

Queijo Serra da Estrela PDO Cheese: Investigation into Its Morpho-Textural Traits, Microbiota, and Volatilome

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling of Cheese
2.2. Evaluation of Physico-Chemical Features
2.3. Analysis of Morpho-Textural Parameters
2.4. Microbiological Analyses
2.5. DNA Extraction and Sequencing
2.6. Isolation and Characterization of the Dominant Non-Starter Lactic Acid Bacteria (NSLAB)
2.6.1. Collection of Pure Cultures
2.6.2. Semi-Quantitative Assessment of the Enzymatic Activity of NSLAB
2.7. Cheese Volatile Profile
2.8. Statistical Analysis
3. Results
3.1. Physico-Chemical Characterization
3.2. Morpho-Textural Characterization
3.3. Viable Counting
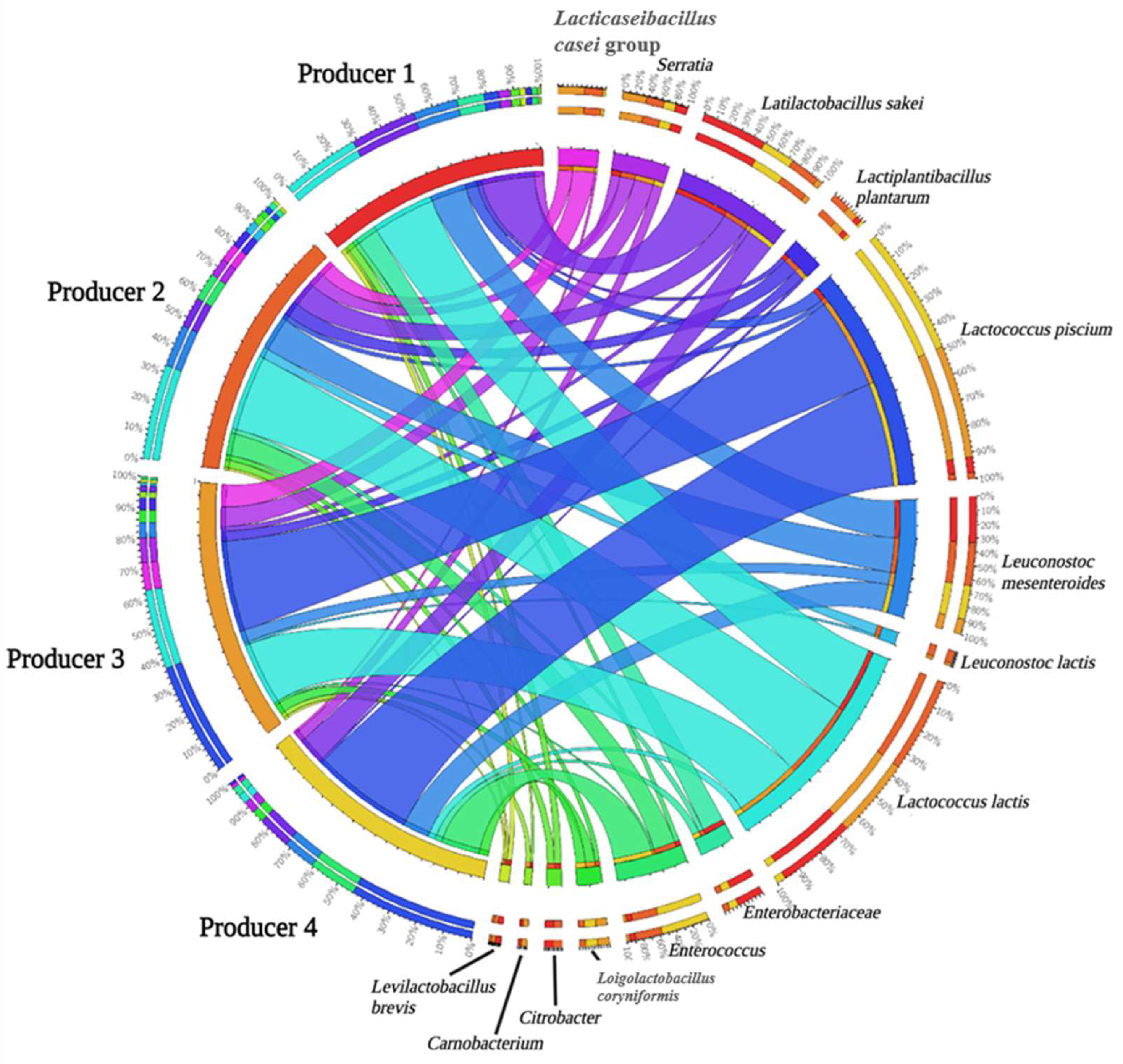
3.4. Microbiota Composition
3.5. Isolation Campaign and Semi-Quantitative Assessment of Enzymatic Activity of Pure Cultures
3.6. Volatile Profile
4. Discussion
4.1. Physico-Chemical and Morpho-Textural Traits of Cheeses
4.2. Bacterial Biota
4.2.1. Enzymatic Activity of Lactic Acid Bacteria Isolates
Enterococcus spp.
Levilactobacillus brevis
Lacticaseibacillus casei Group
Latilactobacillus graminis
Leuconostoc mesenteroides
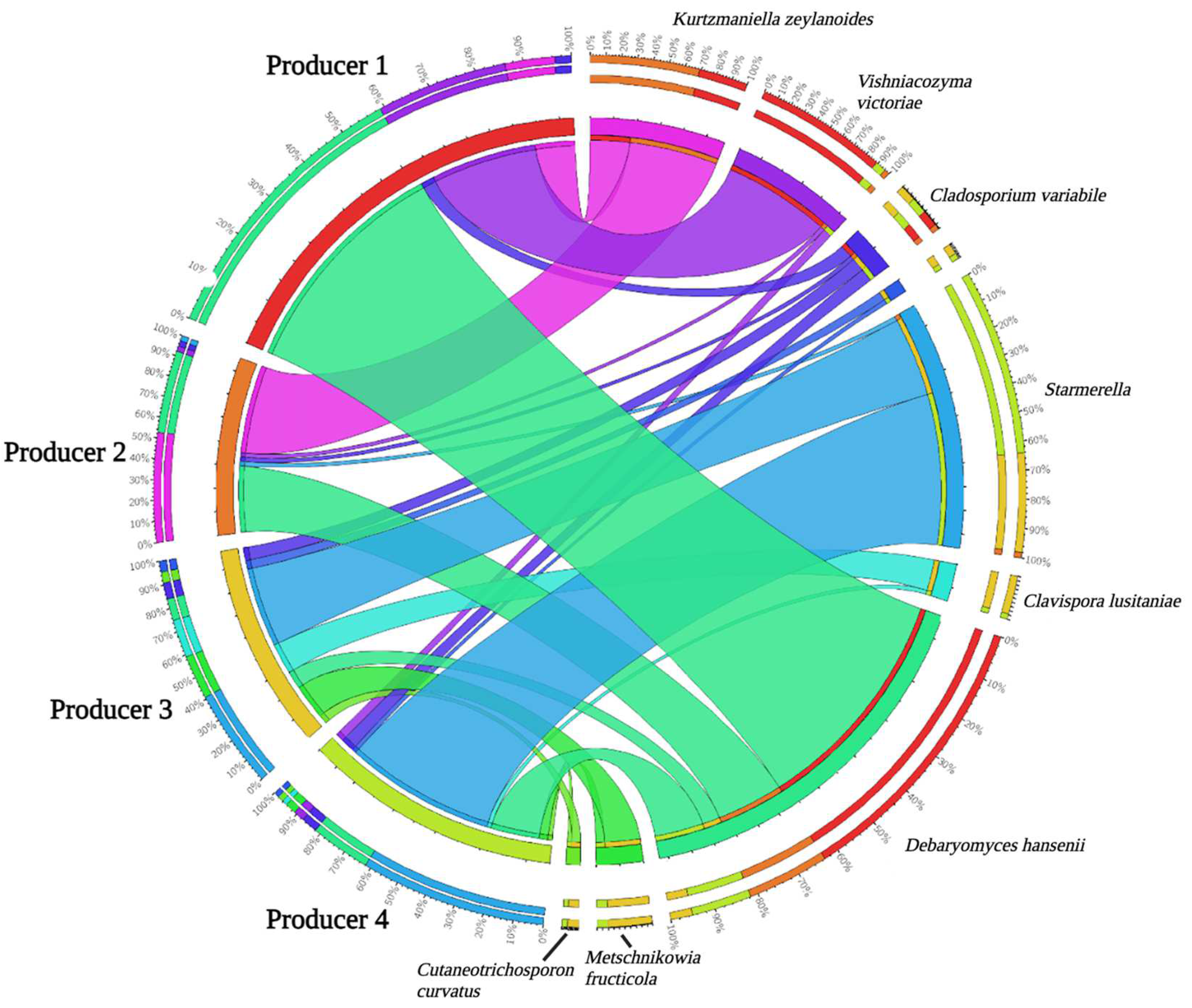
4.3. Mycobiota
4.4. Volatile Organic Compounds
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Laranjo, M.; Potes, M.E. Chapter 6—Traditional Mediterranean cheeses: Lactic acid bacteria populations and functional traits. In Applied Biotechnology Reviews, Lactic Acid Bacteria in Food Biotechnology; Ray, R.C., Paramithiotis, S., Azevedo, V.A.D.C., Montet, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 97–124. [Google Scholar] [CrossRef]
- Boyazoglu, J.; Morand-Fehr, P. Mediterranean dairy sheep and goat products and their quality: A critical review. Small Rumin. Res. 2001, 40, 1–11. [Google Scholar] [CrossRef]
- Possas, A.; Bonilla-Luque, O.M.; Valero, A. From cheese-making to consumption: Exploring the microbial safety of cheeses through predictive microbiology models. Foods 2021, 10, 355. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Y.; Guan, R.; Jia, G.; Ma, Y.; Zhang, Y. Advances in research on calf rennet substitutes and their effects on cheese quality. Food Res. Int. 2021, 149, 110704. [Google Scholar] [CrossRef]
- Cardinali, F.; Foligni, R.; Ferrocino, I.; Harasym, J.; Orkusz, A.; Milanović, V.; Franciosa, I.; Garofalo, C.; Mannozzi, C.; Mozzon, M.; et al. Microbiological, morpho-textural, and volatile characterization of Portuguese Queijo de Nisa PDO cheese. Food Res. Int. 2022, 162, 112011. [Google Scholar] [CrossRef]
- Johnson, M.E. A 100-Year Review: Cheese production and quality. J. Dairy Sci. 2017, 100, 9952–9965. [Google Scholar] [CrossRef]
- Akalin, A.S.; Gönç, S.; Akbaş, Y. Variation in organic acids content during ripening of pickled white cheese. J. Dairy Sci. 2002, 85, 1670–1676. [Google Scholar] [CrossRef] [Green Version]
- Aquilanti, L.; Kahraman, O.; Zannini, E.; Osimani, A.; Silvestri, G.; Ciarrocchi, F.; Garofalo, C.; Tekin, E.; Clementi, F. Response of lactic acid bacteria to milk fortification with dietary zinc salts. Int. Dairy J. 2012, 25, 52–59. [Google Scholar] [CrossRef]
- Steele, J.; Broadbent, J.; Kok, J. Perspectives on the contribution of lactic acid bacteria to cheese flavor development. Curr. Opin. Biotechnol. 2013, 24, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Mancini, L.; Fox, P.F. Pros and cons for using non-starter lactic acid bacteria (NSLAB) as secondary/adjunct starters for cheese ripening. Trends Food Sci. Technol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- European Union. Commission Regulation (EC) No 1107/96 of 12 June 1996 on the Registration of Geographical Indications and Designations of Origin under the Procedure Laid Down in Article 17 of Council Regulation (EEC) No 2081/92. Off. J. Eur. Commun. 1992, 148, 1–10. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:31996R1107&from=EN (accessed on 28 November 2022).
- eAmbrosia. The EU Geographical Indications Register. 2022. Available online: https://ec.europa.eu/info/food-farming-fisheries/food-safety-and-quality/certification/quality-labels/geographical-indications-register/details/EUGI00000013223 (accessed on 19 October 2022).
- Macedo, A.C.; Malcata, F.X.; Oliveira, J.C. The technology, chemistry, and microbiology of Serra cheese: A review. J. Dairy Sci. 1993, 76, 1725–1739. [Google Scholar] [CrossRef]
- Fogeiro, É.; Barracosa, P.; Oliveira, J.; Wessel, D.F. Influence of cardoon flower (Cynara cardunculus L.) and flock lactation stage in PDO Serra da Estrela Cheese. Foods 2020, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Macedo, A.C.; Tavares, T.G.; Malcata, F.X. Influence of native lactic acid bacteria on the microbiological, biochemical and sensory profiles of Serra da Estrela cheese. Food Microbiol. 2004, 21, 233–240. [Google Scholar] [CrossRef]
- Tavaria, F.K.; Malcata, F.X. On the microbiology of Serra da Estrela cheese: Geographical and chronological considerations. Food Microbiol. 2000, 17, 293–304. [Google Scholar] [CrossRef]
- Parker, M.L.; Gunning, P.A.; Macedo, A.C.; Malcata, F.X.; Brocklehurst, T.F. The microstructure and distribution of micro-organisms within mature Serra cheese. J. Appl. Microbiol. 1998, 84, 523–530. [Google Scholar] [CrossRef]
- Reis, P.J.M.; Malcata, F.X. Ripening-related changes in Serra da Estrela cheese: A stereological study. J. Dairy Sci. 2011, 94, 1223–1238. [Google Scholar] [CrossRef] [Green Version]
- Tavaria, F.K.; Malcata, F.X. Microbiological characterization of Serra da Estrela cheese throughout its Appellation d’Origine Protégée region. J. Food Prot. 1998, 61, 601–607. [Google Scholar] [CrossRef]
- Tavaria, F.K.; Reis, P.J.M.; Malcata, F.X. Effect of dairy farm and milk refrigeration on microbiological and microstructural characteristics of matured Serra da Estrela cheese. Int. Dairy J. 2006, 16, 895–902. [Google Scholar] [CrossRef]
- Rocha, R.; Velho, M.V.; Santos, J.; Fernandes, P. Serra da Estrela PDO cheese microbiome as revealed by next generation sequencing. Microorganisms 2021, 9, 2007. [Google Scholar] [CrossRef]
- Belleggia, L.; Milanović, V.; Ferrocino, I.; Cocolin, L.; Haouet, M.N.; Scuota, S.; Maoloni, A.; Garofalo, C.; Cardinali, F.; Aquilanti, L.; et al. Is there any still undisclosed biodiversity in Ciauscolo salami? A new glance into the microbiota of an artisan production as revealed by high-throughput sequencing. Meat Sci. 2020, 165, 108128. [Google Scholar] [CrossRef]
- Harasym, J.; Satta, E.; Kaim, U. Ultrasound treatment of buckwheat grains impacts important functional properties of resulting flour. Molecules 2020, 25, 3012. [Google Scholar] [CrossRef]
- Osimani, A.; Zannini, E.; Aquilanti, L.; Mannazzu, I.; Comitini, F.; Clementi, F. Lactic acid bacteria and yeasts from wheat sourdoughs of the Marche Region. Ital. J. Food Sci. 2009, 21, 269–286. [Google Scholar]
- Cardinali, F.; Foligni, R.; Ferrocino, I.; Harasym, J.; Orkusz, A.; Franciosa, I.; Milanović, V.; Garofalo, C.; Mannozzi, C.; Mozzon, M.; et al. Microbial diversity, morpho-textural characterization, and volatilome profile of the Portuguese thistle-curdled cheese Queijo da Beira Baixa PDO. Food Res. Int. 2022, 157, 111481. [Google Scholar] [CrossRef] [PubMed]
- Osimani, A.; Ferrocino, I.; Corvaglia, M.R.; Roncolini, A.; Milanović, V.; Garofalo, C.; Aquilanti, L.; Riolo, P.; Ruschioni, S.; Jamshidi, E.; et al. Microbial dynamics in rearing trials of Hermetia illucens larvae fed coffee silverskin and microalgae. Food Res. Int. 2021, 140, 110028. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osimani, A.; Garofalo, C.; Aquilanti, L.; Milanović, V.; Clementi, F. Unpasteurised commercial boza as a source of microbial diversity. Int. J. Food Microbiol. 2015, 194, 62–70. [Google Scholar] [CrossRef]
- Edgar, R.C. UCHIME2: Improved chimera prediction for amplicon sequencing. bioRxiv 2016, 074252. [Google Scholar] [CrossRef] [Green Version]
- Belleggia, L.; Aquilanti, L.; Ferrocino, I.; Milanović, V.; Garofalo, C.; Clementi, F.; Cocolin, L.; Mozzon, M.; Foligni, R.; Haouet, M.N.; et al. Discovering microbiota and volatile compounds of surströmming, the traditional Swedish sour herring. Food Microbiol. 2020, 91, 103503. [Google Scholar] [CrossRef]
- Foligni, R.; Mannozzi, C.; Ismaiel, L.; Capelli, F.; Laurita, R.; Tappi, S.; Rosa, M.D.; Mozzon, M. Impact of cold atmospheric plasma (CAP) treatments on the oxidation of pistachio kernel lipids. Foods 2022, 11, 419. [Google Scholar] [CrossRef]
- Mozzon, M.; Foligni, R.; Mannozzi, C. Brewing quality of hop varieties cultivated in Central Italy based on multivolatile fingerprinting and bitter acid content. Foods 2020, 9, 541. [Google Scholar] [CrossRef]
- Maoloni, A.; Milanović, V.; Osimani, A.; Cardinali, F.; Garofalo, C.; Belleggia, L.; Foligni, R.; Mannozzi, C.; Mozzon, M.; Cirlini, M.; et al. Exploitation of sea fennel (Crithmum maritimum L.) for manufacturing of novel high-value fermented preserves. Food Bioprod. Process. 2021, 127, 174–197. [Google Scholar] [CrossRef]
- Ali, A.M.M.; Sant’Ana, A.S.; Bavisetty, S.C.B. Sustainable preservation of cheese: Advanced technologies, physicochemical properties and sensory attributes. Trends Food Sci. Technol. 2022, 129, 306–326. [Google Scholar] [CrossRef]
- Feng, S.; Xiang, S.; Bian, X.; Li, G. Quantitative analysis of total acidity in aqueous lactic acid solutions by direct potentiometric titration. Microchem. J. 2020, 157, 105049. [Google Scholar] [CrossRef]
- Calamari, L.; Gobbi, L.; Bani, P. Improving the prediction ability of FT-MIR spectroscopy to assess titratable acidity in cow’s milk. Food Chem. 2016, 192, 477–484. [Google Scholar] [CrossRef]
- Cardinali, F.; Ferrocino, I.; Milanović, V.; Belleggia, L.; Corvaglia, M.R.; Garofalo, C.; Foligni, R.; Mannozzi, C.; Mozzon, M.; Cocolin, L.; et al. Microbial communities and volatile profile of Queijo de Azeitão PDO cheese, a traditional Mediterranean thistle-curdled cheese from Portugal. Food Res. Int. 2021, 147, 110537. [Google Scholar] [CrossRef]
- Badertscher, R.; Blaser, C.; Noth, P. Validated method for the determination of free volatile carboxylic acids in cheese and bacterial cultures by GC-FID after esterification in aqueous extract and headspace injection. Food Chem. 2023, 398, 133932. [Google Scholar] [CrossRef]
- Pereira, C.I.; Neto, D.M.; Capucho, J.C.; Gião, M.S.; Gomes, A.M.; Malcata, F.X. How three adventitious lactic acid bacteria affect proteolysis and organic acid production in model Portuguese cheeses manufactured from several milk sources and two alternative coagulants. J. Dairy Sci. 2010, 93, 1335–1344. [Google Scholar] [CrossRef] [Green Version]
- Ayyash, M.; Abu-Jdayil, B.; Hamed, F.; Shaker, R. Rheological, textural, microstructural and sensory impact of exopolysaccharide-producing Lactobacillus plantarum isolated from camel milk on low-fat akawi cheese. LWT 2018, 87, 423–431. [Google Scholar] [CrossRef]
- Gontijo, M.T.P.; Silva, J.d.S.; Vidigal, P.M.P.; Martin, J.G.P. Phylogenetic distribution of the bacteriocin repertoire of lactic acid bacteria species associated with artisanal cheese. Food Res. Int. 2020, 128, 108783. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.A.d.; Muller, M.P.; Berghahn, E.; Souza, C.F.V.d.; Granada, C.E. New enterococci isolated from cheese whey derived from different animal sources: High biotechnological potential as starter cultures. LWT 2020, 131, 109808. [Google Scholar] [CrossRef]
- Moreno, M.R.F.; Sarantinopoulos, P.; Tsakalidou, E.; De Vuyst, L. The role and application of enterococci in food and health. Int. J. Food Microbiol. 2006, 106, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Giraffa, G. Functionality of enterococci in dairy products. Int. J. Food Microbiol. 2003, 88, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Khusro, A.; Aarti, C. Metabolic heterogeneity and techno-functional attributes of fermented foods-associated coagulase-negative staphylococci. Food Microbiol. 2022, 105, 104028. [Google Scholar] [CrossRef]
- Cavanagh, D.; Fitzgerald, G.F.; McAuliffe, O. From field to fermentation: The origins of Lactococcus lactis and its domestication to the dairy environment. Food Microbiol. 2015, 47, 45–61. [Google Scholar] [CrossRef]
- Mills, S.; Ross, R.P.; Coffey, A. Lactic Acid Bacteria|Lactococcus lactis. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 132–137. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Bachmann, H.; van Pelt-KleinJan, E.; Douwenga, S.; Smid, E.J.; Teusink, B.; van Mastrigt, O. Lifestyle, metabolism and environmental adaptation in Lactococcus lactis. FEMS Microbiol. Rev. 2020, 44, 804–820. [Google Scholar] [CrossRef]
- Aquilanti, L.; Babini, V.; Santarelli, S.; Osimani, A.; Petruzzelli, A.; Clementi, F. Bacterial dynamics in a raw cow’s milk Caciotta cheese manufactured with aqueous extract of Cynara cardunculus dried flowers. Lett. Appl. Microbiol. 2011, 52, 651–659. [Google Scholar] [CrossRef]
- Cardinali, F.; Osimani, A.; Taccari, M.; Milanović, V.; Garofalo, C.; Clementi, F.; Polverigiani, S.; Zitti, S.; Raffaelli, N.; Mozzon, M.; et al. Impact of thistle rennet from Carlina acanthifolia All. subsp. acanthifolia on bacterial diversity and dynamics of a specialty Italian raw ewes’ milk cheese. Int. J. Food Microbiol. 2017, 255, 7–16. [Google Scholar] [CrossRef]
- Naranjo, D.R.; Callanan, M.; Thierry, A.; McAuliffe, O. Superior esterolytic activity in environmental Lactococcus lactis strains is linked to the presence of the SGNH hydrolase family of esterases. JDS Commun. 2020, 1, 25–28. [Google Scholar] [CrossRef]
- Regado, M.A.; Cristóvão, B.M.; Tavaria, F.K.; Ferreira, J.P.; Moutinho, C.G.; Balcão, V.M.; Malcata, F.X. Changes in the pool of free fatty acids in ovine, bovine and caprine milk fats, effected by viable cells and cell-free extracts of Lactococcus lactis and Debaryomyces vanrijiae. Food Chem. 2007, 103, 1112–1118. [Google Scholar] [CrossRef]
- Hemme, D.; Foucaud-Scheunemann, C. Leuconostoc, characteristics, use in dairy technology and prospects in functional foods. Int. Dairy J. 2004, 14, 467–494. [Google Scholar] [CrossRef]
- Belleggia, L.; Ferrocino, I.; Reale, A.; Haouet, M.N.; Corvaglia, M.R.; Milanović, V.; Boscaino, F.; Di Renzo, T.; Di Bella, S.; Borghi, M.; et al. Unravelling microbial populations and volatile organic compounds of artisan fermented liver sausages manufactured in Central Italy. Food Res. Int. 2022, 154, 111019. [Google Scholar] [CrossRef] [PubMed]
- Pogačić, T.; Maillard, M.-B.; Leclerc, A.; Hervé, C.; Chuat, V.; Yee, A.L.; Valence, F.; Thierry, A. A methodological approach to screen diverse cheese-related bacteria for their ability to produce aroma compounds. Food Microbiol. 2015, 46, 145–153. [Google Scholar] [CrossRef]
- Rocha, P.A.B.; Marques, J.M.M.; Barreto, A.S.; Semedo-Lemsaddek, T. Enterococcus spp. from Azeitão and Nisa PDO-cheeses: Surveillance for antimicrobial drug resistance. LWT 2022, 154, 112622. [Google Scholar] [CrossRef]
- Wuyts, S.; Wittouck, S.; De Boeck, I.; Allonsius, C.N.; Pasolli, E.; Segata, N.; Lebeer, S. Large-scale phylogenomics of the Lactobacillus casei group highlights taxonomic inconsistencies and reveals novel clade-associated features. mSystems 2017, 2, e00061-17. [Google Scholar] [CrossRef] [Green Version]
- Hill, D.; Sugrue, I.; Tobin, C.; Hill, C.; Stanton, C.; Ross, R.P. The Lactobacillus casei group: History and health related applications. Front. Microbiol. 2018, 9, 2107. [Google Scholar] [CrossRef] [Green Version]
- Cebrián, R.; Baños, A.; Valdivia, E.; Pérez-Pulido, R.; Martínez-Bueno, M.; Maqueda, M. Characterization of functional, safety, and probiotic properties of Enterococcus faecalis UGRA10, a new AS-48-producer strain. Food Microbiol. 2012, 30, 59–67. [Google Scholar] [CrossRef]
- Jang, E.; Ryu, B.H.; Ju, H.; Kim, T.D. Identification, characterization, and application of a virulence factor (EfEstA) from Enterococcus faecalis. Bioresour. Technol. 2013, 143, 691–694. [Google Scholar] [CrossRef]
- Hausmann, K.; Allen, R.D. Chapter 7—Electron Microscopy of Paramecium (Ciliata). In Methods in Cell Biology; Müller-Reichert, T., Ed.; Academic Press: Cambridge, MA, USA, 2010; Volume 96, pp. 143–173. [Google Scholar] [CrossRef]
- Serio, A.; Chaves-López, C.; Paparella, A.; Suzzi, G. Evaluation of metabolic activities of enterococci isolated from Pecorino Abruzzese cheese. Int. Dairy J. 2010, 20, 459–464. [Google Scholar] [CrossRef]
- Arizcun, C.; Barcina, Y.; Torre, P. Identification and characterization of proteolytic activity of Enterococcus spp. isolated from milk and Roncal and Idiazábal cheese. Int. J. Food Microbiol. 1997, 38, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Tsanasidou, C.; Asimakoula, S.; Sameli, N.; Fanitsios, C.; Vandera, E.; Bosnea, L.; Koukkou, A.-I.; Samelis, J. Safety evaluation, biogenic amine formation, and enzymatic activity profiles of autochthonous enterocin-producing Greek cheese isolates of the Enterococcus faecium/durans group. Microorganisms 2021, 9, 777. [Google Scholar] [CrossRef] [PubMed]
- Domingos-Lopes, M.F.P.; Stanton, C.; Ross, P.R.; Dapkevicius, M.L.E.; Silva, C.C.G. Genetic diversity, safety and technological characterization of lactic acid bacteria isolated from artisanal Pico cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Caro, I.; Rodríguez-Aparicio, L.B.; Rúa, J.; Ferrero, M.A.; García-Armesto, M.R. Characterization of certain bacterial strains for potential use as starter or probiotic cultures in dairy products. J. Food Prot. 2011, 74, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Kolev, P.; Rocha-Mendoza, D.; Ruiz-Ramírez, S.; Ortega-Anaya, J.; Jiménez-Flores, R.; García-Cano, I. Screening and characterization of β-galactosidase activity in lactic acid bacteria for the valorization of acid whey. JDS Commun. 2022, 3, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Belkheir, K.; Zadi-Karam, H.; Karam, N.-E.; Carballo, J.; Centeno, J.A. Effects of selected mesophilic Lactobacillus strains obtained from camel milk on the volatile and sensory profiles of a model short-ripened pressed cows’ milk cheese. Int. Dairy J. 2020, 109, 104738. [Google Scholar] [CrossRef]
- Son, S.-H.; Jeon, H.-L.; Yang, S.-J.; Lee, N.-K.; Paik, H.-D. In vitro characterization of Lactobacillus brevis KU15006, an isolate from kimchi, reveals anti-adhesion activity against foodborne pathogens and antidiabetic properties. Microb. Pathog. 2017, 112, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Song, M.W.; Chung, Y.; Kim, K.-T.; Hong, W.S.; Chang, H.J.; Paik, H.-D. Probiotic characteristics of Lactobacillus brevis B13-2 isolated from kimchi and investigation of antioxidant and immune-modulating abilities of its heat-killed cells. LWT 2020, 128, 109452. [Google Scholar] [CrossRef]
- Okoth, R.A.; Matofari, J.W.; Nduko, J.M. Effectiveness of Levilactobacillus brevis fermentation on antinutrients and protein quality of leaves of selected cassava varieties. Appl. Food Res. 2022, 2, 100134. [Google Scholar] [CrossRef]
- Bhatia, S.; Singh, A.; Batra, N.; Singh, J. Microbial production and biotechnological applications of α-galactosidase. Int. J. Biol. Macromol. 2020, 150, 1294–1313. [Google Scholar] [CrossRef]
- Corbo, M.R.; Albenzio, M.; De Angelis, M.; Sevi, A.; Gobbetti, M. Microbiological and biochemical properties of Canestrato Pugliese hard cheese supplemented with bifidobacteria. J. Dairy Sci. 2001, 84, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Choi, Y.; Yoon, Y. Lactic acid bacteria in kimchi might be a cause for carcinogen production in intestine. Food Control 2021, 126, 108045. [Google Scholar] [CrossRef]
- De Cort, S.; Kumara, H.M.C.S.; Verachtert, H. Localization and characterization of α-glucosidase activity in Lactobacillus brevis. Appl. Environ. Microbiol. 1994, 60, 3074–3078. [Google Scholar] [CrossRef] [Green Version]
- Fenster, K.M.; Parkin, K.L.; Steele, J.L. Intracellular esterase from Lactobacillus casei LILA: Nucleotide sequencing, purification, and characterization. J. Dairy Sci. 2003, 86, 1118–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, M.; Castilho, N.P.A.; Todorov, S.D.; Nero, L.A. Beneficial properties of lactic acid bacteria naturally present in dairy production. BMC Microbiol. 2018, 18, 219. [Google Scholar] [CrossRef] [PubMed]
- Kamarinou, C.S.; Papadopoulou, O.S.; Doulgeraki, A.I.; Tassou, C.C.; Galanis, A.; Chorianopoulos, N.G.; Argyri, A.A. Mapping the key technological and functional characteristics of indigenous lactic acid bacteria isolated from Greek traditional dairy products. Microorganisms 2022, 10, 246. [Google Scholar] [CrossRef]
- Arora, G.; Cormier, F.; Lee, B. Analysis of odor-active volatiles in Cheddar cheese headspace by multidimensional GC/MS/sniffing. J. Agric. Food Chem. 1995, 43, 748–752. [Google Scholar] [CrossRef]
- de Souza, B.M.S.; Borgonovi, T.F.; Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Lactobacillus casei and Lactobacillus fermentum strains isolated from Mozzarella Cheese: Probiotic potential, safety, acidifying kinetic parameters and viability under gastrointestinal tract conditions. Probiotics Antimicrob. Proteins 2019, 11, 382–396. [Google Scholar] [CrossRef]
- Chu, Y.-H.; Yu, X.-X.; Jin, X.; Wang, Y.-T.; Zhao, D.-J.; Zhang, P.; Sun, G.-M.; Zhang, Y.-H. Purification and characterization of alkaline phosphatase from lactic acid bacteria. RSC Adv. 2019, 9, 354–360. [Google Scholar] [CrossRef]
- González, L.; Sacristán, N.; Arenas, R.; Fresno, J.M.; Tornadijo, M.E. Enzymatic activity of lactic acid bacteria (with antimicrobial properties) isolated from a traditional Spanish cheese. Food Microbiol. 2010, 27, 592–597. [Google Scholar] [CrossRef]
- Huang, D.Q.; Prévost, H.; Diviès, C. Principal characteristics of β-galactosidase from Leuconostoc spp. Int. Dairy J. 1995, 5, 29–43. [Google Scholar] [CrossRef]
- Bintsis, T. Yeasts in different types of cheese. AIMS Microbiol. 2021, 7, 447–470. [Google Scholar] [CrossRef] [PubMed]
- Freitas, C.; Malcata, F.X. Microbiology and biochemistry of cheeses with Appélation d’Origine Protegée and manufactured in the Iberian Peninsula from ovine and caprine milks. J. Dairy Sci. 2000, 83, 584–602. [Google Scholar] [CrossRef] [Green Version]
- Padilla, B.; Manzanares, P.; Belloch, C. Yeast species and genetic heterogeneity within Debaryomyces hansenii along the ripening process of traditional ewes’ and goats’ cheeses. Food Microbiol. 2014, 38, 160–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorordo, M.F.; Lucca, M.E.; Sangorrín, M.P. Biocontrol efficacy of the Vishniacozyma victoriae in semi-commercial assays for the control of postharvest fungal diseases of organic pears. Curr. Microbiol. 2022, 79, 259. [Google Scholar] [CrossRef]
- Schmidt, S.K.; Vimercati, L.; Darcy, J.L.; Arán, P.; Gendron, E.M.S.; Solon, A.J.; Porazinska, D.; Dorador, C. A Naganishia in high places: Functioning populations or dormant cells from the atmosphere? Mycology 2017, 8, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Jacques, N.; Mallet, S.; Laaghouiti, F.; Tinsley, C.R.; Casaregola, S. Specific populations of the yeast Geotrichum candidum revealed by molecular typing. Yeast 2017, 34, 165–178. [Google Scholar] [CrossRef]
- Grygier, A.; Myszka, K.; Juzwa, W.; Białas, W.; Rudzińska, M. Galactomyces geotrichum mold isolated from a traditional fried cottage cheese produced omega-3 fatty acids. Int. J. Food Microbiol. 2020, 319, 108503. [Google Scholar] [CrossRef]
- Hipólito, A.; Silva, R.A.A.d.; Caretta, T.d.O.; Silveira, V.A.I.; Amador, I.R.; Panagio, L.A.; Borsato, D.; Celligoi, M.A.P.C. Evaluation of the antifungal activity of sophorolipids from Starmerella bombicola against food spoilage fungi. Biocatal. Agric. Biotechnol. 2020, 29, 101797. [Google Scholar] [CrossRef]
- Papademas, P.; Robinson, R.K. A comparison of the chemical, microbiological and sensory characteristics of bovine and ovine Halloumi cheese. Int. Dairy J. 2000, 10, 761–768. [Google Scholar] [CrossRef]
- Delgado, F.J.; González-Crespo, J.; Cava, R.; Ramírez, R. Formation of the aroma of a raw goat milk cheese during maturation analysed by SPME–GC–MS. Food Chem. 2011, 129, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Dahl, S.; Tavaria, F.K.; Malcata, F.X. Relationships between flavour and microbiological profiles in Serra da Estrela cheese throughout ripening. Int. Dairy J. 2000, 10, 255–262. [Google Scholar] [CrossRef]
- Mcsweeney, P.L.H.; Sousa, M.J. Biochemical pathways for the production of flavour compounds in cheeses during ripening: A review. Lait 2000, 80, 293–324. [Google Scholar] [CrossRef]
- Collins, Y.F.; McSweeney, P.L.H.; Wilkinson, M.G. Lipolysis and free fatty acid catabolism in cheese: A review of current knowledge. Int. Dairy J. 2003, 13, 841–866. [Google Scholar] [CrossRef]
- Zheng, X.; Shi, X.; Wang, B. A Review on the general cheese processing technology, flavor biochemical pathways and the influence of yeasts in cheese. Front. Microbiol. 2021, 12, 703284. [Google Scholar] [CrossRef] [PubMed]
- Tavaria, F.K.; Tavares, T.G.; Silva-Ferreira, A.C.; Malcata, F.X. Contribution of coagulant and native microflora to the volatile-free fatty acid profile of an artisanal cheese. Int. Dairy J. 2006, 16, 886–894. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cheese Source | pH | Total Titratable Acidity (mL of 0.1N NaOH) | Acetic Acid (g 100 g−1) | Lactic Acid (g 100 g−1) | Water Activity | Specific Volume (g mL−1) |
|---|---|---|---|---|---|---|
| Producer 1 | 5.83 ± 0.06 a | 7.52 ± 1.44 c | 0.11 ± 0.04 a | 0.63 ± 0.09 b | 0.940 ± 0.011 b | 1.11 ± 0.14 a |
| Producer 2 | 5.71 ± 0.18 a | 9.82 ± 2.90 b | 0.12 ± 0.03 a | 1.01 ± 0.09 a | 0.975 ± 0.003 a | 1.09 ± 0.05 ab |
| Producer 3 | 5.54 ± 0.09 b | 13.97 ± 0.41 a | 0.27 ± 0.09 a | 1.13 ± 0.11 a | 0.969 ± 0.004 a | 1.07 ± 0.11 ab |
| Producer 4 | 5.46 ± 0.09 b | 11.80 ± 1.74 ab | 0.26 ± 0.05 a | 1.08 ± 0.04 a | 0.963 ± 0.008 a | 0.98 ± 0.06 b |
| Cheese Source | L* | a* | b* | Chroma | Hue |
|---|---|---|---|---|---|
| Producer 1 | 106.6 ± 3.6 c | 0.35 ± 0.77 a | 40.54 ± 1.47 a | 40.55 ± 1.47 a | 89.58 ± 1.07 b |
| Producer 2 | 117.8 ± 1.7 b | −0.90 ± 0.21 bc | 34.00 ± 1.23 b | 34.01 ± 1.23 b | 91.43 ± 0.3 a |
| Producer 3 | 116.0 ± 1.9 ab | −1.86 ± 0.67 c | 39.42 ± 0.97 a | 39.46 ± 0.99 a | 92.65 ± 0.93 a |
| Producer 4 | 113.3 ± 1.8 a | 0.20 ± 1.04 ab | 39.24 ± 2.16 a | 39.25 ± 2.15 a | 89.63 ± 1.48 b |
| Cheese Source | Hardness | Cohesiveness | Springiness | Chewiness | Resilience |
|---|---|---|---|---|---|
| Producer 1 | 53.36 ± 15.67 a | 0.22 ± 0.04 b | 0.48 ± 0.18 a | 6.79 ± 5.83 a | 0.16 ± 0.03 a |
| Producer 2 | 7.99 ± 1.05 b | 0.30 ± 0.07 a | 0.28 ± 0.07 b | 0.65 ± 0.12 b | 0.18 ± 0.01 a |
| Producer 3 | 15.30 ± 1.52 b | 0.25 ± 0.03 ab | 0.26 ± 0.05 b | 0.95 ± 0.19 b | 0.16 ± 0.03 a |
| Producer 4 | 11.89 ± 0.94 b | 0.26 ± 0.04 ab | 0.28 ± 0.04 b | 0.83 ± 0.09 b | 0.17 ± 0.02 a |
| Cheese Source | Presumptive Lactococci | Presumptive Thermophilic Cocci | Presumptive Lactobacilli | Coagulase-Negative Cocci | Enterococci | Yeasts | Molds |
|---|---|---|---|---|---|---|---|
| Producer 1 | 8.24 ± 0.16 ab | 7.16 ± 0.36 b | 8.40 ± 0.15 a | 5.34 ± 0.21 b | 6.81 ± 0.22 bc | 4.24 ± 0.75 a | n.d. |
| Producer 2 | 8.66 ± 0.35 a | 7.95 ± 0.45 a | 8.50 ± 0.35 a | 5.91 ± 0.46 b | 7.57 ± 0.11 ab | 3.23 ± 0.32 a | 4.07 ± 0.58 a |
| Producer 3 | 7.98 ± 0.20 b | 8.28 ± 0.17 a | 8.45 ± 0.09 a | 5.44 ± 0.10 b | 6.56 ± 0.12 c | 1.27 ± 1.47 b | n.d. |
| Producer 4 | 8.41 ± 0.21 ab | 8.26 ± 0.31 a | 8.13 ± 0.20 a | 6.42 ± 0.22 a | 7.84 ± 0.73 a | 2.51 ± 0.70 ab | n.d. |
| Isolation Source | Isolate Code | Closest Relative | % Identity * | Accession Number ** |
|---|---|---|---|---|
| Producer 1 | SE1 | Enterococcus durans | 98.33% | NR_036922 |
| SE2 | Levilactobacillus brevis | 98.30% | NR_116238 | |
| SE3 | Levilactobacillus brevis | 99.25% | NR_042438 | |
| SE4 | Latilactobacillus graminis | 97.80% | NR_113922 | |
| SE5 | Enterococcus durans | 98.65% | NR_036922 | |
| SE6 | Enterococcus faecium | 99.30% | NR_114742 | |
| SE7 | Enterococcus faecalis | 99.82% | NR_113902 | |
| Producer 2 | SE8 | Lacticaseibacillus casei group | 98.66% | NR_025880 |
| SE9 | Lacticaseibacillus casei group | 97.89% | NR_025880 | |
| SE10 | Lacticaseibacillus casei group | 99.27% | NR_117987 | |
| SE11 | Levilactobacillus brevis | 98.99% | NR_116238 | |
| SE12 | Enterococcus faecalis | 98.82% | NR_113902 | |
| SE13 | Enterococcus faecalis | 98.96% | NR_113902 | |
| SE14 | Enterococcus durans | 99.64% | NR_036922 | |
| SE15 | Enterococcus lactis | 99.34% | NR_117562 | |
| SE16 | Enterococcus durans | 99.35% | NR_036922 | |
| Producer 3 | SE17 | Lacticaseibacillus casei group | 99.42% | NR_025880 |
| SE18 | Levilactobacillus brevis | 99.23% | NR_116238 | |
| SE19 | Levilactobacillus brevis | 99.61% | NR_116238 | |
| SE20 | Levilactobacillus brevis | 99.12% | NR_116238 | |
| SE21 | Leuconostoc mesenteroides subsp. dextranicum | 99.23% | NR_113911 | |
| SE22 | Enterococcus faecalis | 98.86% | NR_113902 | |
| Producer 4 | SE23 | Latilactobacillus graminis | 98.81% | NR_042438 |
| SE24 | Levilactobacillus brevis | 99.40% | NR_116238 | |
| SE25 | Lacticaseibacillus casei group | 97.80% | NR_025880 | |
| SE26 | Latilactobacillus graminis | 99.28% | NR_042438 | |
| SE27 | Leuconostoc mesenteroides subsp. dextranicum | 99.90% | NR_113911 | |
| SE28 | Enterococcus durans | 97.45% | NR_036922 | |
| SE29 | Enterococcus durans | 98.72% | NR_036922 | |
| SE30 | Lacticaseibacillus casei group | 99.72% | NR_025880 |
| Isolation Source | Isolate Code | Isolate | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Producer 1 | SE1 | Enterococcus durans | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 0 | 0 | 0 | 0 | 0 |
| SE2 | Levilactobacillus brevis | 0 | 0 | 1 | 0 | 0 | 3 | 3 | 0 | 0 | 0 | 2 | 1 | 2 | 5 | 0 | 2 | 5 | |
| SE3 | Levilactobacillus brevis | 0 | 0 | 2 | 1 | 0 | 4 | 4 | 1 | 0 | 0 | 5 | 3 | 3 | 5 | 3 | 2 | 5 | |
| SE4 | Latilactobacillus graminis | 0 | 0 | 0 | 0 | 0 | 4 | 3 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | |
| SE5 | Enterococcus durans | 0 | 0 | 2 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 4 | 0 | 0 | 1 | |
| SE6 | Enterococcus faecium | 0 | 0 | 2 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 5 | 0 | 0 | 1 | |
| SE7 | Enterococcus faecalis | 0 | 1 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1 | 0 | 0 | 0 | 3 | 1 | |
| Producer 2 | SE8 | Lacticaseibacillus casei group | 0 | 0 | 3 | 3 | 0 | 4 | 5 | 1 | 0 | 1 | 2 | 4 | 0 | 3 | 0 | 2 | 1 |
| SE9 | Lacticaseibacillus casei group | 0 | 0 | 3 | 2 | 0 | 2 | 4 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 1 | 0 | |
| SE10 | Lacticaseibacillus casei group | 0 | 0 | 2 | 1 | 0 | 4 | 5 | 1 | 0 | 0 | 3 | 4 | 0 | 1 | 0 | 2 | 1 | |
| SE11 | Levilactobacillus brevis | 0 | 0 | 2 | 1 | 0 | 4 | 3 | 1 | 0 | 0 | 5 | 4 | 3 | 5 | 2 | 3 | 5 | |
| SE12 | Enterococcus faecalis | 0 | 1 | 4 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 2 | 2 | 0 | 0 | 0 | 3 | 1 | |
| SE13 | Enterococcus faecalis | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 1 | 0 | 3 | 0 | 2 | 0 | |
| SE14 | Enterococcus durans | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 2 | 5 | 0 | 0 | 1 | |
| SE15 | Enterococcus lactis | 0 | 0 | 2 | 2 | 0 | 2 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 2 | |
| SE16 | Enterococcus durans | 0 | 0 | 2 | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 2 | 5 | 0 | 0 | 2 | |
| Producer 3 | SE17 | Lacticaseibacillus casei group | 0 | 0 | 3 | 3 | 0 | 5 | 5 | 1 | 0 | 1 | 3 | 4 | 0 | 1 | 0 | 4 | 3 |
| SE18 | Levilactobacillus brevis | 0 | 0 | 1 | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 4 | 1 | 2 | 5 | 1 | 1 | 5 | |
| SE19 | Levilactobacillus brevis | 0 | 1 | 2 | 1 | 0 | 5 | 3 | 1 | 0 | 0 | 5 | 5 | 1 | 5 | 3 | 2 | 5 | |
| SE20 | Levilactobacillus brevis | 0 | 0 | 1 | 0 | 0 | 4 | 1 | 0 | 0 | 0 | 4 | 1 | 2 | 5 | 0 | 3 | 4 | |
| SE21 | Leuconostoc mesenteroides | 0 | 0 | 3 | 1 | 0 | 2 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 3 | 0 | 0 | 1 | |
| SE22 | Enterococcus faecalis | 0 | 0 | 2 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 3 | 0 | 0 | 0 | 2 | 1 | |
| Producer 4 | SE23 | Latilactobacillus graminis | 0 | 3 | 0 | 0 | 0 | 5 | 4 | j2 | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 0 |
| SE24 | Levilactobacillus brevis | 0 | 0 | 2 | 1 | 0 | 5 | 4 | 3 | 0 | 0 | 5 | 5 | 5 | 5 | 4 | 4 | 5 | |
| SE25 | Lacticaseibacillus casei group | 0 | 2 | 2 | 2 | 0 | 5 | 4 | 3 | 0 | 2 | 3 | 5 | 0 | 4 | 0 | 4 | 2 | |
| SE26 | Latilactobacillus graminis | 0 | 1 | 0 | 0 | 0 | 4 | 4 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | |
| SE27 | Leuconostoc mesenteroides | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 5 | 5 | 0 | 1 | 5 | |
| SE28 | Enterococcus durans | 0 | 0 | 1 | 3 | 0 | 4 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 3 | 0 | 0 | 4 | |
| SE29 | Enterococcus durans | 0 | 0 | 1 | 3 | 0 | 3 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 3 | |
| SE30 | Lacticaseibacillus casei group | 0 | 0 | 1 | 3 | 0 | 5 | 3 | 1 | 0 | 0 | 2 | 2 | 0 | 3 | 0 | 1 | 2 |
| Compound | CAS-NUMBER | Category | Flavour Note | Producer 1 | Producer 2 | Producer 3 | Producer 4 |
|---|---|---|---|---|---|---|---|
| ethanol | 64-17-5 | Alcohol | Alcohol, mild | 4723 ± 1168 | 12892 ± 4021 | 7485 ± 1964 | 8199 ± 4395 |
| propan-2-ol | 67-63-0 | Alcohol | rubbing alcohol | 844 ± 47 | 4963 ± 5197 | 1350 ± 648 | 1005 ± 476 |
| 2-butanone | 78-93-3 | Ketone | Acetone, etheric | 22638 ± 12768 | 11257 ± 721 | 15085 ± 4830 | 19462 ± 3181 |
| butanal, 3-methyl | 590–86-3 | Aldehyde | Dark chocolate, malt, green | 263 ± 227 | 62 ± 14 | 1257 ± 223 | 385 ± 514 |
| isobutyl acetate | 110-19-0 | Ester | fruity, floral banana-like | 76 ± 11 | 79 ± 78 | 53 ± 12 | 164 ± 112 |
| 3-hydroxybutan-2-one | 5077-67-8 | Ketone | Buttery | 2067 ± 1026 | 2652 ± 1947 | 193 ± 253 | 541 ± 218 |
| 1-butanol, 3-methyl | 123-51-3 | Alcohol | Fruity, alcohol | 21 ± 18 | 49 ± 65 | 31 ± 31 | 34 ± 13 |
| propanoic acid, 2-methyl (isobutytic acid) | 79–31-2 | Acid | rancid butter | 26 ± 6 | 32 ± 65 | 114 ± 87 | 60 ± 43 |
| butanoic acid | 107–92-6 | Acid | Rancid, cheesy, putrid, sweaty | 3135 ± 1923 | 2507 ± 2459 | 9613 ± 2312 | 10872 ± 4345 |
| butanoic acid, 3-methyl (isovaleric acid) | 503–74-2 | Acid | Swiss, cheese, waxy, sweaty, | 100 ± 57 | 176 ± 71 | 88 ± 66 | 97 ± 45 |
| butanoic acid, 2-methyl | 116–53-0 | Acid | Fruity, sour, sweaty | 109 ± 142 | 314 ± 5 | 1105 ± 644 | 719 ± 850 |
| 2-heptanone | 110–43-0 | Ketone | Floral, fruity | 163 ± 55 | 60 ± 63 | 321 ± 104 | 153 ± 140 |
| butanoic acid, 1-methylpropyl ester | 819-97-6 | Ester | 38 ± 35 | 13 ± 11 | 563 ± 711 | 70 ± 60 | |
| benzaldehyde | 100-52-7 | Aldehyde | sweet, strong almond odour | 184 ± 55 | 125 ± 106 | 127 ± 16 | 82 ± 42 |
| ethyl hexanoate | 123-66-0 | Ester | Pineapple, apple powerful | 11139 ± 8817 | 14680 ± 1815 | 60558 ± 24699 | 51008 ± 29977 |
| hexanoic acid | 142-62-1 | Acid | Pungent, blue cheese, goat-like | 227 ± 95 | 213 ± 157 | 246 ± 46 | 509 ± 262 |
| phenylacetaldehyde | 122-78-1 | Aldehyde | pungent green floral and sweet odour of hyacinth type | 182 ± 48 | 134 ± 62 | 215 ± 175 | 131 ± 129 |
| heptanoic acid | 111-14-8 | Acid | rancid, sour, fatty odour | 39 ± 11 | 50 ± 6 | 42 ± 5 | 55 ± 48 |
| 2-nonanone | 821-55-6 | Ketone | Musty, fruity, floral | 188 ± 25 | 126 ± 15 | 184 ± 41 | 197 ± 63 |
| octanoic acid | 124-07-2 | Acid | Goaty, waxy, soapy, rancid | 9636 ± 7581 | 9715 ± 411 | 23449 ± 4690 | 18098 ± 8359 |
| octanoic acid ethyl ester | 106-32-1 | Ester | 1480 ± 987 | 3070 ± 1417 | 13440 ± 16802 | 6082 ± 5108 | |
| dodecane | 112-40-3 | Alkane | 118 ± 59 | 169 ± 204 | 557 ± 581 | 251 ± 102 | |
| 1,3-di-tert-butylbenzene | 1014-60-4 | Aromatic hydrocarbon | 68 ± 59 | 367 ± 445 | 199 ± 139 | 261 ± 275 | |
| nonanoic acid | 112-05-0 | Acid | Coconut, fatty odour | 63 ± 34 | 72 ± 30 | 304 ± 355 | 53 ± 48 |
| decanoic acid | 334-48-5 | Acid | Rancid | 3825 ± 2930 | 2184 ± 529 | 11532 ± 10468 | 17946 ± 17472 |
| decanoic acid ethyl ester | 110-38-3 | Ester | 1161 ± 664 | 1480 ± 496 | 5899 ± 7057 | 2118 ± 1010 | |
| dodecanal | 112-54-9 | Aldehyde | fatty odour | 21 ± 4 | 19 ± 12 | 12 ± 4 | 40 ± 30 |
| dodecanoic acid | 143-07-7 | Acid | like oil of bay | 121 ± 98 | 45 ± 6 | 81 ± 45 | 191 ± 84 |
| dodecanoic acid ethyl ester | 106-33-2 | Ester | fruity, floral | 69 ± 33 | 149 ± 101 | 257 ± 288 | 136 ± 91 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rampanti, G.; Ferrocino, I.; Harasym, J.; Foligni, R.; Cardinali, F.; Orkusz, A.; Milanović, V.; Franciosa, I.; Garofalo, C.; Mannozzi, C.; et al. Queijo Serra da Estrela PDO Cheese: Investigation into Its Morpho-Textural Traits, Microbiota, and Volatilome. Foods 2023, 12, 169. https://doi.org/10.3390/foods12010169
Rampanti G, Ferrocino I, Harasym J, Foligni R, Cardinali F, Orkusz A, Milanović V, Franciosa I, Garofalo C, Mannozzi C, et al. Queijo Serra da Estrela PDO Cheese: Investigation into Its Morpho-Textural Traits, Microbiota, and Volatilome. Foods. 2023; 12(1):169. https://doi.org/10.3390/foods12010169
Chicago/Turabian StyleRampanti, Giorgia, Ilario Ferrocino, Joanna Harasym, Roberta Foligni, Federica Cardinali, Agnieszka Orkusz, Vesna Milanović, Irene Franciosa, Cristiana Garofalo, Cinzia Mannozzi, and et al. 2023. "Queijo Serra da Estrela PDO Cheese: Investigation into Its Morpho-Textural Traits, Microbiota, and Volatilome" Foods 12, no. 1: 169. https://doi.org/10.3390/foods12010169