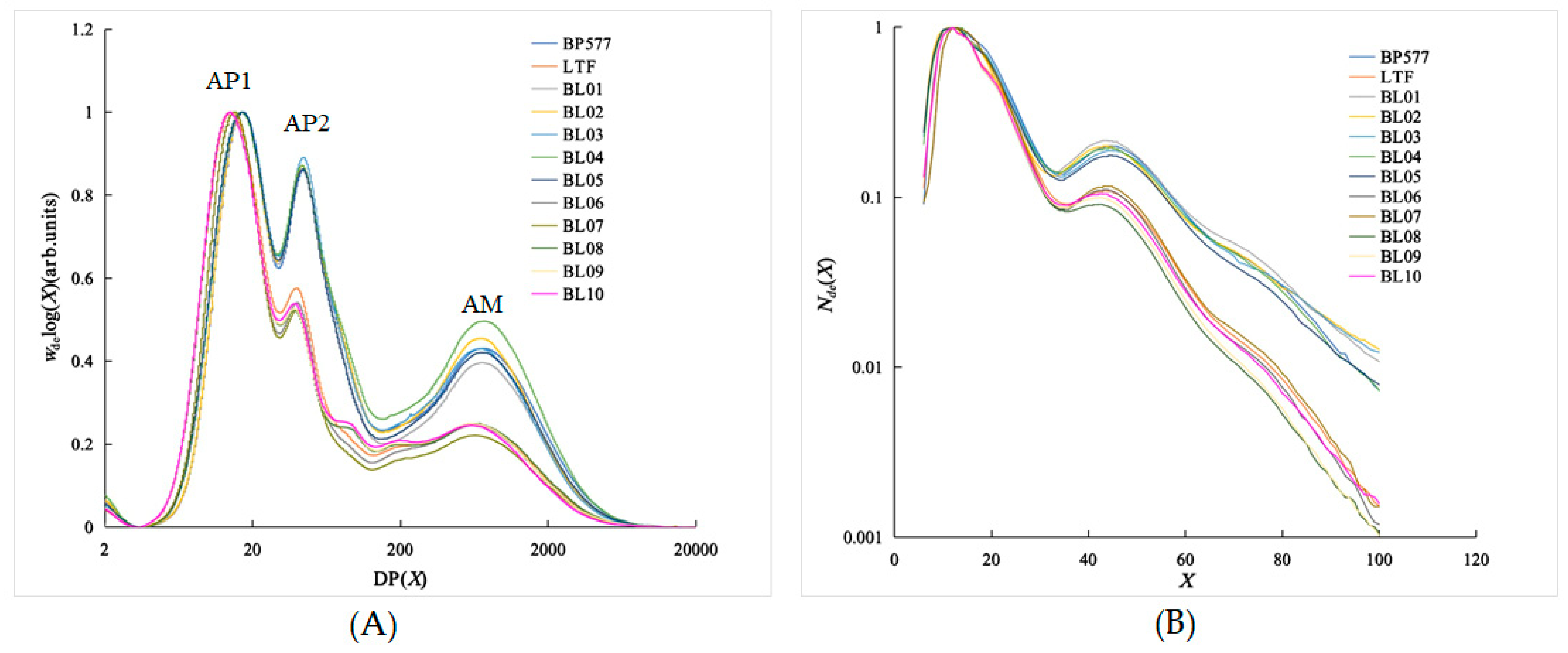
3.2. SEC Chain-Length Distributions of the Debranched Starch
The SEC weight distribution of debranched starch from the GC/
BEIIb, TT/
BEIIb, GC/
be2b, and TT/
be2b series is shown in
Figure 2A. The maximum SEC weight distribution of each sample is normalized to an arbitrary value of 1. Three peaks exist in the SEC-weight CLDs (
Figure 2A). Debranched amylopectin has two peaks: the first peak at DP from 14 to 17, or
XAP1, indicates short amylopectin chains, while the second peak at DP from 38 to 44, or
XAP2, indicates a long amylopectin chain (
Table 1;
Figure 2A). Debranched amylose has only one peak at DP 580 (LTFB) to 729 (BP577). Two parents showed distinctive CLD parameters, and most of the parameters of their offspring were between two parents with some transgressive segregations that had larger or smaller values than the parents (
Table 1). The parent BP577 and the breeding lines with the
be2b allele mostly had larger
XAP1, XAP2, and
XAM, indicating that their amylopectin and amylose had larger molecular sizes than those carrying the
BEIIb allele. The lines with
SSIIa-GC/
BEIIb had slightly larger
XAP1 and
XAP2 than those with
SSIIa-TT/
BEIIb. Rice with the
be2b allele had much higher
hAP2/hAP1 than those with the
BEIIb allele, which is in agreement with the result of Tappiban, et al. [
19], who showed that the mutant deficiency in
BEIIb had the
hAP2/hAP1 of 0.947. The value of
hAP2/hAP1 of the
be2b genotypes was as high as that of potato starches [
35], and a little larger than that of cassava starches and common rice starches [
23,
36].
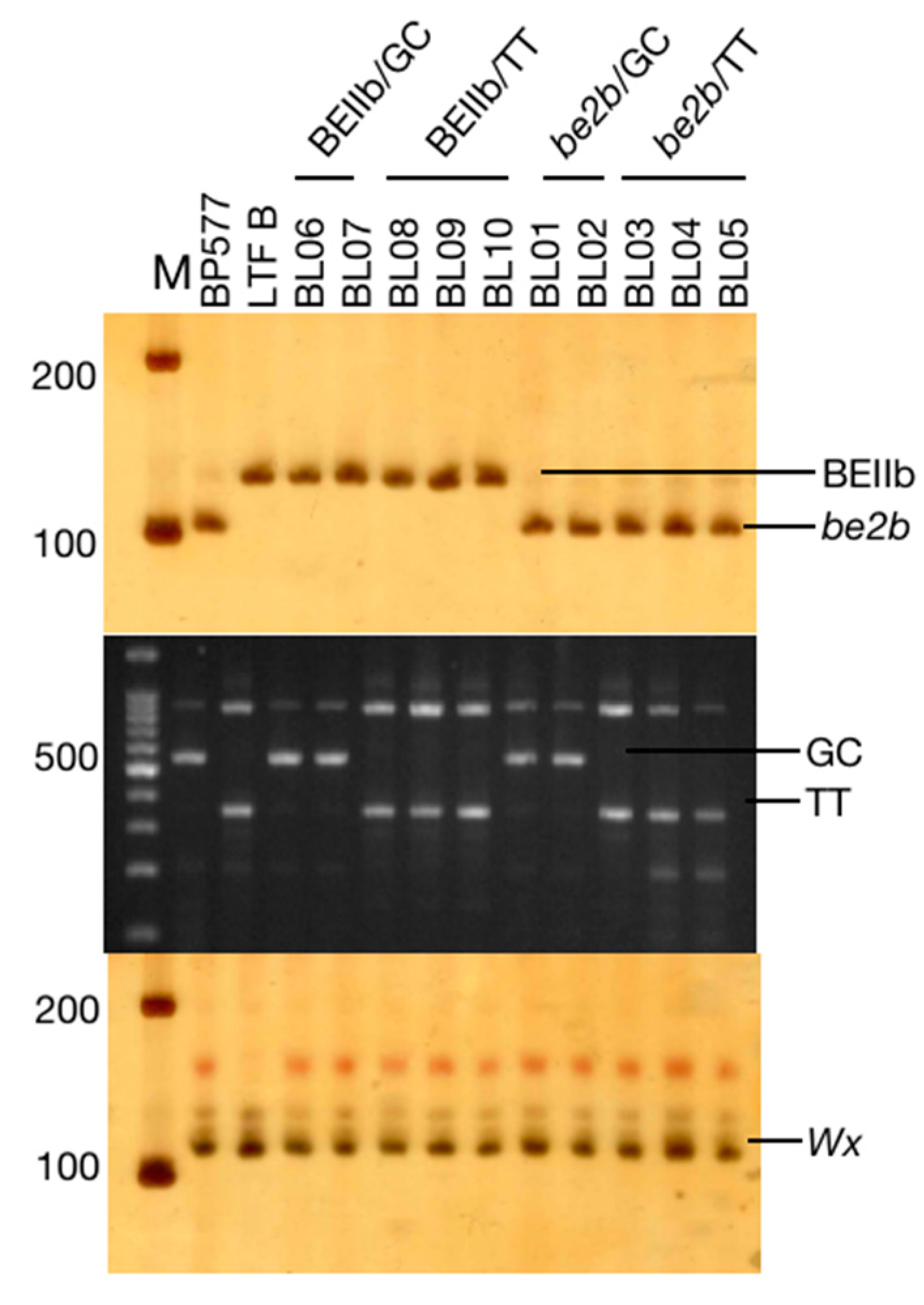
Both parents carry the same
Wxa allele with high AC, so all the breeding lines had AC larger than 27%. The AC of the rice with the
be2b allele (32.27%~34.84%) was significantly higher than those with the
BEIIb allele (26.82%~28.26%). However, there is no significant difference between the
SSIIa GC and TT alleles (
Table 1). In previous reports, the
be2b allele represents the lack of BEIIb and can increase the AC of rice starches [
19,
37,
38,
39] and corn starches [
40]. Since GBSS is responsible for amylose synthesis, these results may suggest that the GBSS might be inhibited by an active BEIIb.
The CLD number of the debranched starch from FACE is shown in
Figure 2B, and the maximum value is normalized to 1. The maximum peak is at DP ~ 12, and a small bump is found at DP30~40. The amylopectin chains can be divided into four groups according to DP: DP ≤ 12 (A chain), 13 ≤ DP ≤ 24 (B1 chain), 25 ≤ DP ≤ 36 (B2 chain), and DP ≥ 37 (B3 and ultra-long chain) [
31]. The proportion of amylopectin CLD showed a significant difference between starches from different
BEIIb and
SSIIa genotypes (
Table 2), which was in agreement with the results from the SEC weight distribution (
Table 1 and
Figure 2A). The proportion of A chain and B1 chain of starches with the
be2b genotype ranged from 20.29% to 21.73% and 41.74% to 44.06%, which was much lower than those with the
BEIIb genotype. Similarly, the proportions of the B2 and B3 chains of
be2b were much higher than those of the
BEIIb allele, leading to the average length of the
be2b starch amylopectin being significantly higher than that of its
BEIIb counterpart. The structural changes derived from BEIIb deficiency were in agreement with previous reports [
18,
21,
22,
26]. These results confirmed that the
ae mutant not only prolongs the branch chain length of amylopectin but also significantly increases amylose content in starch [
37]. Under the same
BEIIb allele background, the A chain content of the SSIIa-GC starch was significantly lower, while the B1 chain content was significantly higher than that of SSIIa-TT (
Table 2). In common rice accessions, it was proven some time ago that rice with the GC allele has lower fa chain content and fa/fb1 ratio than that with the TT allele [
14]. However, under the
be2b allele background, rice materials with SSIIa-GC starch had a little higher A chain content than those with the SSIIa-TT allele, while the reverse was found for the content of the B1 chain (
Table 2).
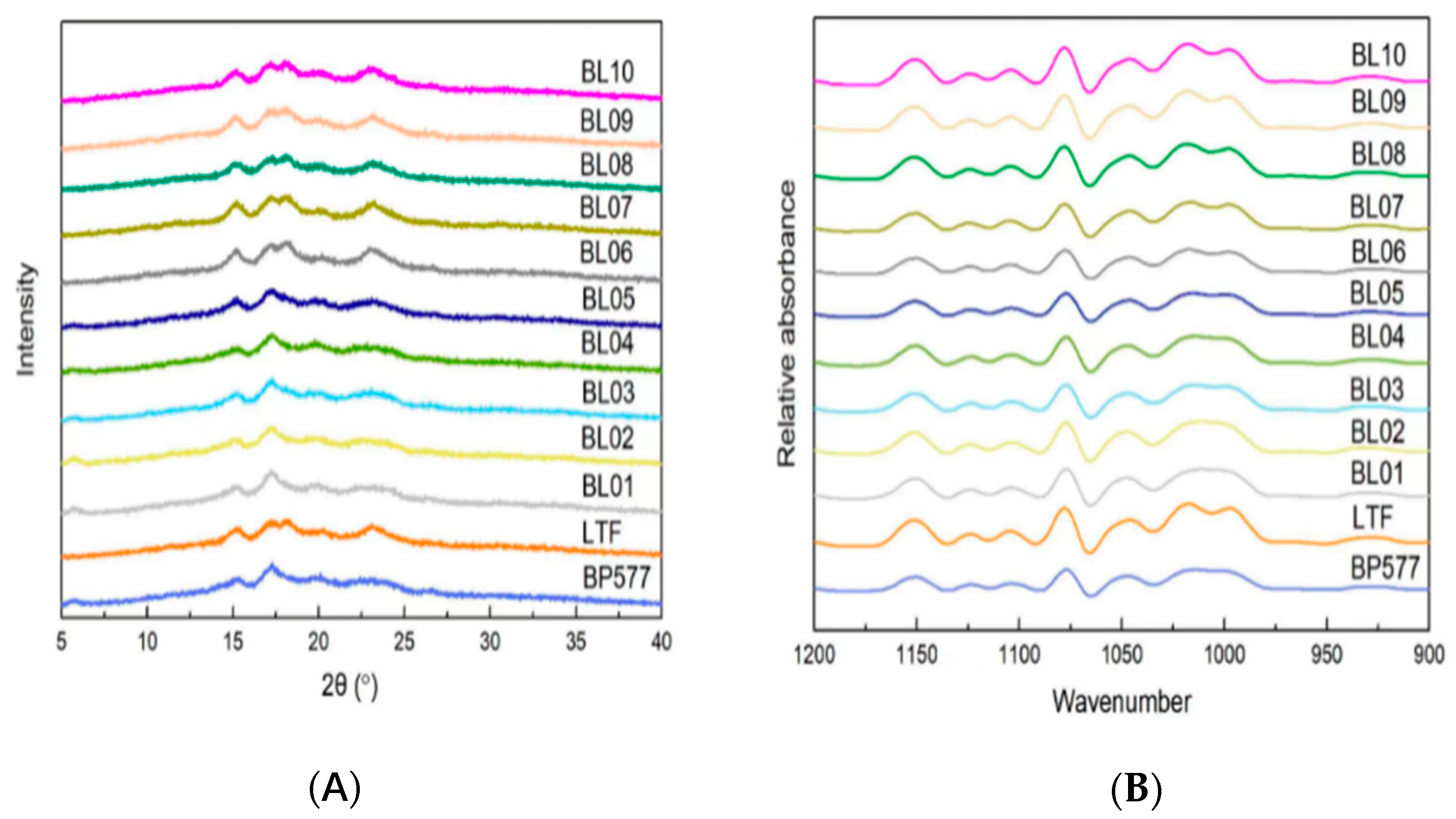
3.3. Crystalline Structure
The XRD patterns of the two parents and 10 breeding lines are shown in
Figure 3A. The A-type crystallinity starch has the characteristic of containing more short branch chains of amylopectin, while the B-type crystallinity has the characteristic of containing more long branch chains, and the C-type starch is a mixture of A and B -type crystallinity. Starches with the
BEIIb allele had a typical A-type crystalline with four peaks at diffraction angles (2
θ) of 15°, 17°, 18°, and 23° in common with those of common rice and other cereal starches [
19,
37]. The BEIIb deficiency mutant starch generally has B-type crystallinity [
19,
22,
24,
37,
40]. In this study, starches with the
be2b allele had a C-type pattern with a strong peak at 17° and weak peaks at 5°, 15°, and 23°, which may be derived from incomplete suppression of the expression of the BEIIb [
37]. The relative crystallinity (RC) showed significant differences between genotypes with different allele combinations (
Table 3). Among starches with A-type crystalline, values of RC with the GC allele (BL06 and BL07) were a little higher than those of the TT allele (BL08-10). Starch with high RC was in alignment with higher B1 chain proportion, so in the common starches, those with the GC allele had higher fb1 content (
Table 2), resulting in high RC. However, among the starches with C-type crystallinity, values of RC with the GC allele (BL01-BL02) were a little lower than those of the TT allele (BL03-05) (
Table 3). It is possible that the short chains generated in the
BEIIb mutant were inhibited from elongation due to the action of the SSIIa isoform. It is also suggested that SSIIa forms a heteromeric protein complex with SSI and BEIIb in cereal endosperm to synthesize amylopectin [
6,
41,
42]. Therefore, the lack of BEIIb isoform may decrease the amount of functional protein complex, which results in less short-chain elongation, even though the SSIIa is abundant.
The 900–1200 cm
−1 region of the FTIR spectra of the two parents and their breeding lines is shown in
Figure 3B. The 1045/1022 cm
−1 ratio displayed significant differences between the rice materials, ranging from 0.634 (BL10) to 0.812 (BL01) (
Table 3). The higher ratio of 1045/1022 cm
−1 may indicate that the starch has a higher degree of ordered structure. In either
BEIIb or
SSIIa allele backgrounds, it is clearly shown that starches with the
be2b or GC allele had a higher degree of ordered structure than their counterparts with the
BEIIb or TT allele. The ratio 1022/995 cm
−1 indicates the proportion of amorphous structure. The starches with the
BEIIb allele had a higher ratio of 1022/995 cm
−1 than those with the
be2b allele, which is not in agreement with the relative crystallinity data (
Table 3).
3.4. Thermal Properties
The onset temperature (T
o), peak temperature (T
p), conclusion temperature (T
c), and the enthalpy of gelatinization (ΔH) of all the rice starches are presented in
Table 4. For common starch with the
BEIIb allele, it is well known that rice accessions with the GC allele had intermediate or high GT, while those with the TT SNP had a low GT [
13,
14,
16]. BL06 and BL07 had a Tp of around 78 °C, and those of BL08-10 had a T
p of around 65 °C (
Table 4). The difference in gelatinization temperature can be easily explained by the difference in amylopectin CLDs since the fa chain content and fa/fb1 ratio are negatively correlated with GT, whereas fb1 chain content is positively correlated with GT [
14]. The ΔH of BL06 and BL07 was the highest among all samples (
Table 4), partially because both these samples also had the highest RC (
Table 3). It is plausible that the higher fb1 chain content in the BL06 and BL07 starches resulted in their highest ΔH and RC (
Table 2). However, among the rice samples with the
be2b allele, none of the thermal properties displayed a significant difference between the GC and TT alleles (
Table 4). However, in japonica rice, Ida et al. [
24] indicated that the GT of the
ss2a/be2b double mutants was higher than that of the WT mutant line but lower than that of the
be2b mutant lines. The
be2b allele or the
ae mutation caused the rice to synthesize a much longer amylopectin chain, leading to a higher gelatinization temperature [
18,
37]. Furthermore,
BEIIb may suppress the expression of
SSIIa to a certain extent, or else, SSIIa function is not so important when BEIIb is defect. Thus, no significant difference between the GC and TT alleles in the
be2b background was found in the
be2b background (
Table 4).
3.6. Relationship between Different SSIIa/BEIIb Genotypes
From the structural and physicochemical properties of the breeding lines derived from the parents BP577 and LTFB with different
SSIIa-GC/TT and
BEIIb/be2b alleles, it is clear that rice lines with the
be2b allele displayed distinct structural and physicochemical properties from those with the
BEIIb allele. However, in the
be2b allele background, most parameters between the GC and TT alleles did not show differences, suggesting that the function of SSIIa is not important in the
be2b allele background. From this aspect, it could be concluded that
BEIIb is more important in determining the structural and physicochemical properties than
SSIIa. By analysis of all the BE mutants, Tappiban, et al. [
19] also indicated that BEIIb played a more important role in determining the structural and physicochemical properties than BEI and BEIIa. To further reveal the relative important functions of the
SSIIa and
BEIIb alleles, clustering analysis was carried out for different genotypes (
Figure 5). As expected, two groups were formed as the breeding lines were derived from two parents, BP577 and LTFB. The two groups were formed according to different
BEIIb alleles but not
SSIIa alleles, further confirming that BEIIb is more important. For the
BEIIb-related group (the upper one), rice materials with different GC/TT were revealed in two subgroups, and the parent LTFB was grouped with the BL08-10, indicating that the group represented the low GT with the TT allele. However, for the
be2b-related group, although two or three subgroups could be revealed, they did not follow the GC/TT groups. Instead, parent BP577, BL01, and BL02 formed a subgroup, BL03 and BL05 formed a second subgroup, and BL04 itself formed a third subgroup (
Figure 5).
3.7. Relationships between Fine Structure and Physicochemical Properties
To explore the structure-function relationships, the correlation analysis between starch structural parameters and physicochemical properties is shown in
Table S2. Most parameters had a significant correlation with each other, except for ΔH, which had no correlation with any other parameters, and breakdown (BD) viscosity, which showed correlations only with some structural parameters. Since there are two types of starch with distinct structural and physicochemical differences, the correlations may also differ from those in previous studies with common starches. The amylose content (AC) was positively correlated with
hAM (r = 0.99,
p < 0.01) and also had a positive correlation with
XAP1, XAP2, XAM, hAP2/AP1, fb2, and fb3, but a negative correlation with fa and fb1, suggesting the longer B chains and longer amylose chains led to a higher AC. AC is synthesized by the action of GBSS encoded by the
Wx gene. All the rice materials contain the same
Wx allele (
Figure 1), so the difference in AC was attributed to the deficiency in BEIIb. The deficiency in BEIIb is responsible for the synthesis of larger amylopectin and amylose molecules with longer chains (
Table 1) by which the synthesis of short chains of amylopectin was suppressed and the elongation of long chains was promoted [
26,
37].
BEIIb has an important effect on the crystalline structures of starches by modifying the synthesis of A and B1 chains [
43]. The significant changes in the rice amylopectin CLDs in breeding lines with the
be2b allele modified the RC of the starch granules. RC had a negative correlation with
XAP1, XAP2, XAM, hAM, hAP2/hAP1, AC, fb3, and
, but a positive correlation with fa and fb1, which was in agreement with the result of Zhang et al. [
26]. The results indicated that lower AC and shorter amylose chain length, or more amylopectin A and B1 chains, would increase the RC.
The 1045/1022 cm
−1 ratio had a negative correlation with fa (r = 0.92,
p < 0.01), but had a positive correlation with
XAP1, XAP2, XAM, hAP2/AP1, fb3, and
(
Table S2), which indicated that longer amylopectin chains can form more double helices and increase the amount of short-range order [
26]. RC had a negative correlation with the 1045/1022 cm
−1 ratio, but a positive correlation with the 1022/995 cm
−1 ratio (
Table S2), which seemed to be contradictory. A possible explanation is that the FTIR data represent the ratio of the proportion of ordered structure to unordered structure, which is irrelevant to long-range order [
33].
GT is genetically controlled by SSIIa, whose function is to elongate the short A and B1 chains of amylopectin with DP < 10 to form long B1 chains [
15]. T
o, T
p, and T
c were found to be positively correlated with
XAP1, XAP2, XAM, hAP2/AP1, fb2, fb3,
, AC, and the 1045/1022 cm
−1 ratio, but had a negative correlation with RC and the 1022/995 cm
−1 ratio (
Table S2). This result was similar to that of Zhang et al. [
26], suggesting these correlations are derived from the
be2b allele in the rice materials. It should be noted that for the common rice starches, GT generally had a negative correlation with the number of fa chains but a positive correlation with fb1 content [
14]. In this study, the correlation between fa and fb1 was positive (r = 0.41,
p > 0.05), indicating these starches are different from common starches.
The PV, HPV, CPV, SB, and CS had a negative correlation with
XAP1, XAP2, XAM, hAP2/AP1, fb2, fb3,
and AC, but a positive correlation with fb1 (
Table S2), which is in agreement with previous reports [
26,
44]. A large amylose chain content, or a large number of long B chains in amylopectin, is expected to extend through crystallites connecting multiple clusters, increasing the integrity of the starch granules, leading to the inhibition of starch swelling [
43] and less resistance to shearing.
In conclusion, the structural and physicochemical properties of rice breeding lines changed with different BEIIb and SSIIa alleles. The BEIIb-deficient mutant starches had a higher AC with more amylose long chains, a larger amount of amylopectin long B chains, and a higher degree of molecular order, but a smaller amount of short chains and smaller RC, leading to a higher GT and lower ΔH and pasting viscosities. Therefore, BEIIb is more important in determining the structural and physicochemical properties than SSIIa. The combination of be2b/GC and be2b/TT showed no significant difference, suggesting that the function of SSIIa was not important in the be2b allele background. The resistant starch content was not measured in this study, but the fact that the ae mutant has high resistant-starch content is well known. In the near future, the breeding lines in the background be2b will be used for breeding new rice varieties with high resistant-starch content, which will benefit human health.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}