3.1. Scope and Search Protocol
Starch enzymatic degradation has been investigated for decades with continuing interest since 2016. Using a literature research engine (Web of Science, WoS) and the procedure described in
Figure 1, 57 articles published between 2016 and 2020 on this topic have been selected. The results of the selection procedure as well as the information extracted for each article are supplied as
Supplementary Materials.
In the first stage of the selection procedure, a list of 2053 journal articles covering the last 30 years was obtained from the WoS using a general research equation about starch hydrolysis by α-amylase, explicitly excluding acid hydrolysis (
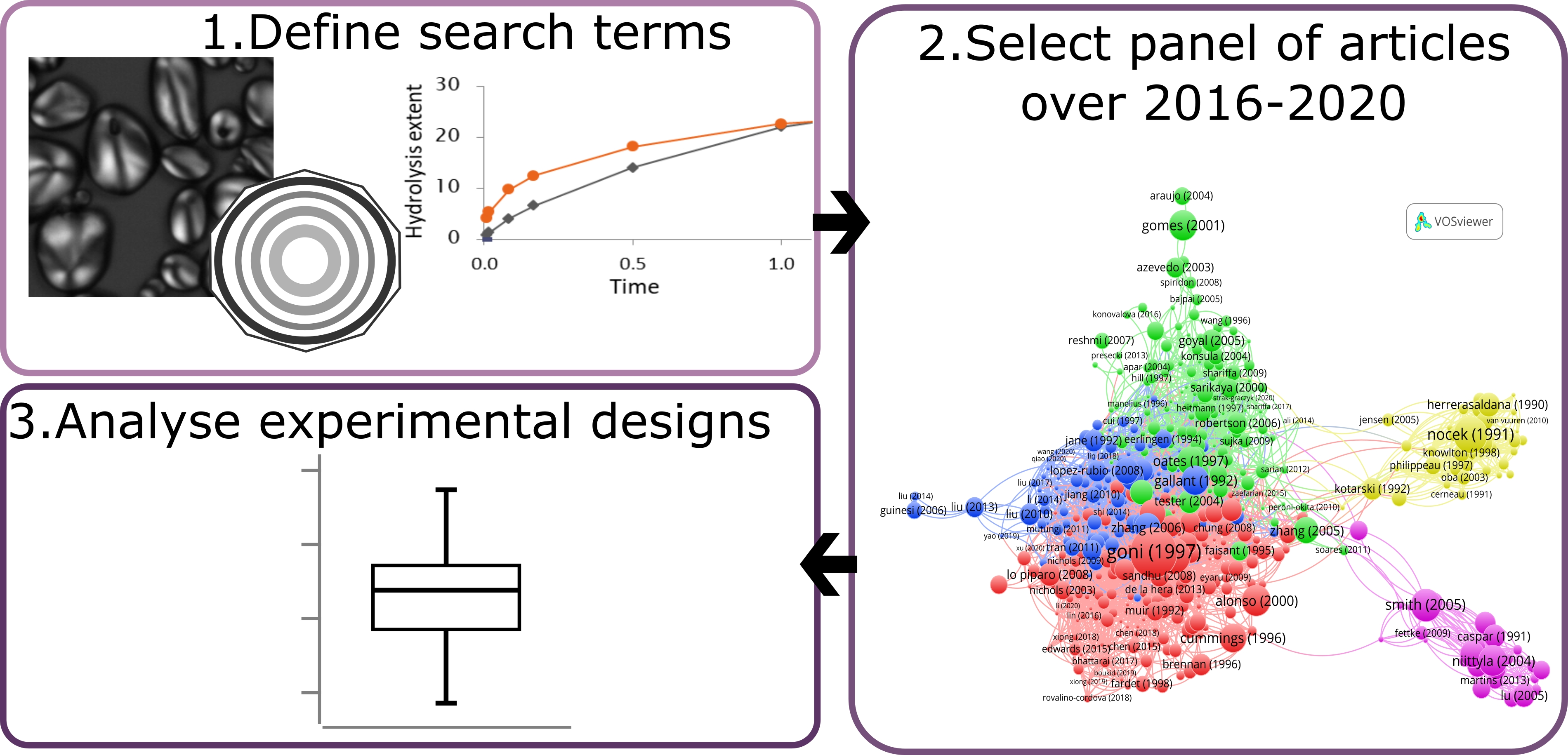
Figure 1). Stage 2 aimed at narrowing down this list to articles that were part of the major citation network on the topic. To this end, we performed a citation network analysis using a dedicated tool (VOSviewer, [
64]). Citation networks are made of nodes, which represent articles, edges or links, which represent inter-citations between the articles. From this information, VOSviewer computes a distance-based map where the distance between two articles reflects their similarity, i.e., the citations they have in common. From the list of 2053 articles, we produced a complete citation map of 6178 links and a distribution of the number of inter-citations per article (
Figure A1 and
Figure A2). The distribution follows a steep decreasing curve, and 509 articles have no links with the others, it is also visible that the network is a “dense” core group of articles. Given the distribution, we estimated that a set of 500 articles was large enough to capture a consistent group of interlinked articles on the topic, while having a fair representation of the ongoing research. Using VOSviewer again, we selected a set of 500 articles with the highest similarity level, starting from Goñi et al. [
15], which is the article with the highest number of links as well as a reference on the topic. The resulting citation map is provided in
Figure A3. It has 3195 links and only two articles without links, which indicates that we succeeded in sampling the “densest” part of the network. In the final stages, we retained the publications of the last five years and manually removed the articles not in the scope.
The information about the 57 articles of the panel used for the following analysis is supplied as an excel table; in the
Supplementary Materials, each article is assigned an id with the name of the first author, the year of publication and, when needed, a letter (‘a’, ‘b’…) to distinguish between articles with same name that were published in the same year.
Table 1 reports the topics addressed by the panel. The large majority (70%) focused on the factors and mechanisms influencing starch amylolysis. The remaining papers described the effect of modification treatments on starch structure and amylolysis to meet various end-product performances. The modifications included an amylolysis optimization process involving novel degrading enzymes as well as comparing and developing different kinetic models to improve amylolysis analysis.
3.2. Type and Size of Samples
Cereal starch from maize, rice and wheat, together with potato starch, were extensively studied in our publication panel (
Table 2), most likely as ingredients available worldwide. Maize starch was the most frequently investigated among crops (35% of the papers); it includes the starch from well-known commercial lines of maize mutants, e.g.,
wx (waxy),
ae (amylose-extender) and
su2 (sugary), with various granular and macromolecular structures [
65]. Other starch types, including cassava sweet potato, bean and pea starch, complete the publication list.
Table 2 also provides the number of papers that compare wild-type, mutant, transgenic lines and single-segment substitution lines starch, all with a similar genetic background. This type of experiments is reported in 14% of the panel and involves mainly rice (
Table 2). Surprisingly while maize mutant starch is present in many publications, the panel does not include specific works on maize genetic variants. This suggests that this crop has been already well covered in past publications. Starch obtained from genetic variants presents structural alterations caused by either deletion of or modifications of the expression (silencing, downregulation, upregulation) in the enzymes of the starch biosynthesis pathway. Those enzymes are generally acting on the elongation, the branching or the debranching of the starch polymers [
66]. Genetic variants introduce targeted starch structural modifications and often generate near isogenic control lines, allowing comparative studies focusing primarily on the altered features [
37,
63].
The experimental designs found within the panel include on average eight distinct samples (
Figure 2), which allows the investigation of a limited number of factors. In practice, the number of samples is limited by the availability of starch materials with desirable characteristics and by the experimental resources required to characterize the samples or to run the hydrolysis assays. Hence, many publications included a selective number of diverse samples with marked characteristics and/or innovative structural or functional characterization techniques. An alternative approach was to analyse a higher number of samples using established analytical techniques and hydrolysis assays, as for Martens et al. [
28,
67]. These authors reported the results of a retrospective power analysis to confirm that the number of samples were sufficiently large to reveal the discrepancies between the degradation kinetics.
3.3. Characterisation of Starch Structure
The number of studied factors described in the publication panel varies from one to nine, with an average of four (
Figure 3). No trend could be identified in the number of factors investigated over the last five years.
The most frequent factors describe distinct organizational levels of native starch (from granular to molecular level) as well as few functional properties (
Table 3). At the molecular level, amylopectin chain length distribution and amylose content were both measured in 32% of the panel. However, as mentioned in
Section 2.1, the impact of amylose content on amylolysis is still unclear, and a few works of the panel observed an impact, e.g., Kuang, Xu, Wang, Zhou & Liu [
68] and Lin et al. [
12], while others did not, e.g., Martens et al. [
28]. The amylose content is commonly estimated with the iodine binding colorimetric method [
69], even though this method can overestimate the amount of amylose due to the formation of amylopectin-iodine complexes [
70]. Therefore, alternatives, e.g., concanavalin A precipitation and the size-exclusion chromatography weight distributions, are increasingly applied for a more accurate estimation [
12,
28,
71,
72,
73]. Investigations of minor non-starch components are reported in 17% of the panel. This concerns especially the proteins either via the protein content estimation using nitrogen estimation by the Kjeldahl method, the Dumas method, a nitrogen analyzer [
71,
74,
75] or by comparing the hydrolysis results before and after protein removal (e.g., [
51]). At the crystalline and lamellar levels, the crystallinity (type and quantification) measured by X-ray diffraction appears to be the most studied feature (53% of the panel), certainly due to the fact that the starch crystallinity is often effectively correlated to physical, mechanical and technological properties and of course to amylolysis [
43]. The lamellar structure, described through lamellar ordering and thickness using small-angle X-ray scattering, is also reported in 23% of the panel. At the granular level, 37% of the panel includes a granule morphology analysis by Scanning Electron Microscopy (SEM). SEM is used to determine the size and shape of the granules, as well as their surface properties, number of pores, size and dispersity and degradation signs [
28,
76,
77]. The granule size distribution, usually determined by Laser Diffraction Sizing (LDS), is present in only 19% of the panel.
The functional properties reported in the panel publications are mainly related to starch gelatinization (28%) measured by Differential Scanning Calorimetry (DSC). DSC reflects on the swelling, the leaching of soluble polysaccharides, the disruption of the molecular order and the melting of crystallites [
78], thus providing a compelling understanding of starch gelatinization properties. In comparison, results from the Rapid Visco Analyzer (RVA) (pasting properties in
Table 3) are much less common (<9%), possibly because RVA requires a significant amount of biological material for a single analysis and it has a rather low throughput.
Each of the uncommon factors (“others” in the
Table 3) had been measured in less than five studies in the panel. They can describe the starch intrinsic structure, e.g., molecular weight distribution and endogenous lipid content, or extrinsic factors, e.g., enzyme and substrate concentrations, temperature, incubation time, plant genetics and environment.
3.4. Characterisation of Hydrolysis Kinetics
In most publications, the extent of starch enzymatic hydrolysis is determined by an enzymatic assay quantifying the glucose release. The most frequent method, with 51% of the panel, is the glucose oxidase and peroxidase (GOPOD) assay [
79] (
Table 4). It has high specificity but lacks relative stability over time and has a relatively high cost per sample [
80]. The measurement of reducing ends by dinitrosalicylic acid (DNS) [
81] and by 4-hydroxybenzoic acid hydrazide (PAHBAH) colorimetric methods are alternative options to glucose assays that are simple to apply with relatively low cost. This family of dosage techniques is in general much more rapid and easier to handle than the dosage of total solubilized sugars by the orcinol–H
2SO
4 method used in older works (e.g., in [
37]). Nevertheless, with equimolar amounts of maltodextrins, the response of the reducing sugar by the DNS method increased with the increasing number of glucose units [
82], which leads to an overestimation of the hydrolysis extent.
For 91% of the panel, an enzymatic assay is used to obtain kinetic data of the starch degradation by an α-amylase (30%), sometimes in association with an amyloglucosidase (61%). Several methods were used to interpret the kinetic data. Twelve percent of the panel used a starch classification method, more precisely derived from Englyst’s method, to identify starch fractions with decreasing digestibility based on their digestion time [
14].
Figure 4a presents an example of the classification of digested starch from three distinct sources into different fractions of increasing digestibility level. This approach basically requires one measurement of the hydrolysis extent per fraction. Identification of Resistant Starch (RS) content via the AACC (American Association of Cereal Chemists)-approved Method, which separates the RS from the non-resistant starch and then hydrolyzes this fraction into glucose using AMG, was used by Uriarte-Aceves et al. [
71], whereas the quantification via the AOAC (Association of Official Agricultural Chemists) Method [
83] was not reported in the panel. Eighty-eight percent of the panel described the starch degradation over time using a kinetic model fitted to the data, which appears to be a common practice to quantitatively analyze the kinetics parameters [
18]. A simple kinetic model can describe not only the degradability level at different times but also the shape of the curve describing the hydrolysis time course and the reaction rate coefficient.
Figure 4b presents examples of time-course measurements of the hydrolysis extents fitted with a modified first-order kinetic model, i.e., the Weibull model, Equation (6).
Seventy percent of the panel applied the empirical first-order kinetics (
Table 5) using Equation (1) proposed by Goñi, Garcia-Alonso & Saura-Calixto [
15]:
where
is the product concentration at time
t,
is the production concentration at the end of the hydrolysis and
k is the reaction rate constant. Although the first-order kinetics describes well the hydrolysis profile, it needs an accurate estimation of
, which can be difficult to achieve even with a prolonged hydrolysis assay [
84]. The application of the log of slope (LOS) plot via equation:
by Butterworth et al. [
84] allows the calculation of
from the intercept on the y axis that could avoid the aforementioned issue. Forty-six percent of the panel applied the LOS plot for obtaining a more reliable estimation of
and
k.
Kinetics data do not always follow a first-order kinetics, which led to adaptations of Equations (1) and (2). Two main adaptations of the first-order kinetics model have been proposed to obtain a better fitting of kinetic data, a sequential model that describes a fraction of the starch digested before others and a parallel model that describes two or more fractions of starch digested simultaneously at different rates [
85]. Edwards, Maillot, Parker and Warren [
86] applied a sequential first-order kinetic model (Equation (3)), whereas Li et al. [
85] and Bello-perez, Agama-Acevedo, Garcia-Valle and Alvarez-Ramirez [
87] applied variants of parallel first-order kinetic models, respectively Equations (4) and (5):
where
is the time of intersection of two consecutive reactions;
is the concentration of product at
;
and
are the corresponding rate constants for each type of starch;
is the starting concentration of digestible starch;
and
are concentrations of two digestible starch types with distinct susceptibilities to amylolysis.
Yt is the normalized starch concentration and
,
and
are the concentrations of
RS,
RDS and
SDS;
and
are the rate constants of
RDS and
SDS. Compared to the sequential model, the parallel model avoids breakpoints at the times of intersection between two hydrolysis phases [
85].
A Weibull model proposed by Kansou, Buléon, Gérard & Rolland-Sabaté [
88] is another variant of the first-order kinetics model (Equation (6)). In the panel, it has been applied by Olawoye & Gbadamosi [
89] to obtain an accurate fitting of biphasic hydrolyses. In this model the reaction rate-related parameter
k is affected by a parameter
h that reflects the time-dependency of the reaction rate coefficient:
3.5. Statistical Analysis of Starch Structure-Amylolysis Relationships
The majority of the panel publications employed standard statistical analysis methods (
Table 6). Up to 72% present the results generated using ANOVA, often followed by the result of a test distinguishing the different groups, e.g., the Tukey test, 19% computed linear correlations (Pearson) or the most robust rank correlation (Spearman) that assesses monotonic relations between two variables. Multivariate statistical analyses are reported in only 7% of the panel (
Table 6). Principal component analysis (PCA) is the most common multivariate analysis, it is used to identify the multilinear relations between several factors via a new set of orthogonal variables, so-called principal components. PCA allows the relation of combinations of interacting factors such as granule diameter, pores, crystalline type and side-chain length of amylopectin with kinetic parameters of the amylolysis (e.g., [
28,
39]).
The conditions of application of the usual linear models, including the linear regression models or the ANOVA, are often problematic in the context of starch structure-degradation studies because of the collinearity of the data [
39]. Despite this limitation, the relatively small size of the datasets envisioned in
Section 3.2 and
Section 3.3 hinders the application of multifactorial analysis or clustering techniques alike.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}