
Oil Bodies from Chia (Salvia hispanica L.) and Camelina (Camelina sativa L.) Seeds for Innovative Food Applications: Microstructure, Composition and Physical Stability
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chia Seeds and Camelina Seeds
2.2. Sample Preparation
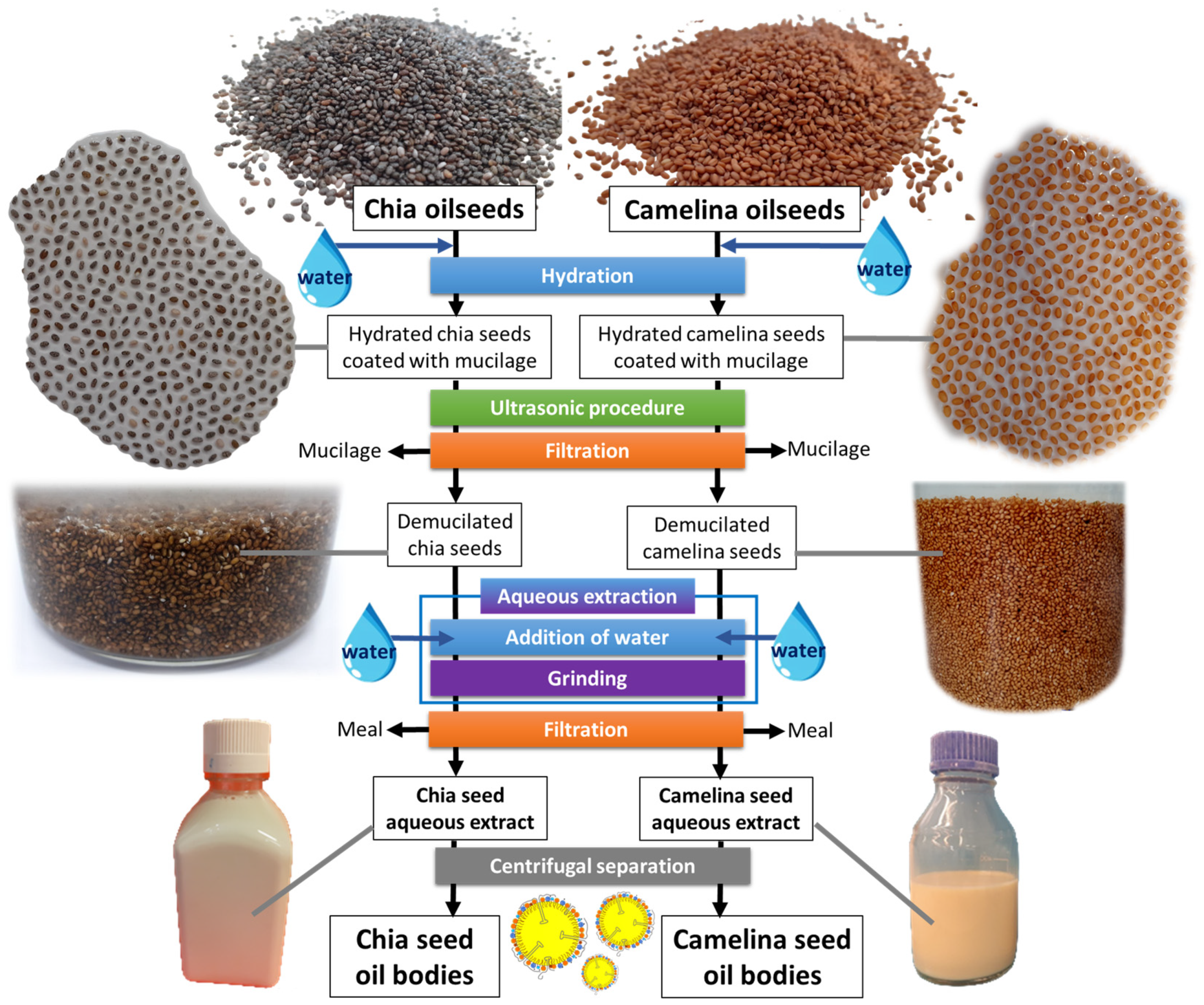
2.2.1. Plant Seed Processing: Preparation of Seed Aqueous Extracts and Oil Bodies
2.2.2. Changes in pH
2.3. Microstructure
2.4. Analysis of Proteins by Gel Electrophoresis
2.4.1. Preparation of the Samples for Gel Electrophoresis
2.4.2. Gel Electrophoresis
2.5. Particle Size Measurements
2.6. Chemical Analysis
2.6.1. Total Lipids: Extraction and Quantification
2.6.2. Fatty Acid Profiles of Chia and Camelina Seed OBs
2.6.3. Analysis of Triacylglycerols (TAGs)
Recovering of TAGs
Analysis of the Molecular Species of TAGs
Analysis of the Fatty Acid Distribution in TAGs: Production of sn-2 MAG
Analysis of Fatty Acid Composition by Gas Chromatography
2.6.4. Polar Lipid Analysis of Chia and Camelina Seed OBs
2.6.5. Quantification of Sterols
2.6.6. Quantification of Tocopherols
2.6.7. Quantification of Total and Individual Phenolic Compounds
2.7. Zeta Potential Measurements
2.8. Statistical Analysis
3. Results and Discussion
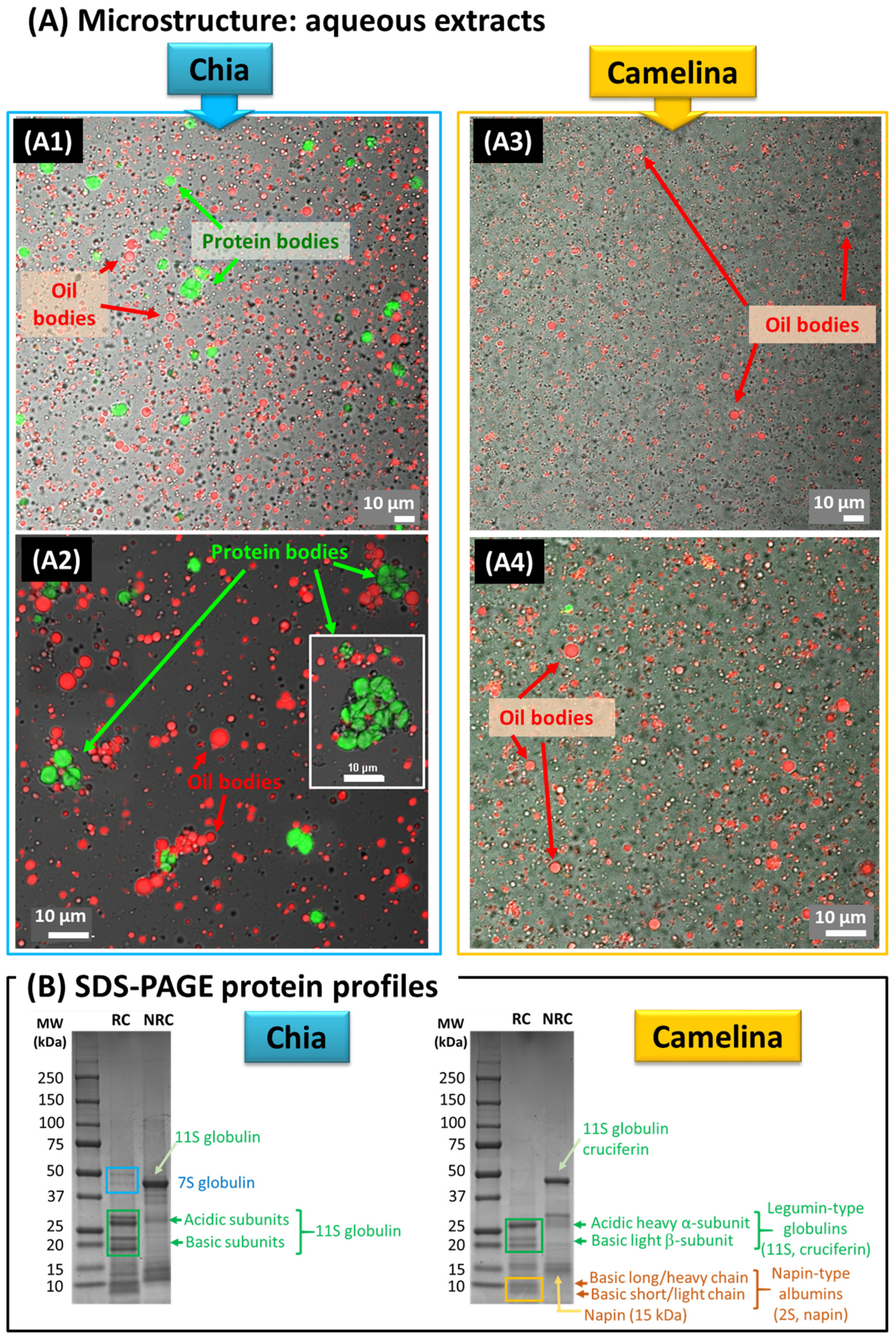
3.1. Microstructure of Chia and Camelina Seed Aqueous Extracts and OBs
3.1.1. Aqueous Extracts Obtained from Chia and Camelina Seeds
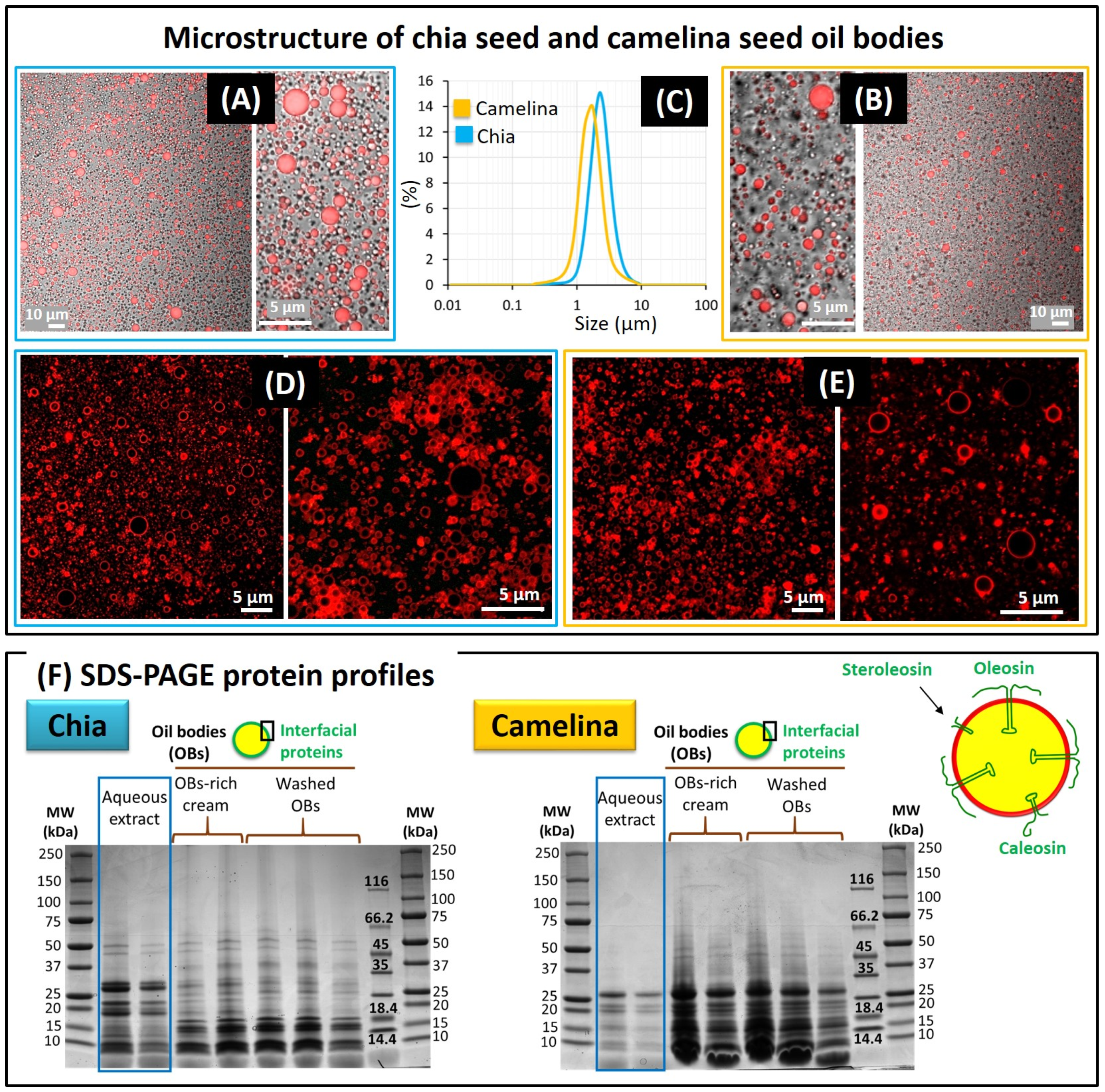
3.1.2. Oil Bodies Recovered from the Chia and Camelina Seed Aqueous Extracts
3.2. Composition
3.2.1. Lipid Content of the Chia and Camelina Seeds
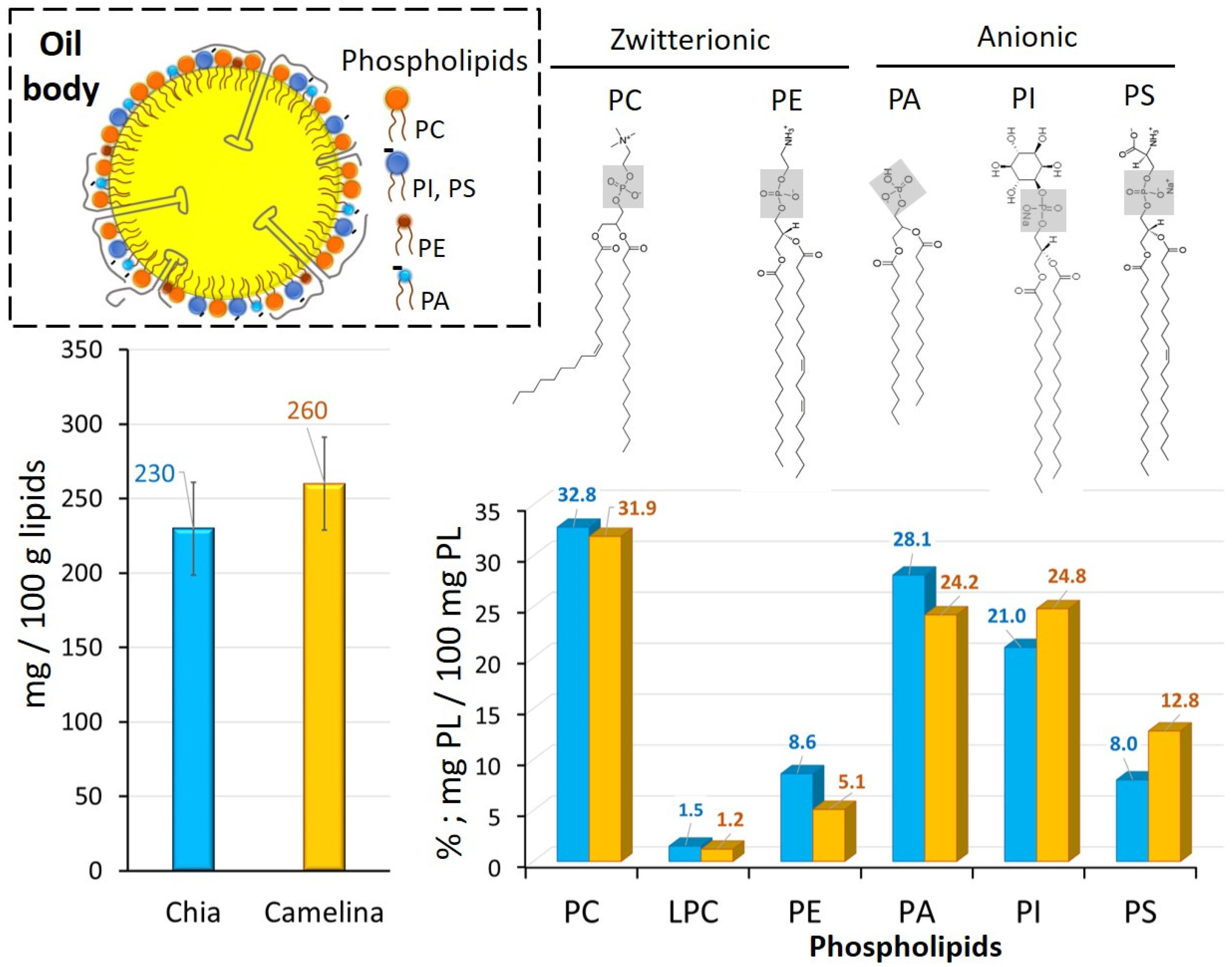
3.2.2. Phospholipid Composition of OBs
3.2.3. Fatty Acid and Triacylglycerol Composition of OBs
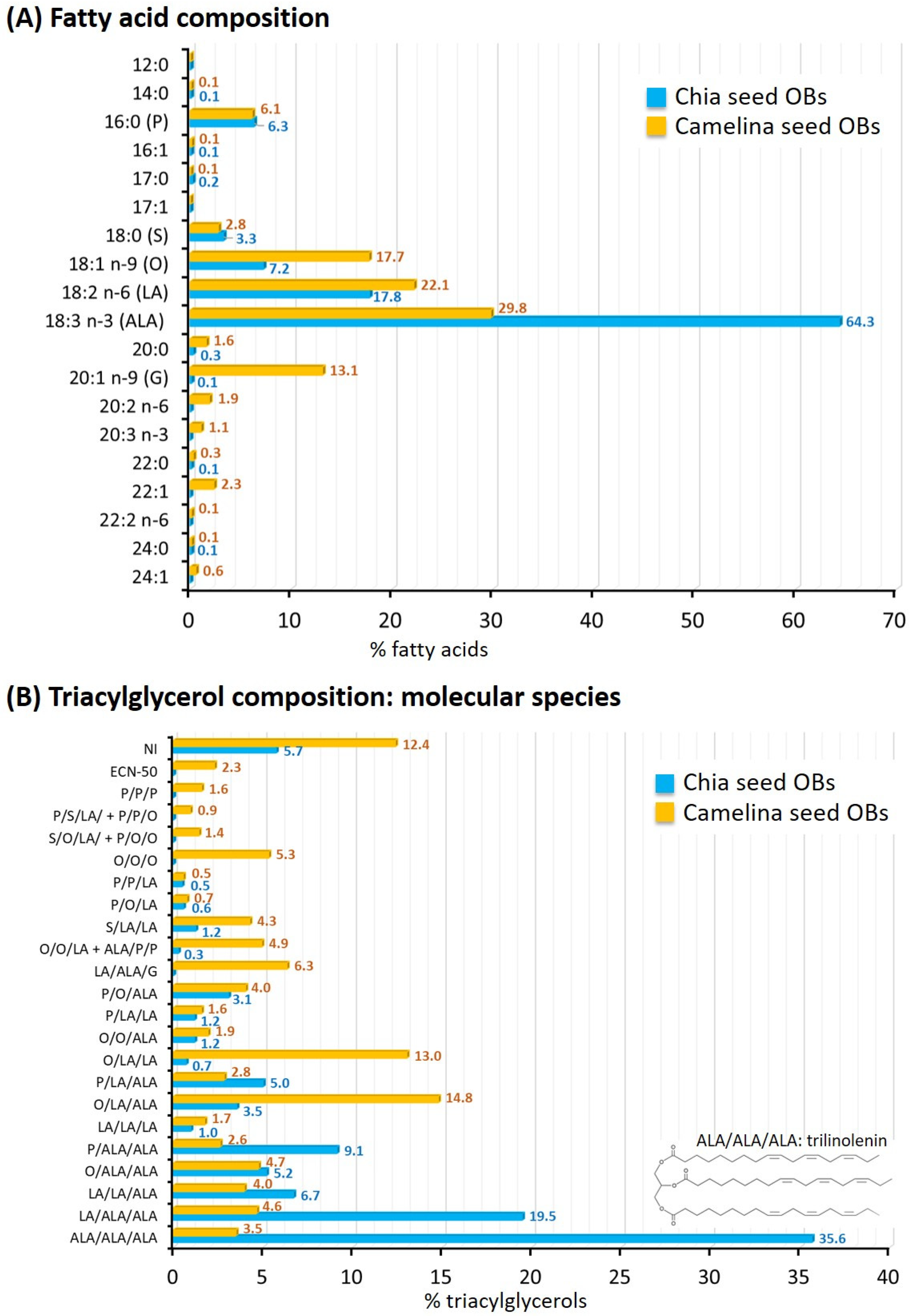
Fatty Acid Composition
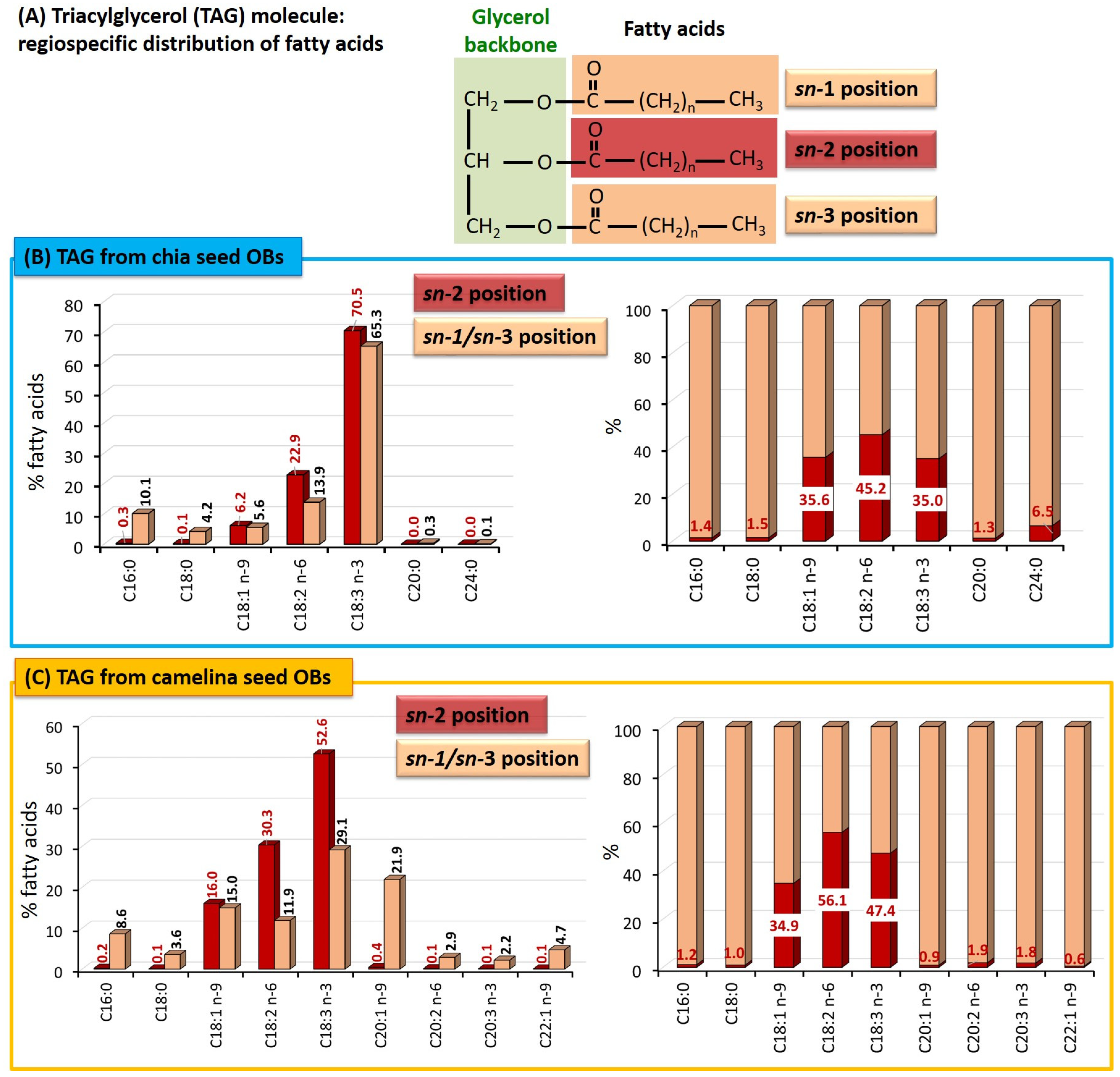
Triacylglycerols: Molecular Species and Distribution of the Fatty Acids on the Glycerol Backbone
3.2.4. Plant Sterols, Tocopherols and Phenolic Compounds
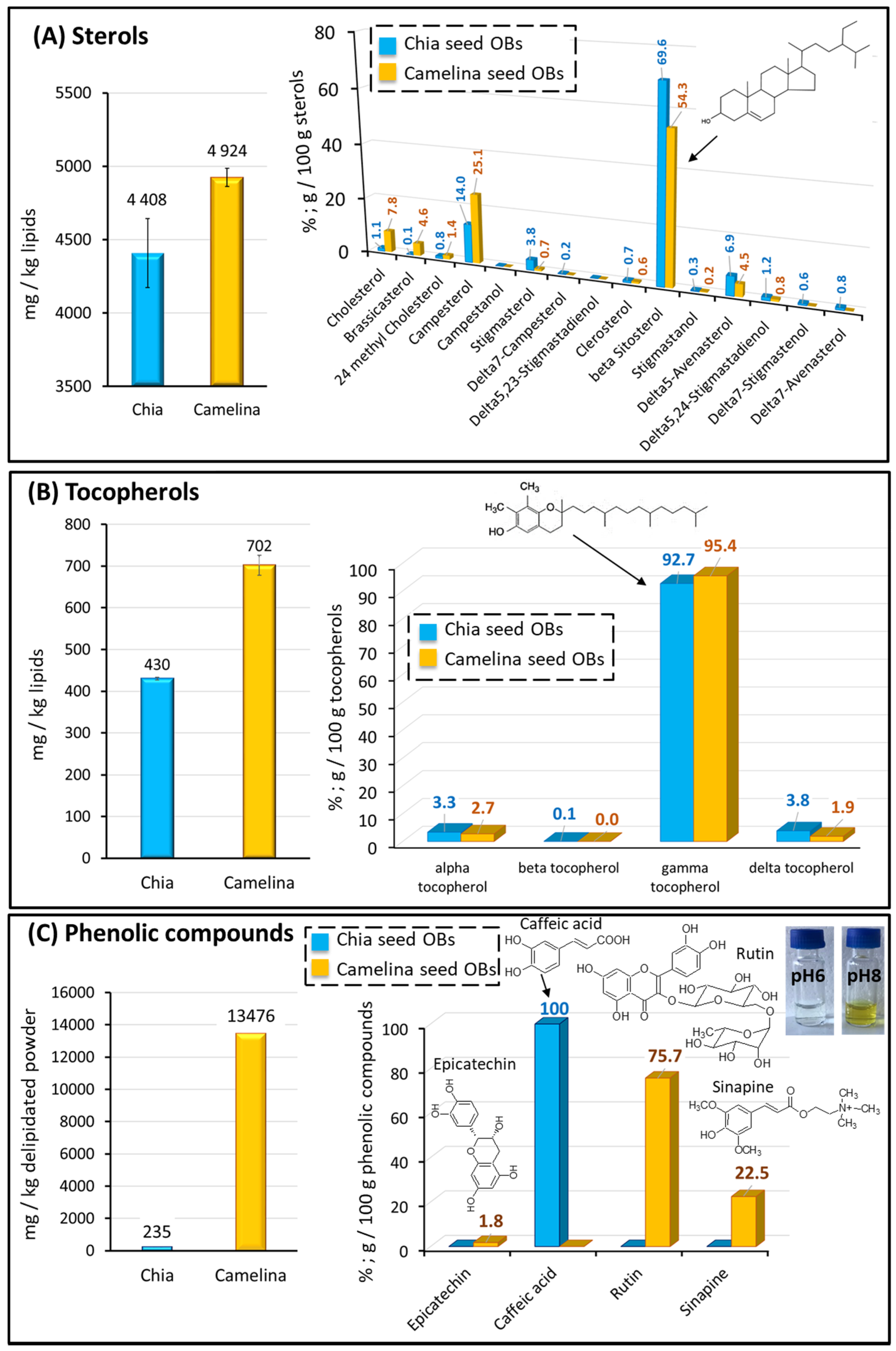
Plant Sterols
Tocopherols
Phenolic Compounds
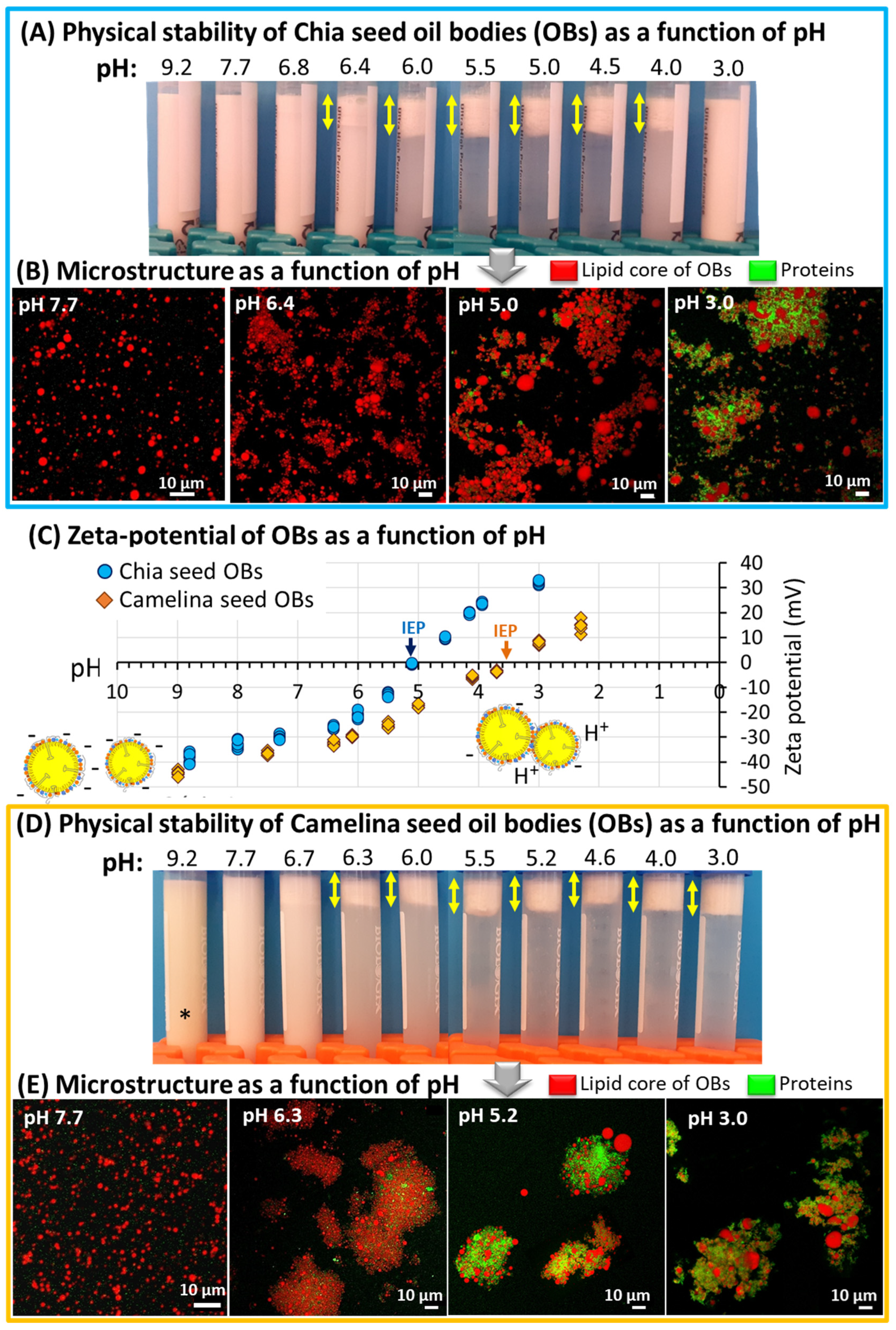
3.3. Physical Stability of Chia and Camelina Seed OBs as a Function of pH: Toward Innovative and Sustainable Food Applications
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopez, C.; Novales, B.; Rabesona, H.; Weber, M.; Chardot, T.; Anton, M. Deciphering the Properties of Hemp Seed Oil Bodies for Food Applications: Lipid Composition, Microstructure, Surface Properties and Physical Stability. Food Res. Int. 2021, 150, 110759. [Google Scholar] [CrossRef] [PubMed]
- Nikiforidis, C.V. Structure and Functions of Oleosomes (Oil Bodies). Adv. Colloid Interface Sci. 2019, 274, 102039. [Google Scholar] [CrossRef] [PubMed]
- Nikiforidis, C.V.; Matsakidou, A.; Kiosseoglou, V. Composition, Properties and Potential Food Applications of Natural Emulsions and Cream Materials Based on Oil Bodies. RSC Adv. 2014, 4, 25067–25078. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Fani, A.; Dave, A.; Singh, H. Nature-Assembled Structures for Delivery of Bioactive Compounds and Their Potential in Functional Foods. Front. Chem. 2020, 8, 564021. [Google Scholar] [CrossRef]
- Tzen, J.; Cao, Y.; Laurent, P.; Ratnayake, C.; Huang, A. Lipids, Proteins, and Structure of Seed Oil Bodies from Diverse Species. Plant Physiol. 1993, 101, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzen, J.T.; Lie, G.C.; Huang, A.H. Characterization of the Charged Components and Their Topology on the Surface of Plant Seed Oil Bodies. J. Biol. Chem. 1992, 267, 15626–15634. [Google Scholar] [CrossRef]
- Kapchie, V.N.; Yao, L.; Hauck, C.C.; Wang, T.; Murphy, P.A. Oxidative Stability of Soybean Oil in Oleosomes as Affected by PH and Iron. Food Chem. 2013, 141, 2286–2293. [Google Scholar] [CrossRef]
- Abad, A.; Shahidi, F. Fatty Acid, Triacylglycerol and Minor Component Profiles Affect Oxidative Stability of Camelina and Sophia Seed Oils. Food Biosci. 2021, 40, 100849. [Google Scholar] [CrossRef]
- Ciftci, O.N.; Przybylski, R.; Rudzińska, M. Lipid Components of Flax, Perilla, and Chia Seeds. Eur. J. Lipid Sci. Technol. 2012, 114, 794–800. [Google Scholar] [CrossRef]
- Gravé, G.; Mouloungui, Z.; Cerny, M.; Lacroux, E.; Valentin, R.; Fabre, J.-F.; Merah, O. Oil Content, Fatty Acid and Phytosterol Compositions of Chia Seeds Cultivated under Organic Conditions in France. OCL 2022, 29, 32. [Google Scholar] [CrossRef]
- De Falco, B.; Amato, M.; Lanzotti, V. Chia Seeds Products: An Overview. Phytochem. Rev. 2017, 16, 745–760. [Google Scholar] [CrossRef]
- Kulczyński, B.; Kobus-Cisowska, J.; Taczanowski, M.; Kmiecik, D.; Gramza-Michałowska, A. The Chemical Composition and Nutritional Value of Chia Seeds—Current State of Knowledge. Nutrients 2019, 11, 1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, R.; Nadeem, M.; Khalique, A.; Imran, M.; Mehmood, S.; Javid, A.; Hussain, J. Nutritional and Therapeutic Perspectives of Chia (Salvia hispanica L.): A Review. J. Food Sci. Technol. 2016, 53, 1750–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, L.D.A.; Verneque, B.J.F.; Mota, A.P.L.; Duarte, C.K. Chia Seed (Salvia hispanica L.) Consumption and Lipid Profile: A Systematic Review and Meta-Analysis. Food Funct. 2021, 12, 8835–8849. [Google Scholar] [CrossRef] [PubMed]
- Teoh, S.L.; Lai, N.M.; Vanichkulpitak, P.; Vuksan, V.; Ho, H.; Chaiyakunapruk, N. Clinical Evidence on Dietary Supplementation with Chia Seed (Salvia hispanica L.): A Systematic Review and Meta-Analysis. Nutr. Rev. 2018, 76, 219–242. [Google Scholar] [CrossRef] [PubMed]
- EC Commission. Implementing Decision of 22 January 2013 Authorising an Extension of Use of Chia (Salvia hispanica) Seed as a Novel Food Ingredient under Regulation (EC) No 258/97 of the European Parliament and of the Council (Notified under Document C (2013) 123); European Commission: Brussels, Belgium, 2013; Volume 2. [Google Scholar]
- EC Commission. Decision of 13 October 2009 Authorising the Placing on the Market of Chia Seed (Salvia Hispanica) as Novel Food Ingredient under Regulation (EC) No 258/97 of the European Parliament and of the Council (Notified under Document C(2009) 7645); European Commission: Brussels, Belgium, 2009; Volume 2. [Google Scholar]
- Arshad, M.; Mohanty, K.A.; Acker, R.V.; Riddle, R.; Todd, J.; Khalil, H.; Misra, M. Valorization of Camelina Oil to Biobased Materials and Biofuels for New Industrial Uses: A Review. RSC Adv. 2022, 12, 27230–27245. [Google Scholar] [CrossRef]
- Neupane, D.; Lohaus, R.H.; Solomon, J.K.Q.; Cushman, J.C. Realizing the Potential of Camelina Sativa as a Bioenergy Crop for a Changing Global Climate. Plants 2022, 11, 772. [Google Scholar] [CrossRef]
- Veljković, V.B.; Kostić, M.D.; Stamenković, O.S. Camelina Seed Harvesting, Storing, Pretreating, and Processing to Recover Oil: A Review. Ind. Crops Prod. 2022, 178, 114539. [Google Scholar] [CrossRef]
- Righini, D.; Zanetti, F.; Monti, A. The Bio-Based Economy Can Serve as the Springboard for Camelina and Crambe to Quit the Limbo. OCL 2016, 23, D504. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, F.; Alberghini, B.; Marjanović Jeromela, A.; Grahovac, N.; Rajković, D.; Kiprovski, B.; Monti, A. Camelina, an Ancient Oilseed Crop Actively Contributing to the Rural Renaissance in Europe. A Review. Agron. Sustain. Dev. 2021, 41, 2. [Google Scholar] [CrossRef]
- Garcia, F.L.; Ma, S.; Dave, A.; Acevedo-Fani, A. Structural and Physicochemical Characteristics of Oil Bodies from Hemp Seeds (Cannabis sativa L.). Foods 2021, 10, 2930. [Google Scholar] [CrossRef] [PubMed]
- Nikiforidis, C.V.; Kiosseoglou, V. Aqueous Extraction of Oil Bodies from Maize Germ (Zea mays) and Characterization of the Resulting Natural Oil-in-Water Emulsion. J. Agric. Food Chem. 2009, 57, 5591–5596. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.A.; Sinnecker, P.; Cavalari, A.A.; Sato, A.C.K.; Perrechil, F.A. Extraction of Chia Seed Mucilage: Effect of Ultrasound Application. Food Chem. Adv. 2022, 1, 100024. [Google Scholar] [CrossRef]
- Lopez, C.; Madec, M.-N.; Jiménez-Flores, R. Lipid Rafts in the Bovine Milk Fat Globule Membrane Revealed by the Lateral Segregation of Phospholipids and Heterogeneous Distribution of Glycoproteins. Food Chem. 2010, 120, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Lawrence, A.-M.; Besir, H. Staining of Proteins in Gels with Coomassie G-250 without Organic Solvent and Acetic Acid. J. Vis. Exp. 2009, 30, 1350. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- American Oil Chemists’ Society. Ce5c-93 AOCS Official Method Ce 5c-93; Individual Triglycerides in Oils and Fats by HPLC; AOCS: Urbana, IL, USA, 2022. [Google Scholar]
- Japan Oil Chemists’ Society; American Oil Chemists’ Society. Joint JOCS/AOCS Official Method Ch 3a-19 Determination of the Composition of Fatty Acids at the 2-Position of Oils and Fats-Enzymatic Transesterification Method Using Candida antarctica Lipase. In Official Methods and Recommended Practices of the AOCS; AOCS: Urbana, IL, USA, 2019. [Google Scholar]
- Mochizuki, M.; Matsuyama, H.; Suzuki, K.; Watanabe, Y. Fatty Acid Composition and Distribution in Triacylglycerols of Cultured Fishes in Japan. Biocatal. Agric. Biotechnol. 2021, 31, 101687. [Google Scholar] [CrossRef]
- Watanabe, Y.; Sato, S.; Asada, M.; Arishima, T.; Iida, Y.; Imagi, J.; Saito, K.; Sano, T.; Sasaki, A.; Sasaki, R.; et al. Enzymatic Analysis of Positional Fatty Acid Distributions in Triacylglycerols by 1(3)-Selective Transesterification with Candida antarctica Lipase B: A Collaborative Study. J. Oleo Sci. 2015, 64, 1193–1205. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Sato, S.; Sera, S.; Sato, C.; Yoshinaga, K.; Nagai, T.; Sato, R.; Iwasaka, H.; Aki, T. Enzymatic Analysis of Positional Distribution of Fatty Acids in Solid Fat by 1,3-Selective Transesterification with Candida antarctica Lipase B. J. Am. Oil Chem. Soc. 2014, 91, 1323–1330. [Google Scholar] [CrossRef]
- American Oil Chemists’ Society. Official Method Ch 1-91 Preparation of Methyl Esters of Long-Chain Fatty Acids; AOCS: Urbana, IL, USA, 2017. [Google Scholar]
- Lopez, C.; Briard-Bion, V.; Ménard, O. Polar Lipids, Sphingomyelin and Long-Chain Unsaturated Fatty Acids from the Milk Fat Globule Membrane Are Increased in Milks Produced by Cows Fed Fresh Pasture Based Diet during Spring. Food Res. Int. 2014, 58, 59–68. [Google Scholar] [CrossRef]
- ISO-122228-1; Determination of Individual and Total Sterols Contents—Gas Chromatographic Method—Part 1: Animal and Vegetable Fats and Oils; ISO: Geneva, Switzerland, 2014; Available online: https://www.iso.org/cms/render/live/en/sites/isoorg/contents/data/standard/06/02/60248.html (accessed on 21 May 2021).
- Le Deun, E.; Van der Werf, R.; Le Bail, G.; Le Quéré, J.-M.; Guyot, S. HPLC-DAD-MS Profiling of Polyphenols Responsible for the Yellow-Orange Color in Apple Juices of Different French Cider Apple Varieties. J. Agric. Food Chem. 2015, 63, 7675–7684. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Oliveros, M.R.; Paredes-López, O. Isolation and Characterization of Proteins from Chia Seeds (Salvia hispanica L.). J. Agric. Food Chem. 2013, 61, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Boyle, C.; Hansen, L.; Hinnenkamp, C.; Ismail, B.P. Emerging Camelina Protein: Extraction, Modification, and Structural/Functional Characterization. J. Am. Oil Chem. Soc. 2018, 95, 1049–1062. [Google Scholar] [CrossRef]
- Ngo, N.T.T.; Shahidi, F. Functional Properties of Protein Isolates from Camelina (Camelina sativa (L.) Crantz) and Flixweed (Sophia, Descurainis sophia L.) Seed Meals. Food Prod. Process. Nutr. 2021, 3, 31. [Google Scholar] [CrossRef]
- Wanasundara, J.P.D. Proteins of Brassicaceae Oilseeds and Their Potential as a Plant Protein Source. Crit. Rev. Food Sci. Nutr. 2011, 51, 635–677. [Google Scholar] [CrossRef]
- Yang, N.; Su, C.; Zhang, Y.; Jia, J.; Leheny, R.L.; Nishinari, K.; Fang, Y.; Phillips, G.O. In Situ Nanomechanical Properties of Natural Oil Bodies Studied Using Atomic Force Microscopy. J. Colloid Interface Sci. 2020, 570, 362–374. [Google Scholar] [CrossRef]
- Cassen, A.; Fabre, J.-F.; Lacroux, E.; Cerny, M.; Vaca-Medina, G.; Mouloungui, Z.; Merah, O.; Valentin, R. Aqueous Integrated Process for the Recovery of Oil Bodies or Fatty Acid Emulsions from Sunflower Seeds. Biomolecules 2022, 12, 149. [Google Scholar] [CrossRef]
- Da Silva Marineli, R.; Moraes, É.A.; Lenquiste, S.A.; Godoy, A.T.; Eberlin, M.N.; Maróstica, M.R., Jr. Chemical Characterization and Antioxidant Potential of Chilean Chia Seeds and Oil (Salvia hispanica L.). LWT Food Sci. Technol. 2014, 59, 1304–1310. [Google Scholar] [CrossRef]
- Shen, Y.; Zheng, L.; Jin, J.; Li, X.; Fu, J.; Wang, M.; Guan, Y.; Song, X. Phytochemical and Biological Characteristics of Mexican Chia Seed Oil. Molecules 2018, 23, 3219. [Google Scholar] [CrossRef]
- Kuznetcova, D.V.; Linder, M.; Jeandel, C.; Paris, C.; Desor, F.; Baranenko, D.A.; Nadtochii, L.A.; Arab-Tehrany, E.; Yen, F.T. Nanoliposomes and Nanoemulsions Based on Chia Seed Lipids: Preparation and Characterization. Int. J. Mol. Sci. 2020, 21, 9079. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.V.; Villanueva-Bermejo, D.; Castro-Gómez, P.; Fornari, T.; Fontecha, J. Appraisal of the Suitability of Two-Stage Extraction Process by Combining Compressed Fluid Technologies of Polar Lipid Fractions from Chia Seed. Food Res. Int. 2020, 131, 109007. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.; Shrestha, P.; Belide, S.; Petrie, J.; Nichols, P.; Singh, S. Characterization of Oilseed Lipids from “DHA-Producing Camelina sativa”: A New Transformed Land Plant Containing Long-Chain Omega-3 Oils. Nutrients 2014, 6, 776–789. [Google Scholar] [CrossRef] [Green Version]
- Abad, A.; Shahidi, F. Compositional Characteristics and Oxidative Stability of Chia Seed Oil (Salvia hispanica L). Food Prod. Process. Nutr. 2020, 2, 9. [Google Scholar] [CrossRef]
- Moser, B.R. Camelina (Camelina sativa L.) Oil as a Biofuels Feedstock: Golden Opportunity or False Hope? Lipid Technol. 2010, 22, 270–273. [Google Scholar] [CrossRef]
- Ostrikov, A.N.; Kleimenova, N.L.; Kopylov, M.V.; Bolgova, I.N. The Study of the Fatty Acid Composition of Camelina Oil Obtained by Cold Pressing. IOP Conf. Ser. Earth Environ. Sci. 2021, 640, 042009. [Google Scholar] [CrossRef]
- Rodriguez-Rodriguez, M.F.; Sánchez-García, A.; Salas, J.J.; Garcés, R.; Martínez-Force, E. Characterization of the Morphological Changes and Fatty Acid Profile of Developing Camelina sativa Seeds. Ind. Crops Prod. 2013, 50, 673–679. [Google Scholar] [CrossRef]
- Popa, A.-L.; Drumea, V.; Niţă, R.-A.; Florea, M.; Olariu, L.; Jurcoane, Ş.; Cristea, S. A Physico-Chemical Characterization of Oil from Camelina sativa Seeds Grown in Romania. Rom. Biotechnol. Lett. 2019, 24, 776–782. [Google Scholar] [CrossRef]
- Nitrayová, S.; Brestenský, M.; Heger, J.; Patráš, P.; Rafay, J.; Sirotkin, A. Amino Acids and Fatty Acids Profile of Chia (Salvia hispanica L.) and Flax (Linum usitatissimum L.) Seed. Potravin. Slovak J. Food Sci. 2014, 8, 72–76. [Google Scholar] [CrossRef]
- Ayerza, R. Oil Content and Fatty Acid Composition of Chia (Salvia hispanica L.) from Five Northwestern Locations in Argentina. J. Am. Oil Chem. Soc. 1995, 72, 1079–1081. [Google Scholar] [CrossRef]
- Alfieri, A.; Imperlini, E.; Nigro, E.; Vitucci, D.; Orrù, S.; Daniele, A.; Buono, P.; Mancini, A. Effects of Plant Oil Interesterified Triacylglycerols on Lipemia and Human Health. Int. J. Mol. Sci. 2018, 19, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, H.L.; Hoy, C.E. The Digestion of Dietary Triacylglycerols. Prog. Lipid Res. 2004, 43, 105–133. [Google Scholar] [CrossRef] [PubMed]
- Berry, S.E.E. Triacylglycerol Structure and Interesterification of Palmitic and Stearic Acid-Rich Fats: An Overview and Implications for Cardiovascular Disease. Nutr. Res. Rev. 2009, 22, 3–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, J.E. Studies on Effects of Dietary Fatty Acids as Related to Their Position on Triglycerides. Lipids 2001, 36, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Michalski, M.C.; Genot, C.; Gayet, C.; Lopez, C.; Fine, F.; Joffre, F.; Vendeuvre, J.L.; Bouvier, J.; Chardigny, J.M.; Raynal-Ljutovac, K. Multiscale Structures of Lipids in Foods as Parameters Affecting Fatty Acid Bioavailability and Lipid Metabolism. Prog. Lipid Res. 2013, 52, 354–373. [Google Scholar] [CrossRef] [Green Version]
- Gylling, H.; Plat, J.; Turley, S.; Ginsberg, H.N.; Ellegård, L.; Jessup, W.; Jones, P.J.; Lütjohann, D.; Maerz, W.; Masana, L.; et al. Plant Sterols and Plant Stanols in the Management of Dyslipidaemia and Prevention of Cardiovascular Disease. Atherosclerosis 2014, 232, 346–360. [Google Scholar] [CrossRef]
- Oliveira-Alves, S.C.; Vendramini-Costa, D.B.; Betim Cazarin, C.B.; Maróstica Júnior, M.R.; Borges Ferreira, J.P.; Silva, A.B.; Prado, M.A.; Bronze, M.R. Characterization of Phenolic Compounds in Chia (Salvia hispanica L.) Seeds, Fiber Flour and Oil. Food Chem. 2017, 232, 295–305. [Google Scholar] [CrossRef]
- Taga, M.S.; Miller, E.E.; Pratt, D.E. Chia Seeds as a Source of Natural Lipid Antioxidants. J. Am. Oil Chem. Soc. 1984, 61, 928–931. [Google Scholar] [CrossRef]
- Semwal, R.; Joshi, S.K.; Semwal, R.B.; Semwal, D.K. Health Benefits and Limitations of Rutin—A Natural Flavonoid with High Nutraceutical Value. Phytochem. Lett. 2021, 46, 119–128. [Google Scholar] [CrossRef]
- Matthäus, B.; Zubr, J. Variability of Specific Components in Camelina sativa Oilseed Cakes. Ind. Crops Prod. 2000, 12, 9–18. [Google Scholar] [CrossRef]
- Salminen, H.; Estévez, M.; Kivikari, R.; Heinonen, M. Inhibition of Protein and Lipid Oxidation by Rapeseed, Camelina and Soy Meal in Cooked Pork Meat Patties. Eur. Food Res. Technol. 2006, 223, 461–468. [Google Scholar] [CrossRef]
- Salminen, H.; Heinonen, M. Plant Phenolics Affect Oxidation of Tryptophan. J. Agric. Food Chem. 2008, 56, 7472–7481. [Google Scholar] [CrossRef] [PubMed]
- Terpinc, P.; Polak, T.; Makuc, D.; Ulrih, N.P.; Abramovič, H. The Occurrence and Characterisation of Phenolic Compounds in Camelina sativa Seed, Cake and Oil. Food Chem. 2012, 131, 580–589. [Google Scholar] [CrossRef]
- Ratusz, K.; Symoniuk, E.; Wroniak, M.; Rudzińska, M. Bioactive Compounds, Nutritional Quality and Oxidative Stability of Cold-Pressed Camelina (Camelina sativa L.) Oils. Appl. Sci. 2018, 8, 2606. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez, C.; Sotin, H.; Rabesona, H.; Novales, B.; Le Quéré, J.-M.; Froissard, M.; Faure, J.-D.; Guyot, S.; Anton, M. Oil Bodies from Chia (Salvia hispanica L.) and Camelina (Camelina sativa L.) Seeds for Innovative Food Applications: Microstructure, Composition and Physical Stability. Foods 2023, 12, 211. https://doi.org/10.3390/foods12010211
Lopez C, Sotin H, Rabesona H, Novales B, Le Quéré J-M, Froissard M, Faure J-D, Guyot S, Anton M. Oil Bodies from Chia (Salvia hispanica L.) and Camelina (Camelina sativa L.) Seeds for Innovative Food Applications: Microstructure, Composition and Physical Stability. Foods. 2023; 12(1):211. https://doi.org/10.3390/foods12010211
Chicago/Turabian StyleLopez, Christelle, Hélène Sotin, Hanitra Rabesona, Bruno Novales, Jean-Michel Le Quéré, Marine Froissard, Jean-Denis Faure, Sylvain Guyot, and Marc Anton. 2023. "Oil Bodies from Chia (Salvia hispanica L.) and Camelina (Camelina sativa L.) Seeds for Innovative Food Applications: Microstructure, Composition and Physical Stability" Foods 12, no. 1: 211. https://doi.org/10.3390/foods12010211