caffeine in Sewage by Pseudomonas putida: Implications for Water Pollution Index. World J. Microbiol. Biotechnol. 1996, 12, 251–256. [Google Scholar] [CrossRef]
- Shanmugam, M.K. Self-Directing Optimization for Enhanced Caffeine Degradation in Synthetic Coffee Wastewater Using Induced Cells of Pseudomonas sp.: Bioreactor Studies. J. Water Process Eng. 2021, 44, 102341. [Google Scholar] [CrossRef]
- Singh, N.; Shreshtha, A.K.; Thakur, M.S.; Patra, S. Xanthine Scaffold: Scope and Potential in Drug Development. Heliyon 2018, 4, e00829. [Google Scholar] [CrossRef] [Green Version]
- Janitschke, D.; Lauer, A.A.; Bachmann, C.M.; Grimm, H.S.; Hartmann, T.; Grimm, M.O.W. Methylxanthines and Neurodegenerative Diseases: An Update. Nutrients 2021, 13, 803. [Google Scholar] [CrossRef]
- Cyr, M.C.; Beauchesne, M.F.; Lemière, C.; Blais, L. Effect of Theophylline on the Rate of Moderate to Severe Exacerbations among Patients with Chronic Obstructive Pulmonary Disease. Br. J. Clin. Pharmacol. 2008, 65, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Barnes, P.J. Theophylline. Am. J. Respir. Crit. Care Med. 2013, 188, 901–906. [Google Scholar] [CrossRef]
- Usmani, O.S.; Belvisi, M.G.; Patel, H.J.; Crispino, N.; Birrell, M.A.; Korbonits, M.; Korbonits, D.; Barnes, P.J. Theobromine Inhibits Sensory Nerve Activation and Cough. FASEB J. 2005, 19, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Guerreiro, S.; Toulorge, D.; Hirsch, E.; Marien, M.; Sokoloff, P.; Michel, P.P. Paraxanthine, the Primary Metabolite of Caffeine, Provides Protection against Dopaminergic Cell Death via Stimulation of Ryanodine Receptor Channels. Mol. Pharmacol. 2008, 74, 980–989. [Google Scholar] [CrossRef] [Green Version]
- Trier, K.; Munk Ribel-Madsen, S.; Cui, D.; Brøgger Christensen, S. Systemic 7-Methylxanthine in Retarding Axial Eye Growth and Myopia Progression: A 36-Month Pilot Study. J. Ocul. Biol. Dis. Inform. 2008, 1, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Mock, M.B.; Mills, S.B.; Cyrus, A.; Campo, H.; Dreischarf, T.; Strock, S.; Summers, R.M. Biocatalytic Production and Purification of the High-Value Biochemical Paraxanthine. Biotechnol. Bioprocess Eng. 2022, 27, 640–651. [Google Scholar] [CrossRef]
- Mock, M.B.; Cyrus, A.; Summers, R.M. Biocatalytic Production of 7-methylxanthine by a Caffeine-degrading Escherichia coli Strain. Biotechnol. Bioeng. 2022, 119, 3326–3331. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Ma, C.; Xia, T.; Li, X.; Zheng, C.; Wu, T.; Liu, X. Isolation, Characterization and Application of Theophylline-Degrading Aspergillus Fungi. Microb. Cell Fact. 2020, 19, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Ma, C.; Ren, X.; Xia, T.; Li, X.; Wu, Y. Production of Theophylline via Aerobic Fermentation of Pu-Erh Tea Using Tea-Derived Fungi. BMC Microbiol. 2019, 19, 261. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Substrate | Product | Km (μM) | kcat (min−1) | kcat/Km (min−1·μΜ−1) |
|---|---|---|---|---|---|
| NdmA-His6 | Caffeine | Theobromine | 37 ± 8 | 190 ± 10 | 5.1 ± 1.2 |
| Theophylline | 3-methylxanthine | 9.1 ± 1.7 | 83 ± 1.7 | 9.1 ± 1.7 | |
| Paraxanthine | 7-methylxanthine | 53 ± 20 | 130 ± 10 | 2.5 ± 0.8 | |
| Theobromine | – | >500 | NA | NA | |
| 1-methylxanthine | Xanthine | 270 ± 50 | 16 ± 1 | 0.06 ± 0.01 | |
| 3-methylxanthine | – | >500 | NA | NA | |
| 7-methylxanthine | – | >500 | NA | NA | |
| NdmB-His6 | Caffeine | Paraxanthine | 42 ± 9 | 0.23 ± 0.03 | 0.006 ± 0.001 |
| Theophylline | Methylxanthine | 170 ± 50 | 0.27 ± 0.03 | 0.016 ± 0.005 | |
| Paraxanthine | – | >500 | NA | NA | |
| Theobromine | 7-methylxanthine | 25 ± 5 | 46 ± 1.9 | 1.8 ± 0.4 | |
| 1-methylxanthine | – | >500 | NA | NA | |
| 3-methylxanthine | Xanthine | 22 ± 5 | 32 ± 1.5 | 1.4 ± 0.3 | |
| 7-methylxanthine | – | >500 | NA | NA |
| Gene | Homologous Protein | % Identity | Organism/NCBI Accession Number | Proposed Function |
|---|---|---|---|---|
| orf1 | S-formylglutathione hydrolase | 75 | Pseudomonas fluorescens WP_017340027.1 | Involved in formaldehyde metabolism |
| NdmE | Glutathione S-transferase | 86 | Pseudomonas fluorescens WP_017340028.1 | Structural protein/chaperone |
| NdmD | Oxidoreductase | 81 | Pseudomonas fluorescens WP_080995163.1 | Reductase component of methylxanthine demethylases |
| NdmC | Aromatic-ring-hydroxylating dioxygenase subunit alpha | 88 | Pseudomonas fluorescens WP_017340030.1 | Methyxanthine N7-demethylation |
| orf5 | Hypothetical protein | 92 | Pseudomonas fluorescens WP_017340031.1 | Unknown |
| orf6 | Purine permease | 88 | Pseudomonas fluorescens WP_017340032.1 | Transport of methylxanthines |
| NdmB | Aromatic-ring-hydroxylating dioxygenase subunit alpha | 91 | Pseudomonas fluorescens WP_017340033.1 | Methyxanthine N3-demethylation |
| orf8 | GntR family transcriptional regulator | 91 | Pseudomonas fluorescens WP_017340034.1 | Repressor protein of methylxanthine operon |
| orf9 | Membrane protein | 83 | Pseudomonas fluorescens WP_017340035.1 | Unknown |
| NdmA | Aromatic-ring-hydroxylating dioxygenase subunit alpha | 92 | Pseudomonas fluorescens WP_031319057.1 | Methyxanthine N1-demethylation |
| orf11 | VOC family protein | 83 | Pseudomonas fluorescens WP_057008287.1 | Metal dependent vicinal oxygen chelating enzyme |
| orf12 | S-(hydroxymethyl) glutathione dehydrogenase | 93 | Pseudomonas fluorescens WP_046819641.1 | Involved in formaldehyde metabolism |
| Enzyme | Substrate | Km (μM) | kcat (min−1) | kcat/Km (min−1·μΜ−1) |
|---|---|---|---|---|
| TmuM-His6 | Trimethyluric | 10.2 ± 2.2 | 448.9 ± 21.7 | 44.1 ± 2.1 |
| 1,3-dimethyluric acid | 126.5 ± 29.3 | 185.0 ± 16.4 | 1.5 ± 0.1 | |
| 3,7-dimethyluric acid | 1.3 ± 0.6 | 118.2 ± 8.4 | 89.4 ± 6.3 | |
| 1-methyluric acid | 1.2 ± 0.5 | 29.1 ± 2.4 | 24.5 ± 2.0 | |
| Uric acid | NA | – | NA |
| Gene | Homologous Protein | % Identity | NCBI Accession Number | Proposed Function |
|---|---|---|---|---|
| tmuH | Hydroxyisourate hydrolase | 50 | YP_002976942 | TM-HIU hydrolase |
| tmuD | OHCU decarboxylase | 39 | YP_003695277 | TM-OHCU decarboxylase |
| tmuM | FAD-binding monooxygenase | 38 | YP_003741647 | TMU monooxygenase |
| cdhA | Xanthine dehydrogenase Molybdopterin-binding protein | 49 | YP_003395893 | Cdh molybdopterin-binding subunit |
| cdhB | Alcohol dehydrogenase medium subunit | 39 | ADV16272 | Cdh FAD-binding subunit |
| cdhC | Aldehyde oxidase small subunit | 59 | YP_002521823 | Cdh [2FE-2S]-binding subunit |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Z.; Wei, J.; Hu, Y.; Pi, D.; Jiang, M.; Lang, T. Caffeine Synthesis and Its Mechanism and Application by Microbial Degradation, A Review. Foods 2023, 12, 2721. https://doi.org/10.3390/foods12142721
Lin Z, Wei J, Hu Y, Pi D, Jiang M, Lang T. Caffeine Synthesis and Its Mechanism and Application by Microbial Degradation, A Review. Foods. 2023; 12(14):2721. https://doi.org/10.3390/foods12142721
Chicago/Turabian StyleLin, Zhipeng, Jian Wei, Yongqiang Hu, Dujuan Pi, Mingguo Jiang, and Tao Lang. 2023. "Caffeine Synthesis and Its Mechanism and Application by Microbial Degradation, A Review" Foods 12, no. 14: 2721. https://doi.org/10.3390/foods12142721