
Genome-Wide Investigation and Functional Analysis Reveal That CsKCS3 and CsKCS18 Are Required for Tea Cuticle Wax Formation
 ,
,
Abstract
:1. Introduction
2. Results
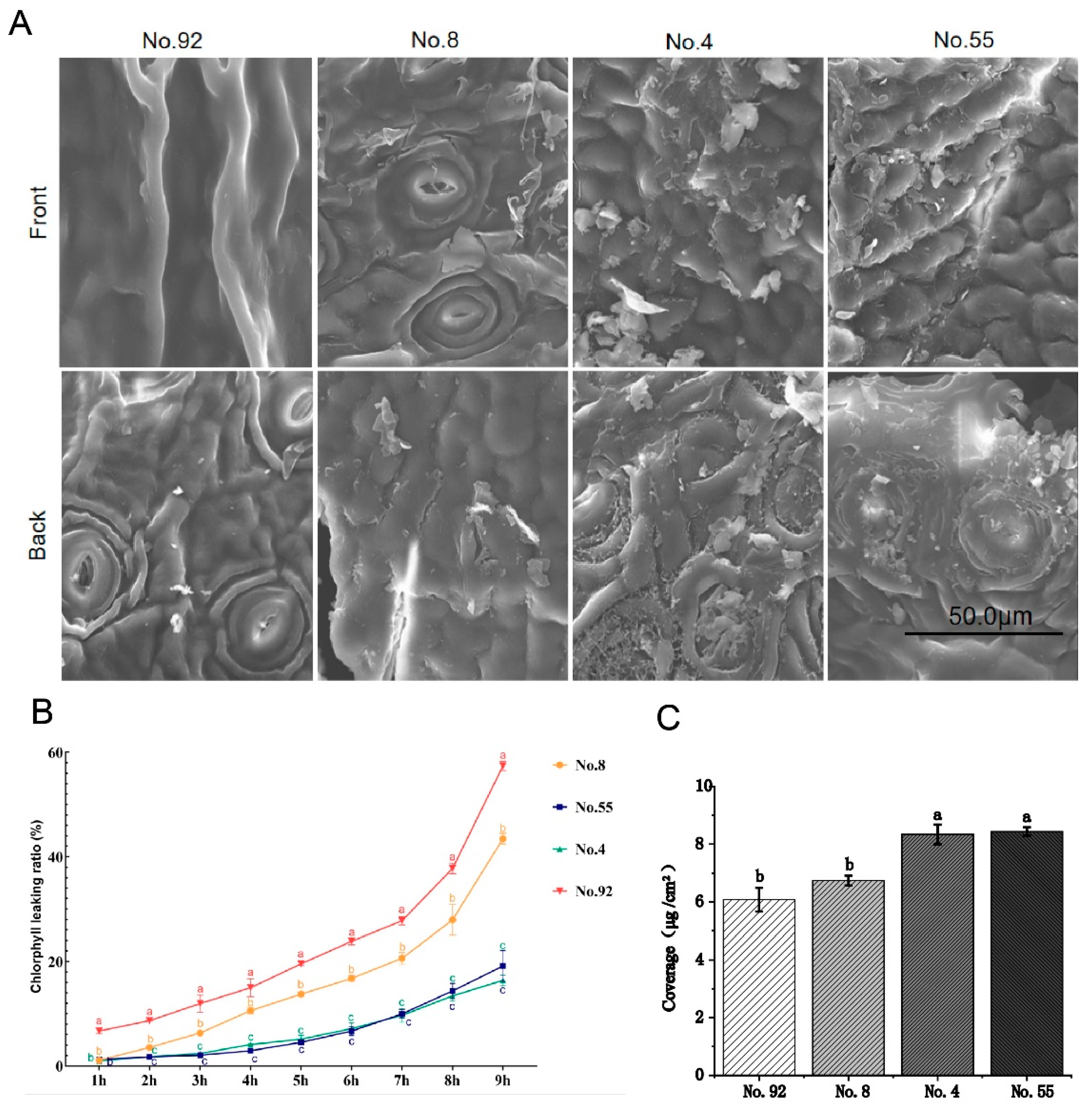
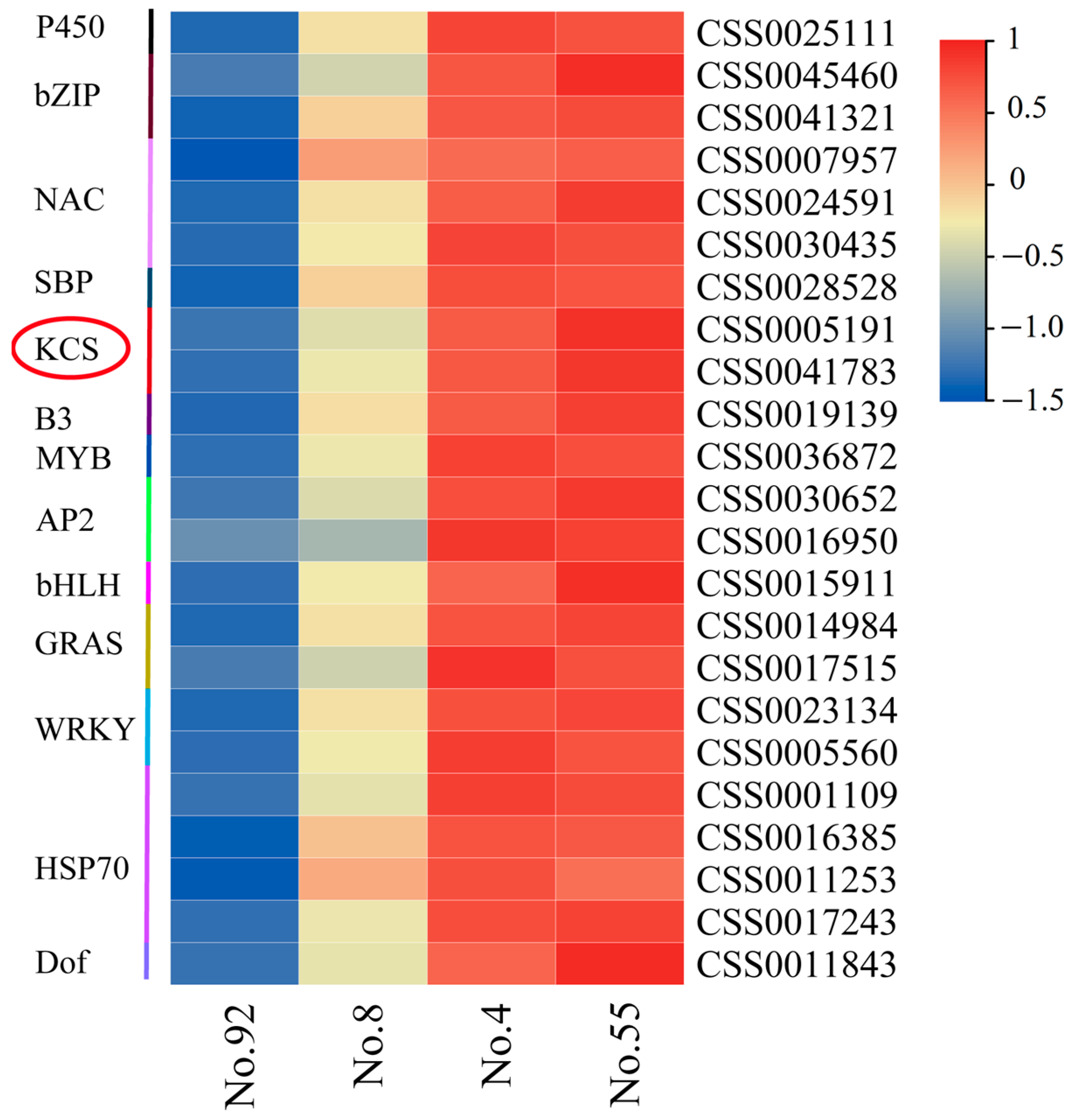
2.1. Correlation Analysis between Cuticle Wax and Expression Profile
2.2. Identification, Physicochemical Properties, and Subcellular Localization Prediction Analysis of CsKCS Genes
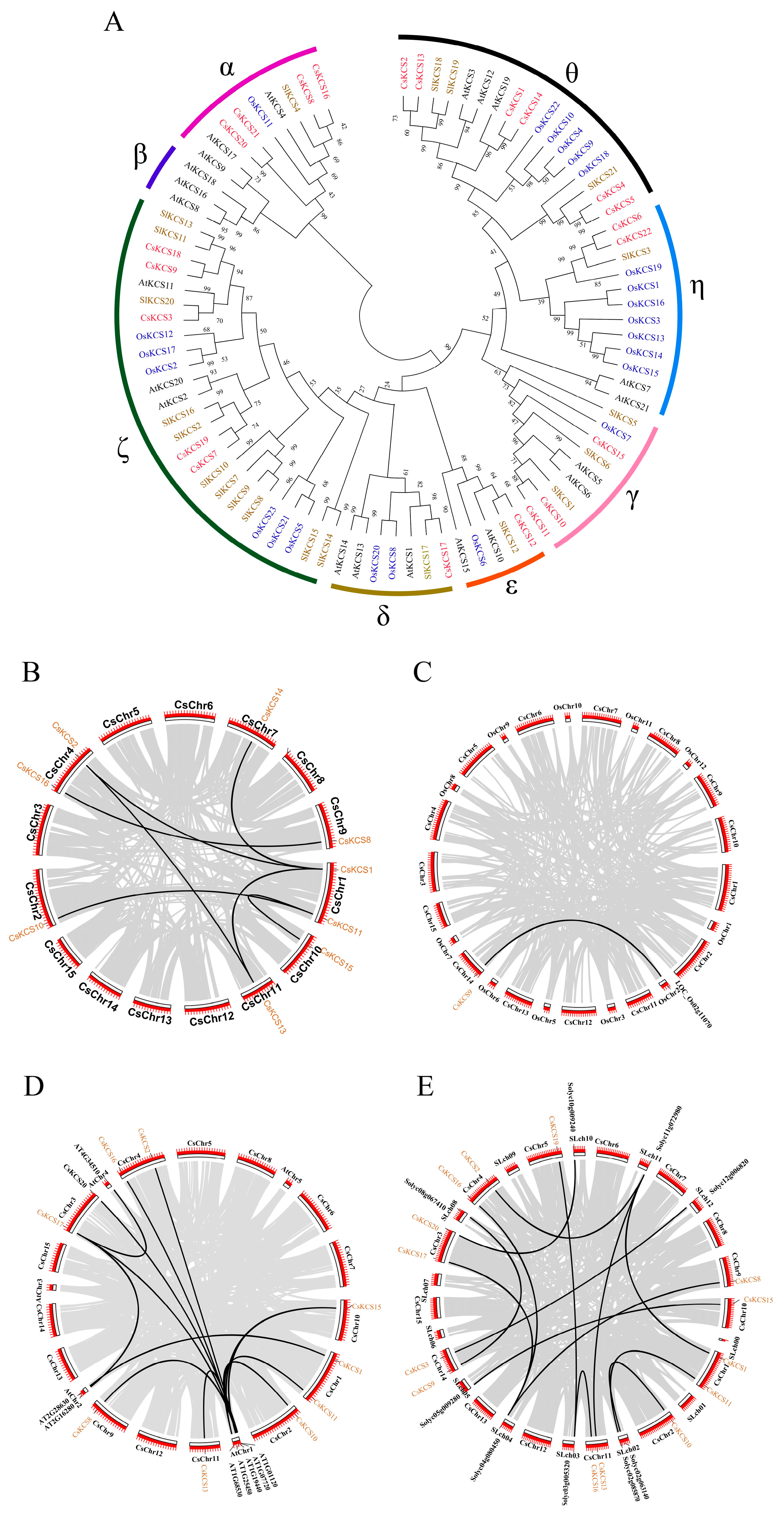
2.3. Phylogenetic and Collinearity Analysis of KCS Genes
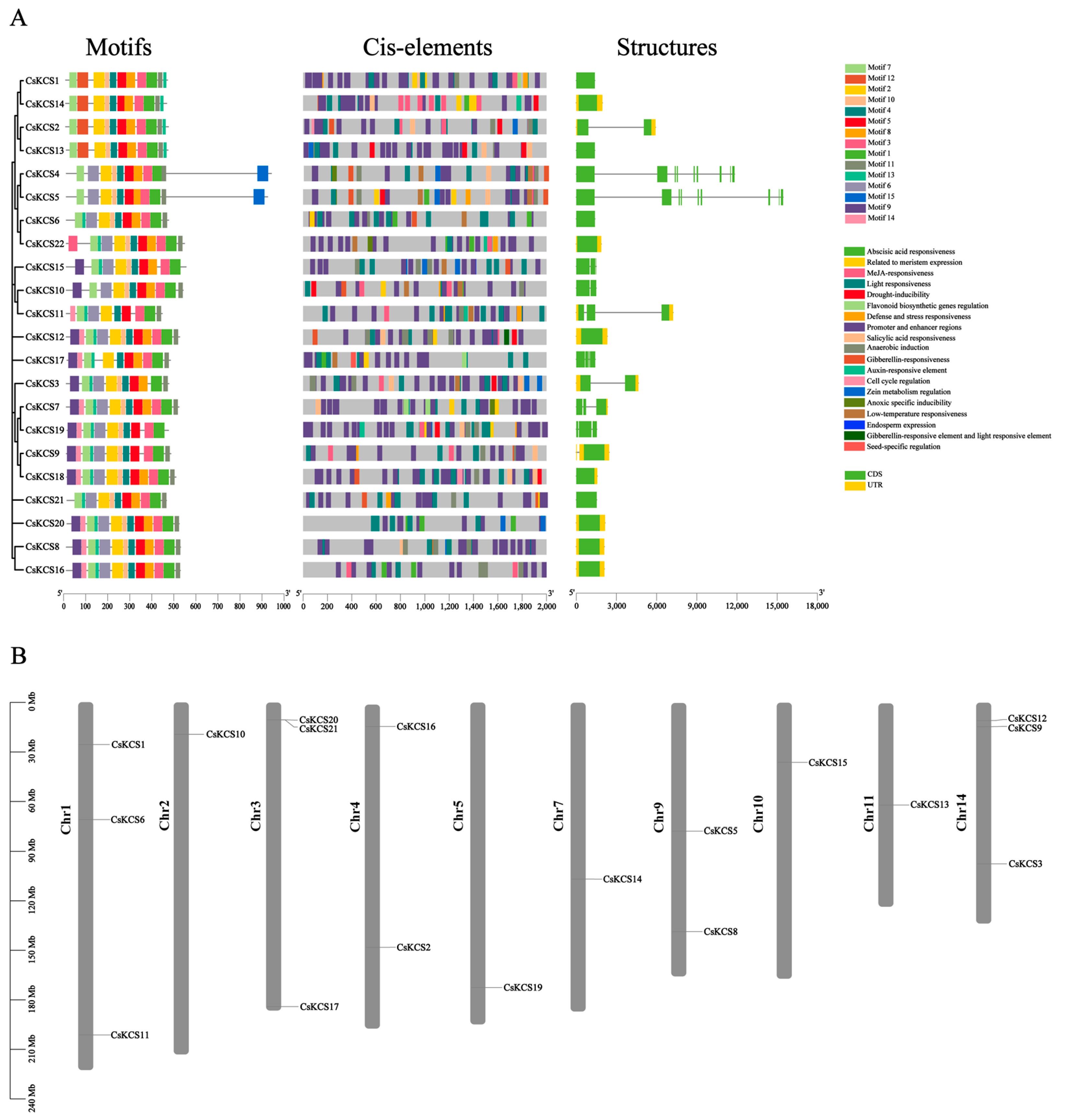
2.4. Analysis of Conservative Protein Motif, Gene Structure, and Chromosomal Location of CsKCS Genes
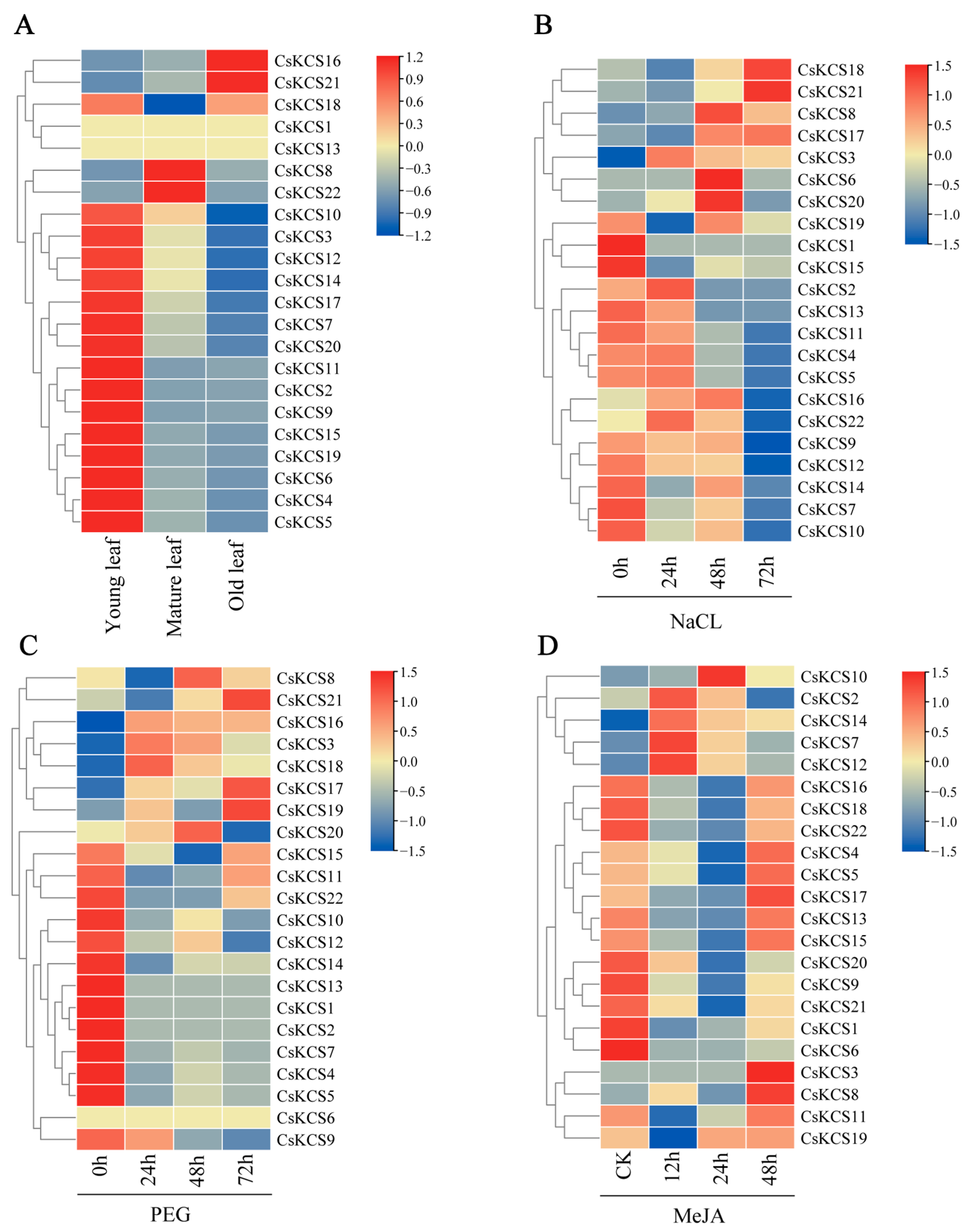
2.5. Expression Analysis of CsKCS Genes under Different Stresses
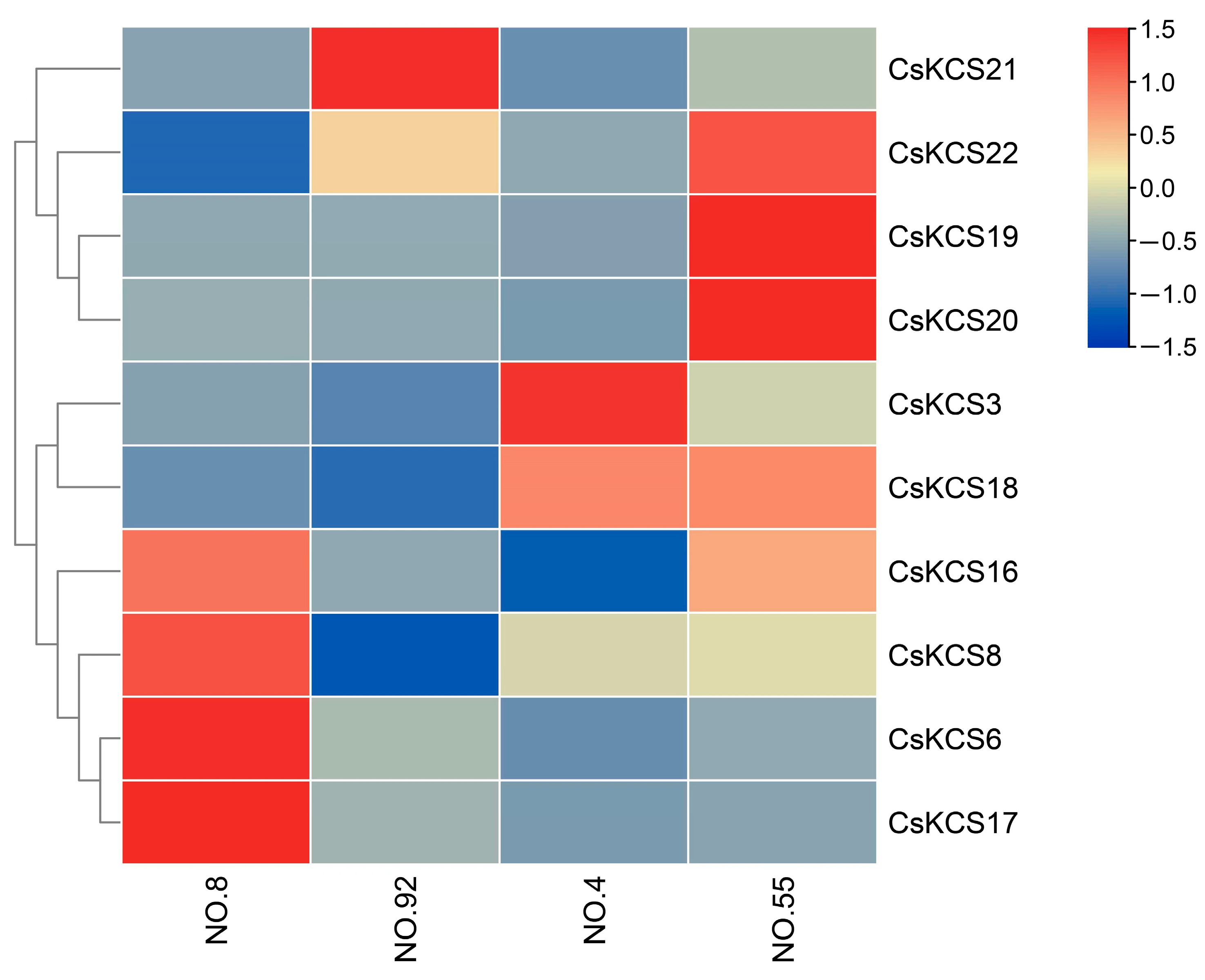
2.6. Expression Analysis of CsKCSs in Tea Plants with Different Cuticular Wax Content
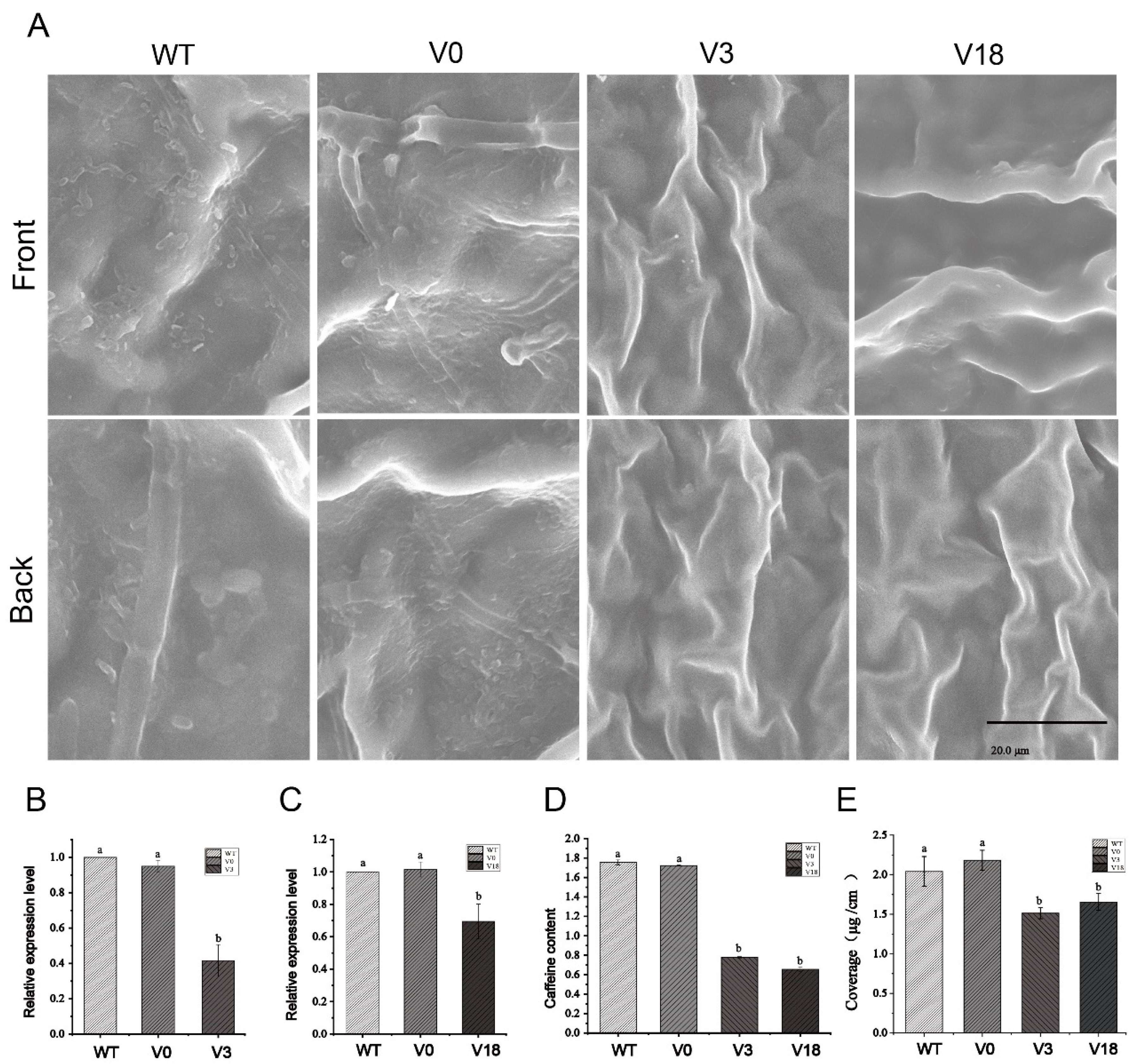
2.7. VIGS of CsKCS3 and CsKCS18 in Tea Plants
3. Discussion
4. Material and Methods
4.1. Plant Material
4.2. Scanning Electron Microscopy for Observing Tea Leaf Cuticular Wax
4.3. Determination of Chlorophyll Extraction Rate
4.4. Differentially Expressed Gene Analysis by Transcriptome Sequencing
4.5. Extraction and Measurement of Cuticle Wax of Tea Leaves
4.6. Bioinformatics Analysis of the CsKCS Gene Family
4.7. RNA Extraction and qRT-PCR Analysis
4.8. Virus-Induced Gene Silencing of CsKCS3 and CsKCS18 in Tea Plants
4.9. Determination of Caffeine Content
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Koch, K.; Hartmann, K.D.; Schreiber, L.; Barthlott, W.; Neinhuis, C. Influences of air humidity during the cultivation of plants on wax chemical composition, morphology and leaf surface wettability. Environ. Exp. Bot. 2006, 56, 1–9. [Google Scholar] [CrossRef]
- Isaacson, T.; Kosma, D.K.; Matas, A.J.; Buda, G.J.; He, Y.; Yu, B.; Pravitasari, A.; Batteas, J.D.; Stark, R.E.; Jenks, M.A.; et al. Cutin deficiency in the tomato fruit cuticle consistently affects resistance to microbial infection and biomechanical properties, but not transpirational water loss. Plant J. 2009, 60, 363–377. [Google Scholar] [CrossRef]
- Todd, J.; Post-Beittenmiller, D.; Jaworski, J. KCS1encodes a fatty acid elongase 3-ketoacyl-CoA synthase affecting wax biosynthesis in Arabidopsis thaliana. Plant J. 1999, 17, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Sassa, T.; Kihara, A. Metabolism of Very Long-Chain Fatty Acids: Genes and Pathophysiology. Biomol. Ther. 2014, 22, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Huai, D.; Zhang, Y.; Zhang, C.; Cahoon, E.B.; Zhou, Y. Combinatorial Effects of Fatty Acid Elongase Enzymes on Nervonic Acid Production in Camelina sativa. PLoS ONE 2015, 10, e0131755. [Google Scholar] [CrossRef]
- Usher, S.; Han, L.; Haslam, R.P.; Michaelson, L.V.; Sturtevant, D.; Aziz, M.; Chapman, K.D.; Sayanova, O.; Napier, J.A. Tailoring seed oil composition in the real world: Optimising omega-3 long chain polyunsaturated fatty acid accumulation in transgenic Camelina sativa. Sci. Rep. 2017, 7, 6570. [Google Scholar] [CrossRef]
- Sagar, M.; Pandey, N.; Qamar, N.; Singh, B.; Shukla, A. Domain analysis of 3 Keto Acyl-CoA synthase for structural variations in Vitis vinifera and Oryza brachyantha using comparative modelling. Interdiscip. Sci. Comput. Life Sci. 2015, 7, 7–20. [Google Scholar] [CrossRef]
- Joubès, J.; Raffaele, S.; Bourdenx, B.; Garcia, C.; Laroche-Traineau, J.; Moreau, P.; Domergue, F.; Lessire, R. The VLCFA elongase gene family in Arabidopsis thaliana: Phylogenetic analysis, 3D modelling and expression profiling. Plant Mol. Biol. 2008, 67, 547–566. [Google Scholar] [CrossRef]
- Guo, H.-S.; Zhang, Y.-M.; Sun, X.-Q.; Li, M.-M.; Hang, Y.-Y.; Xue, J.-Y. Evolution of the KCS gene family in plants: The history of gene duplication, sub/neofunctionalization and redundancy. Mol. Genet. Genom. 2016, 291, 739–752. [Google Scholar] [CrossRef]
- Huai, D.; Xue, X.; Li, Y.; Wang, P.; Li, J.; Yan, L.; Chen, Y.; Wang, X.; Liu, N.; Kang, Y.; et al. Genome-Wide Identification of Peanut KCS Genes Reveals That AhKCS1 and AhKCS28 Are Involved in Regulating VLCFA Contents in Seeds. Front. Plant Sci. 2020, 11, 406. [Google Scholar] [CrossRef]
- Fan, Y.; Yuan, C.; Jin, Y.; Hu, G.-R.; Li, F.-L. Characterization of 3-ketoacyl-coA synthase in a nervonic acid producing oleaginous microalgae Mychonastes afer. Algal Res. 2018, 31, 225–231. [Google Scholar] [CrossRef]
- James, D.W.; Lim, E.; Keller, J.; Plooy, I.; Ralston, E.; Dooner, H.K. Directed tagging of the Arabidopsis FATTY ACID ELONGATION1 (FAE1) gene with the maize transposon activator. Plant Cell 1995, 7, 309–319. [Google Scholar] [CrossRef]
- Denic, V.; Weissman, J.S. A Molecular Caliper Mechanism for Determining Very Long-Chain Fatty Acid Length. Cell 2007, 130, 663–677. [Google Scholar] [CrossRef]
- Sun, X.; Pang, H.; Li, M.; Peng, B.; Guo, H.; Yan, Q.; Hang, Y. Evolutionary Pattern of the FAE1 Gene in Brassicaceae and Its Correlation with the Erucic Acid Trait. PLoS ONE 2013, 8, e83535. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Lang, C.; Wang, F.; Wu, X.; Liu, R.; Zheng, T.; Zhang, D.; Chen, J.; Wu, G. Depressed expression of FAE1 and FAD2 genes modifies fatty acid profiles and storage compounds accumulation in Brassica napus seeds. Plant Sci. 2017, 263, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Ozseyhan, M.E.; Kang, J.; Mu, X.; Lu, C. Mutagenesis of the FAE1 genes significantly changes fatty acid composition in seeds of Camelina sativa. Plant Physiol. Biochem. 2018, 123, 1–7. [Google Scholar] [CrossRef]
- Guo, Y.; Mietkiewska, E.; Francis, T.; Katavic, V.; Brost, J.M.; Giblin, M.; Barton, D.L.; Taylor, D.C. Increase in nervonic acid content in transformed yeast and transgenic plants by introduction of a Lunaria annua L. 3-ketoacyl-CoA synthase (KCS) gene. Plant Mol. Biol. 2009, 69, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Blacklock, B.J.; Jaworski, J.G. Substrate specificity of Arabidopsis 3-ketoacyl-CoA synthases. Biochem. Biophys. Res. Commun. 2006, 346, 583–590. [Google Scholar] [CrossRef]
- Nath, U.K.; Wilmer, J.A.; Wallington, E.J.; Becker, H.C.; Möllers, C. Increasing erucic acid content through combination of endogenous low polyunsaturated fatty acids alleles with Ld-LPAAT + Bn-fae1 transgenes in rapeseed (Brassica napus L.). Theor. Appl. Genet. 2009, 118, 765–773. [Google Scholar] [CrossRef]
- Li, X.; van Loo, E.N.; Gruber, J.; Fan, J.; Guan, R.; Frentzen, M.; Stymne, S.; Zhu, L.-H. Development of ultra-high erucic acid oil in the industrial oil crop Crambe abyssinica. Plant Biotechnol. J. 2012, 10, 862–870. [Google Scholar] [CrossRef]
- Wang, N.; Shi, L.; Tian, F.; Ning, H.; Wu, X.; Long, Y.; Meng, J. Assessment of FAE1 polymorphisms in three Brassica species using EcoTILLING and their association with differences in seed erucic acid contents. BMC Plant Biol. 2010, 10, 137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Xu, J.; Xu, X.; Wu, J.; Li, P.; Wang, B.; Fang, H. Genome-wide identification and characterization of the KCS gene family in sorghum (Sorghum bicolor (L.) Moench). PeerJ 2022, 10, e14156. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Tang, H.; Wang, B.; Yue, C.; Wang, L.; Zeng, J.; Yang, Y.; Wang, X. Integrative transcriptional and metabolic analyses provide insights into cold spell response mechanisms in young shoots of the tea plant. Tree Physiol. 2018, 38, 1655–1671. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-C.; Yao, M.-Z.; Ma, C.-L.; Jin, J.-Q.; Ma, J.-Q.; Li, C.-F.; Chen, L. Physiological changes and differential gene expression of tea plant under dehydration and rehydration conditions. Sci. Hortic. 2015, 184, 129–141. [Google Scholar] [CrossRef]
- Wang, S.; Liu, L.; Mi, X.; Zhao, S.; An, Y.; Xia, X.; Guo, R.; Wei, C. Multi-omics analysis to visualize the dynamic roles of defense genes in the response of tea plants to gray blight. Plant J. 2021, 106, 862–875. [Google Scholar] [CrossRef]
- Wang, Y.-N.; Tang, L.; Hou, Y.; Wang, P.; Yang, H.; Wei, C.-L. Differential transcriptome analysis of leaves of tea plant (Camellia sinensis) provides comprehensive insights into the defense responses to Ectropis oblique attack using RNA-Seq. Funct. Integr. Genom. 2016, 16, 383–398. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, X.; Zhang, Y.; Du, Z.; Chen, X.; Kong, X.; Sun, W.; Chen, C. Drought stress modify cuticle of tender tea leaf and mature leaf for transpiration barrier enhancement through common and distinct modes. Sci. Rep. 2020, 10, 6696. [Google Scholar] [CrossRef]
- Ladaniya, M.S. Physico–chemical, respiratory and fungicide residue changes in wax coated mandarin fruit stored at chilling temperature with intermittent warming. J. Food Sci. Technol. 2011, 48, 150–158. [Google Scholar] [CrossRef]
- Zhu, J.; Huang, K.; Cheng, D.; Zhang, C.; Li, R.; Liu, F.; Wen, H.; Tao, L.; Zhang, Y.; Li, C.; et al. Characterization of Cuticular Wax in Tea Plant and Its Modification in Response to Low Temperature. J. Agric. Food Chem. 2022, 70, 13849–13861. [Google Scholar] [CrossRef]
- Zuo, H.; Si, X.; Li, P.; Li, J.; Chen, Z.; Li, P.; Chen, C.; Liu, Z.; Zhao, J. Dynamic change of tea (Camellia sinensis) leaf cuticular wax in white tea processing for contribution to tea flavor formation. Food Res. Int. 2023, 163, 112182. [Google Scholar] [CrossRef]
- Zhao, J.; Li, P.; Xia, T.; Wan, X. Exploring plant metabolic genomics: Chemical diversity, metabolic complexity in the biosynthesis and transport of specialized metabolites with the tea plant as a model. Crit. Rev. Biotechnol. 2020, 40, 667–688. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Li, Y.; Yao, X.; Lu, L. Establishment of a Virus-Induced Gene-Silencing (VIGS) System in Tea Plant and Its Use in the Functional Analysis of CsTCS1. Int. J. Mol. Sci. 2022, 24, 392. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Joubès, J. Arabidopsis cuticular waxes: Advances in synthesis, export and regulation. Prog. Lipid Res. 2013, 52, 110–129. [Google Scholar] [CrossRef]
- Pascal, S.; Bernard, A.; Deslous, P.; Gronnier, J.; Fournier-Goss, A.; Domergue, F.; Rowland, O.; Joubès, J. Arabidopsis CER1-LIKE1 Functions in a Cuticular Very-Long-Chain Alkane-Forming Complex. Plant Physiol. 2019, 179, 415–432. [Google Scholar] [CrossRef]
- Li, J.-J.; Zhang, C.-L.; Zhang, Y.-L.; Gao, H.-N.; Wang, H.-B.; Jiang, H.; Li, Y.-Y. An apple long-chain acyl-CoA synthase, MdLACS1, enhances biotic and abiotic stress resistance in plants. Plant Physiol. Biochem. 2022, 189, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Meng, W.; Wang, Y.; Zhou, Y.; Wang, S.; Qi, F.; Wang, N.; Ma, J. Comparative Analysis of Physiological, Hormonal and Transcriptomic Responses Reveal Mechanisms of Saline-Alkali Tolerance in Autotetraploid Rice (Oryza sativa L.). Int. J. Mol. Sci. 2022, 23, 16146. [Google Scholar] [CrossRef]
- Tariq, F.; Zhao, S.; Ahmad, N.; Wang, P.; Shao, Q.; Ma, C.; Yang, X. Overexpression of β-Ketoacyl CoA Synthase 2B.1 from Chenopodium quinoa Promotes Suberin Monomers’ Production and Salt Tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 13204. [Google Scholar] [CrossRef] [PubMed]
- Franke, R.; Höfer, R.; Briesen, I.; Emsermann, M.; Efremova, N.; Yephremov, A.; Schreiber, L. The DAISY gene from Arabidopsis encodes a fatty acid elongase condensing enzyme involved in the biosynthesis of aliphatic suberin in roots and the chalaza-micropyle region of seeds. Plant J. 2009, 57, 80–95. [Google Scholar] [CrossRef]
- Huang, H.; Ayaz, A.; Zheng, M.; Yang, X.; Zaman, W.; Zhao, H.; Lü, S. Arabidopsis KCS5 and KCS6 Play Redundant Roles in Wax Synthesis. Int. J. Mol. Sci. 2022, 23, 4450. [Google Scholar] [CrossRef]
- Zhou, L.; Ni, E.; Yang, J.; Zhou, H.; Liang, H.; Li, J.; Jiang, D.; Wang, Z.; Liu, Z.; Zhuang, C. Rice OsGL1-6 Is Involved in Leaf Cuticular Wax Accumulation and Drought Resistance. PLoS ONE 2013, 8, e65139. [Google Scholar] [CrossRef]
- Bourdenx, B.; Bernard, A.; Domergue, F.; Pascal, S.; Léger, A.; Roby, D.; Pervent, M.; Vile, D.; Haslam, R.P.; Napier, J.A.; et al. Overexpression of Arabidopsis ECERIFERUM1 Promotes Wax Very-Long-Chain Alkane Biosynthesis and Influences Plant Response to Biotic and Abiotic Stresses. Plant Physiol. 2011, 156, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-Y.; Broeckling, C.D.; Blancaflor, E.B.; Sledge, M.K.; Sumner, L.W.; Wang, Z.-Y. Overexpression of WXP1, a putative Medicago truncatula AP2 domain-containing transcription factor gene, increases cuticular wax accumulation and enhances drought tolerance in transgenic alfalfa (Medicago sativa). Plant J. 2005, 42, 689–707. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Goodwin, S.M.; Boroff, V.L.; Liu, X.; Jenks, M.A. Cloning and Characterization of the WAX2 Gene of Arabidopsis Involved in Cuticle Membrane and Wax Production. Plant Cell 2003, 15, 1170–1185. [Google Scholar] [CrossRef] [PubMed]
- Batsale, M.; Bahammou, D.; Fouillen, L.; Mongrand, S.; Joubès, J.; Domergue, F. Biosynthesis and Functions of Very-Long-Chain Fatty Acids in the Responses of Plants to Abiotic and Biotic Stresses. Cells 2021, 10, 1284. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.J.; Zhang, Z.B.; Fu, Z.Y.; Xu, P.; Hu, S.B.; Li, W.Q. Significance of a β-ketoacyl-CoA synthase gene expression for wheat tolerance to adverse environments. Biol. Plant. 2010, 54, 575–578. [Google Scholar] [CrossRef]
- Li, C.; Haslam, T.M.; Kr�Ger, A.; Schneider, L.M.; Mishina, K.; Samuels, L.; Yang, H.; Kunst, L.; Schaffrath, U.; Nawrath, C.; et al. The β-Ketoacyl-CoA Synthase HvKCS1, Encoded by Cerzh, Plays a Key Role in Synthesis of Barley Leaf Wax and Germination of Barley Powdery Mildew. Plant Cell Physiol. 2018, 59, 811–827. [Google Scholar] [CrossRef]
- Rui, C.; Chen, X.; Xu, N.; Wang, J.; Zhang, H.; Li, S.; Huang, H.; Fan, Y.; Zhang, Y.; Lu, X.; et al. Identification and Structure Analysis of KCS Family Genes Suggest Their Reponding to Regulate Fiber Development in Long-Staple Cotton Under Salt-Alkaline Stress. Front. Genet. 2022, 13, 812449. [Google Scholar] [CrossRef]
- Ahmed, S.; Stepp, J.R.; Orians, C.; Griffin, T.; Matyas, C.; Robbat, A.; Cash, S.; Xue, D.; Long, C.; Unachukwu, U.; et al. Effects of Extreme Climate Events on Tea (Camellia sinensis) Functional Quality Validate Indigenous Farmer Knowledge and Sensory Preferences in Tropical China. PLoS ONE 2014, 9, e109126. [Google Scholar] [CrossRef]
- Xia, E.; Li, F.; Tong, W.; Li, P.; Wu, Q.; Zhao, H.; Ge, R.; Li, R.; Li, Y.; Zhang, Z.; et al. Tea Plant Information Archive: A comprehensive genomics and bioinformatics platform for tea plant. Plant Biotechnol. J. 2019, 17, 1938–1953. [Google Scholar] [CrossRef]
- Xia, E.; Tong, W.; Hou, Y.; An, Y.; Chen, L.; Wu, Q.; Liu, Y.; Yu, J.; Li, F.; Li, R.; et al. The Reference Genome of Tea Plant and Resequencing of 81 Diverse Accessions Provide Insights into Its Genome Evolution and Adaptation. Mol. Plant 2020, 13, 1013–1026. [Google Scholar] [CrossRef]
- Liu, M.; Ma, Z.; Wang, A.; Zheng, T.; Huang, L.; Sun, W.; Zhang, Y.; Jin, W.; Zhan, J.; Cai, Y.; et al. Genome-Wide Investigation of the Auxin Response Factor Gene Family in Tartary Buckwheat (Fagopyrum tataricum). Int. J. Mol. Sci. 2018, 19, 3526. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, D302–D305. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.-J.; Obayashi, T.; Nakai, K. Protein subcellular localization prediction with wolf psort. In Proceedings of the 4th Asia-Pacific Bioinformatics Conference, Taipei, Taiwan, 13–16 February 2006; Imperial College Press; World Scientific Publishing Co.: Taipei, Taiwan, 2005; pp. 39–48. [Google Scholar]
- Rizzacasa, B.; Morini, E.; Mango, R.; Vancheri, C.; Budassi, S.; Massaro, G.; Maletta, S.; Macrini, M.; D’Annibale, S.; Romeo, F.; et al. Comparative Ct Method Quantification (2−ΔCt Method). PLoS ONE 2019, 1. [Google Scholar] [CrossRef]
- Bispo, M.S.; Veloso, M.C.C.; Pinheiro, H.L.C.; De Oliveira, R.F.; Reis, J.O.N.; De Andrade, J.B. Simultaneous determination of caffeine, theobromine, and theophylline by high-performance liquid chromatography. J. Chromatogr. Sci. 2002, 40, 45–48. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tea Leaf Cuticle Content | Tea Germplasms |
|---|---|
| High content | No. 4, No. 9, No. 11, No. 18, No. 22, No. 25, No. 28, No. 38, No. 39, No. 43, No. 45, No. 47, No. 55, No. 62, No. 65, No. 77, No. 82, No. 91, No. 96, No. 103, No. 108 |
| Medium content | No. 1, No. 2, No. 5, No. 8, No. 10, No. 13, No. 15, No. 16, No. 17, No. 19, No. 20, No. 21, No. 23, No. 24, No. 26, No. 27, No. 29, No. 30, No. 31, No. 32, No. 34, No. 35, No. 36, No. 37, No. 40, No. 41, No. 44, No. 46, No. 48, No. 49, No. 50, No. 51, No. 52, No. 53, No. 54, No. 57, No. 58, No. 63, No. 66, No. 67, No. 72, No. 73, No. 74, No. 75, No. 76, No. 79, No. 80, No. 81,No. 86, No. 87, No. 90, No. 93, No. 97, No. 98,No. 99, No. 104 |
| Low content | No. 3, No. 6, No. 7, No. 12, No. 14, No. 33, No. 42, No. 56, No. 59, No. 60, No. 61, No. 64, No. 68, No. 69, No. 70, No. 71, No. 78, No. 83, No. 84,No. 85, No. 88, No. 89, No. 92, No. 94, No. 95, No. 100, No. 101, No. 102, No. 105, No. 106, No. 107 |
| Gene Name | Gene ID | Gene Location | Protein Properties | Subcellular Localization | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Chr.No | Start | End | Length (aa) | MW/(Kda) | Pl | Instability Index | Aliphatic Index | Hydrophilicity | |||
| CsKCS1 | CSS0003658 | Chr1 | 25505510 | 25506898 | 462 | 51.96 | 7.99 | 36.88 | 93.27 | −0.029 | cytoplasm |
| CsKCS2 | CSS0004626 | Chr4 | 146654333 | 146660247 | 466 | 52.46 | 9.03 | 38.99 | 87.4 | −0.139 | Plasma membrane |
| CsKCS3 | CSS0005191 | Chr14 | 97178676 | 97180094 | 435 | 49.23 | 9.08 | 40.46 | 90.99 | −0.127 | cytoplasm |
| CsKCS4 | CSS0010963 | Contig763 | 184894 | 196709 | 930 | 104.41 | 6.37 | 44.23 | 91.37 | −0.236 | chloroplast |
| CsKCS5 | CSS0011525 | Chr9 | 77062346 | 77077789 | 913 | 102.66 | 6.17 | 43.54 | 90.82 | −0.244 | chloroplast, cytoplasm |
| CsKCS6 | CSS0013672 | Chr1 | 70974884 | 70976278 | 464 | 52.48 | 7.95 | 42.82 | 93.66 | 0.059 | cytoplasm |
| CsKCS7 | CSS0014846 | Contig56 | 380801 | 385441 | 514 | 58.10 | 9.18 | 36.52 | 96.36 | −0.015 | cytoplasm |
| CsKCS8 | CSS0025444 | Chr9 | 138119554 | 138121704 | 518 | 57.86 | 9.09 | 37.45 | 96.12 | −0.008 | cytoplasm |
| CsKCS9 | CSS0025961 | Chr14 | 14320192 | 14321730 | 465 | 52.65 | 8.87 | 44.55 | 93.7 | −0.092 | cytoplasm |
| CsKCS10 | CSS0029062 | Chr2 | 19083909 | 19085399 | 460 | 52.34 | 9.31 | 37.38 | 97.89 | 0.018 | cytoplasm |
| CsKCS11 | CSS0030456 | Chr1 | 201268181 | 201269674 | 474 | 53.85 | 9.28 | 38.99 | 101.18 | −0.01 | cytoplasm |
| CsKCS12 | CSS0031750 | Chr14 | 10648591 | 10655818 | 543 | 61.34 | 9.13 | 42.88 | 84.36 | −0.171 | cytoplasm |
| CsKCS13 | CSS0032609 | Chr11 | 61406575 | 61407960 | 461 | 52.11 | 8.81 | 40.58 | 87.55 | −0.193 | cytoplasm |
| CsKCS14 | CSS0032944 | Chr7 | 106477234 | 106479183 | 458 | 51.65 | 8.68 | 35.39 | 96 | −0.063 | cytoplasm |
| CsKCS15 | CSS0037394 | Chr10 | 36097474 | 36099342 | 497 | 56.15 | 9.2 | 45.93 | 104.41 | 0.114 | cytoplasm |
| CsKCS16 | CSS0038600 | Chr4 | 13063749 | 13065843 | 517 | 57.68 | 9.36 | 37.8 | 95.05 | −0.046 | cytoplasm |
| CsKCS17 | CSS0038850 | Chr3 | 183897657 | 183899975 | 530 | 59.42 | 8.94 | 31.08 | 92.53 | 0.013 | cytoplasm |
| CsKCS18 | CSS0041783 | Contig18 | 419337 | 421796 | 511 | 57.64 | 9.08 | 41.02 | 92.7 | −0.031 | chloroplast, cytoplasm |
| CsKCS19 | CSS0044924 | Chr5 | 172224143 | 172226480 | 469 | 52.83 | 9.38 | 33.79 | 98.96 | 0.054 | cytoplasm |
| CsKCS20 | CSS0045193 | Chr3 | 10280927 | 10282456 | 509 | 57.22 | 9.24 | 37.75 | 92.89 | −0.087 | cytoplasm |
| CsKCS21 | CSS0046169 | Chr3 | 10298869 | 10300437 | 451 | 51.11 | 9.36 | 34.67 | 98.18 | 0.079 | cytoplasm |
| CsKCS22 | CSS0047402 | Chr1 | 71036309 | 71037995 | 536 | 60.58 | 8.23 | 41.76 | 90.19 | −0.067 | chloroplast, cytoplasm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, L.; Yao, X.; Tang, H.; Li, Y.; Jiao, Y.; He, Y.; Jiang, Y.; Tian, S.; Lu, L. Genome-Wide Investigation and Functional Analysis Reveal That CsKCS3 and CsKCS18 Are Required for Tea Cuticle Wax Formation. Foods 2023, 12, 2011. https://doi.org/10.3390/foods12102011
Mo L, Yao X, Tang H, Li Y, Jiao Y, He Y, Jiang Y, Tian S, Lu L. Genome-Wide Investigation and Functional Analysis Reveal That CsKCS3 and CsKCS18 Are Required for Tea Cuticle Wax Formation. Foods. 2023; 12(10):2011. https://doi.org/10.3390/foods12102011
Chicago/Turabian StyleMo, Lilai, Xinzhuan Yao, Hu Tang, Yan Li, Yujie Jiao, Yumei He, Yihe Jiang, Shiyu Tian, and Litang Lu. 2023. "Genome-Wide Investigation and Functional Analysis Reveal That CsKCS3 and CsKCS18 Are Required for Tea Cuticle Wax Formation" Foods 12, no. 10: 2011. https://doi.org/10.3390/foods12102011