
Metabolomic Analysis, Combined with Enzymatic and Transcriptome Assays, to Reveal the Browning Resistance Mechanism of Fresh-Cut Eggplant

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
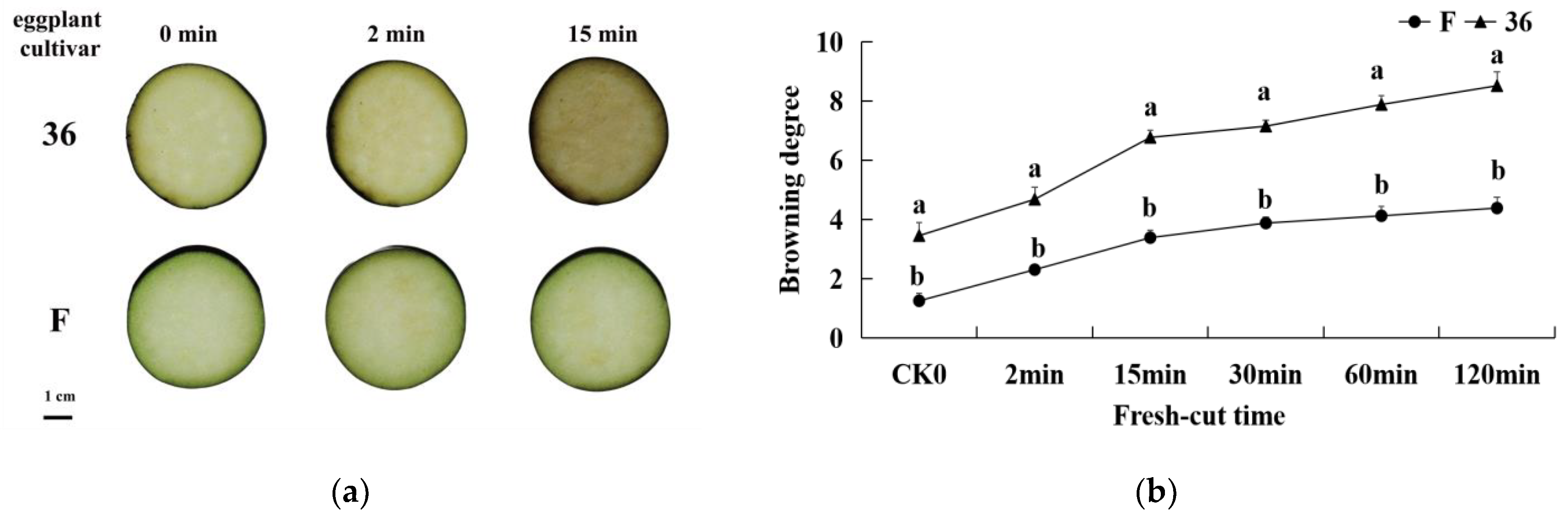
2.2. Browning Degree
2.3. Metabolite Identification and Statistical Analysis
2.4. Integrated Analysis of the Metabolome and Transcriptome
2.5. Membrane Lipid Peroxidation (MDA, H2O2, and O2−)
2.6. Activities of PPO, LOX, PLD, PAL, CAT, SOD, POD, and APX
2.7. Determination of Antioxidant Capacity (DPPH, ABTS, and FRAP)
2.8. Determination of Total Phenol (TP) Content
2.9. Statistical Analyses
3. Results
3.1. Phenotypic Characteristics of Physiological Responses of Fresh-Cut Eggplant
3.2. Qualitative and Quantitative Analysis of Metabolites
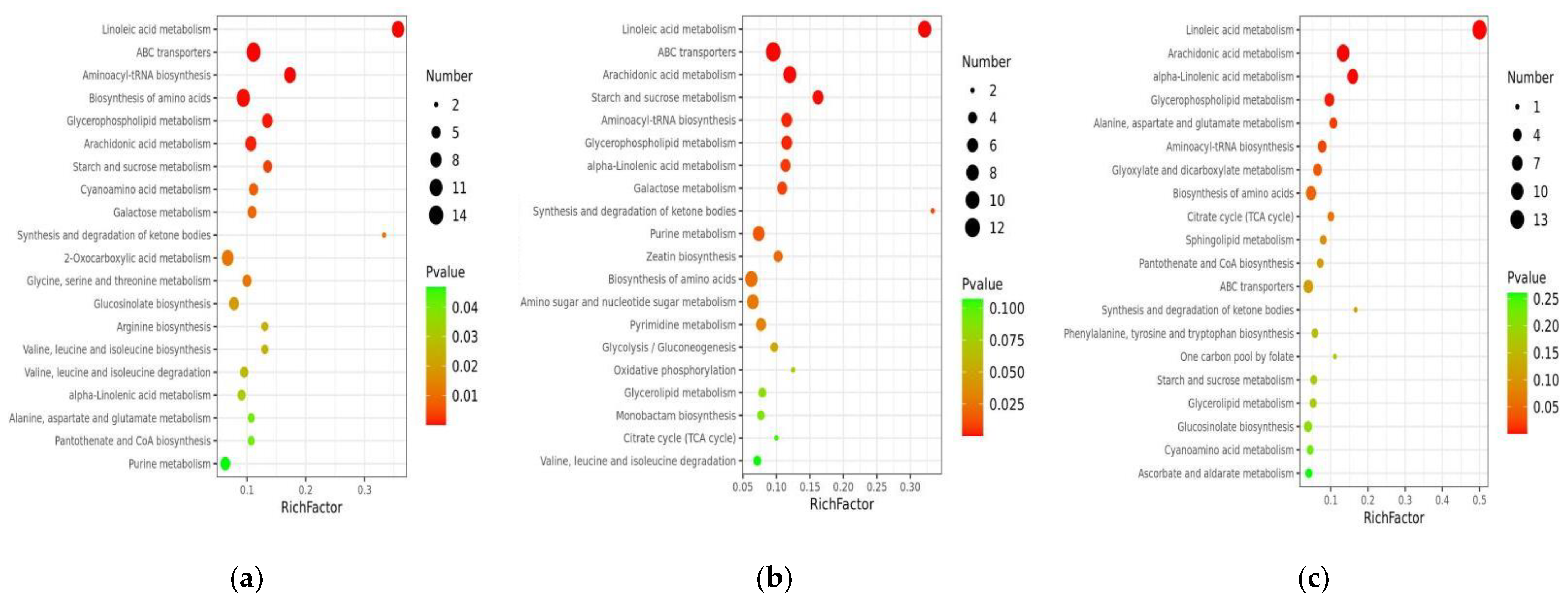
3.3. Differential Metabolites (DEMs) Screening, Functional Annotation, and Enrichment Analysis
3.4. Enrichment Analysis of DEMs
3.5. Integrated Analysis of the Transcriptome and Metabolome of the Fresh-Cut Eggplant
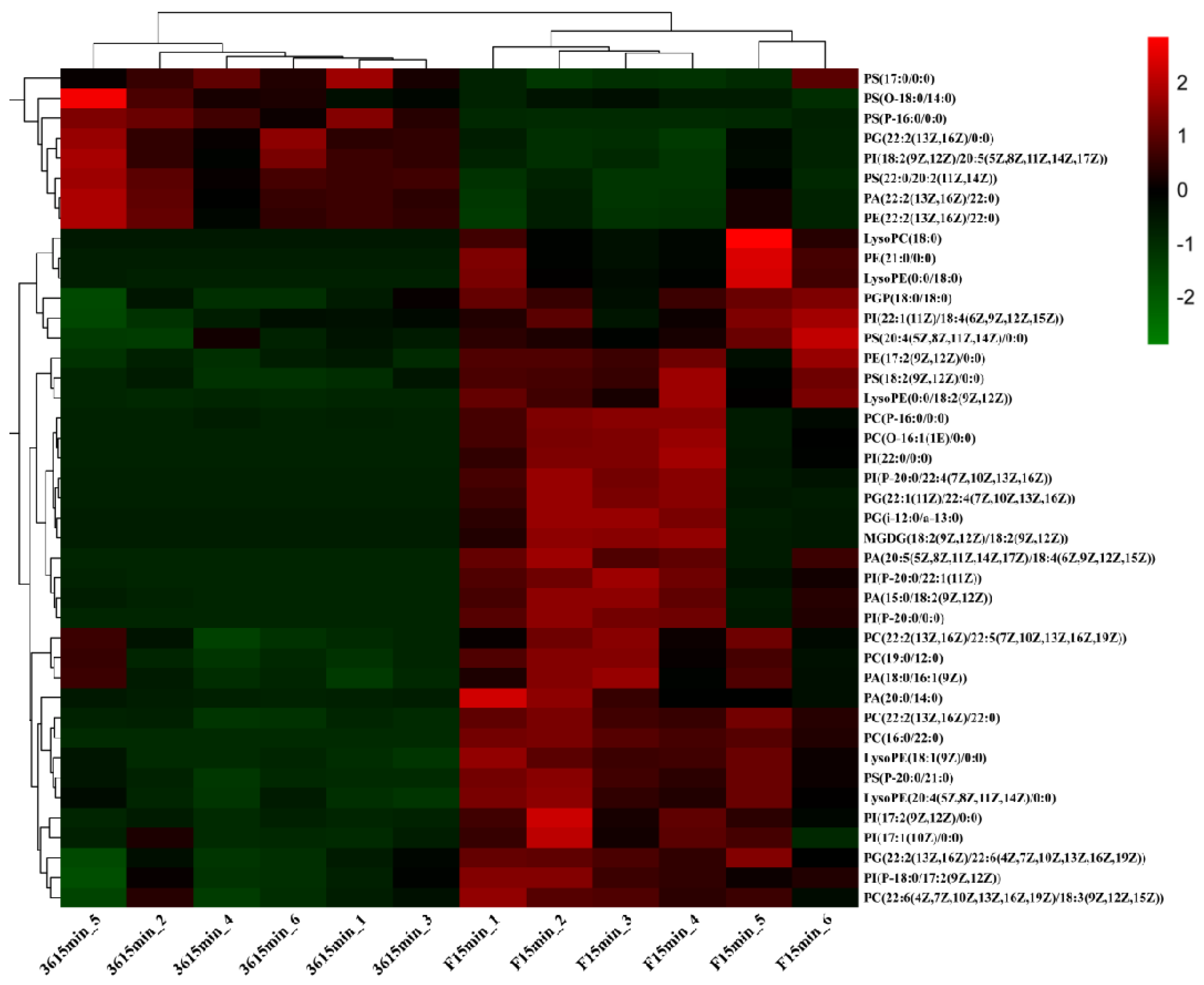
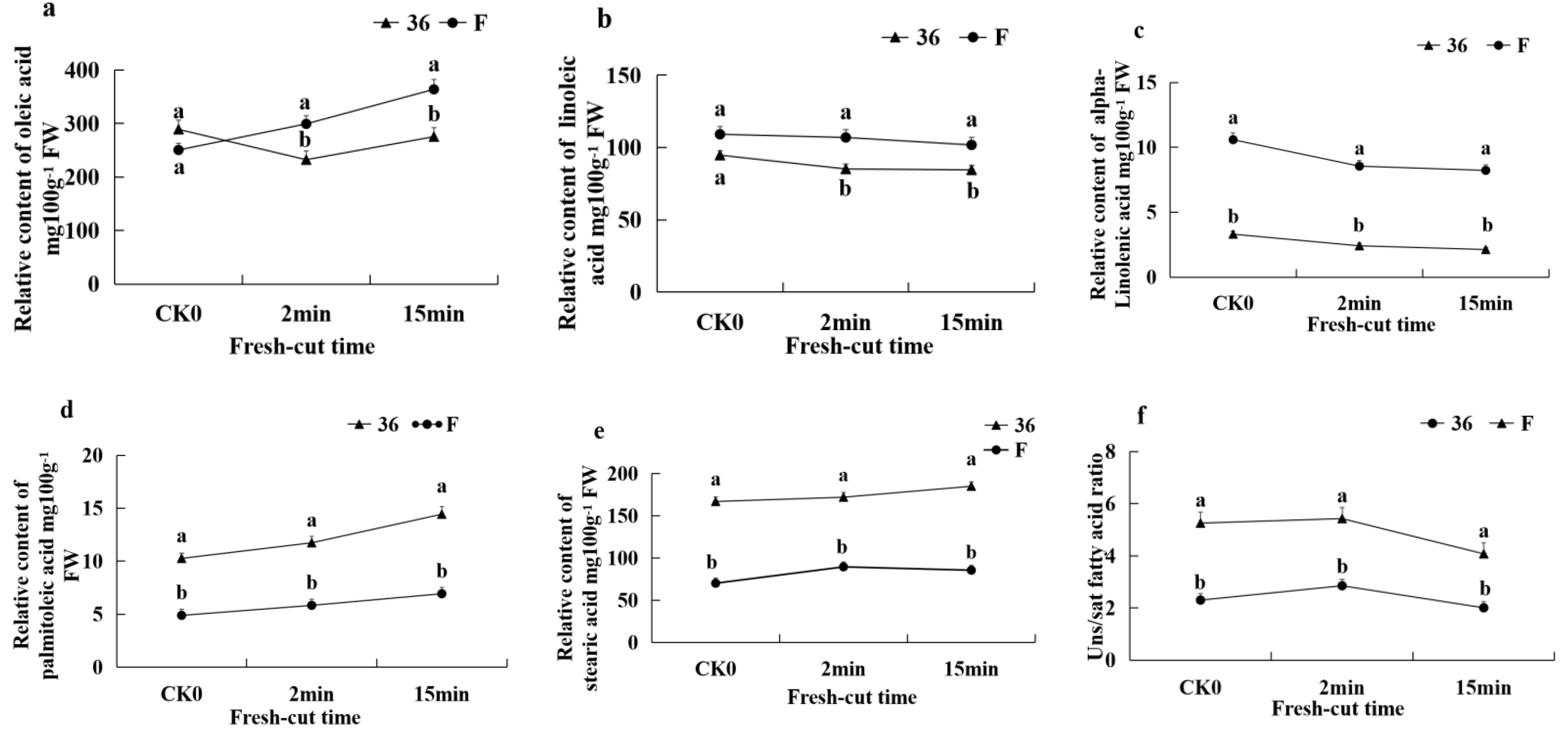
3.5.1. Changes in Membrane Lipid Composition of Fresh-Cut Eggplant
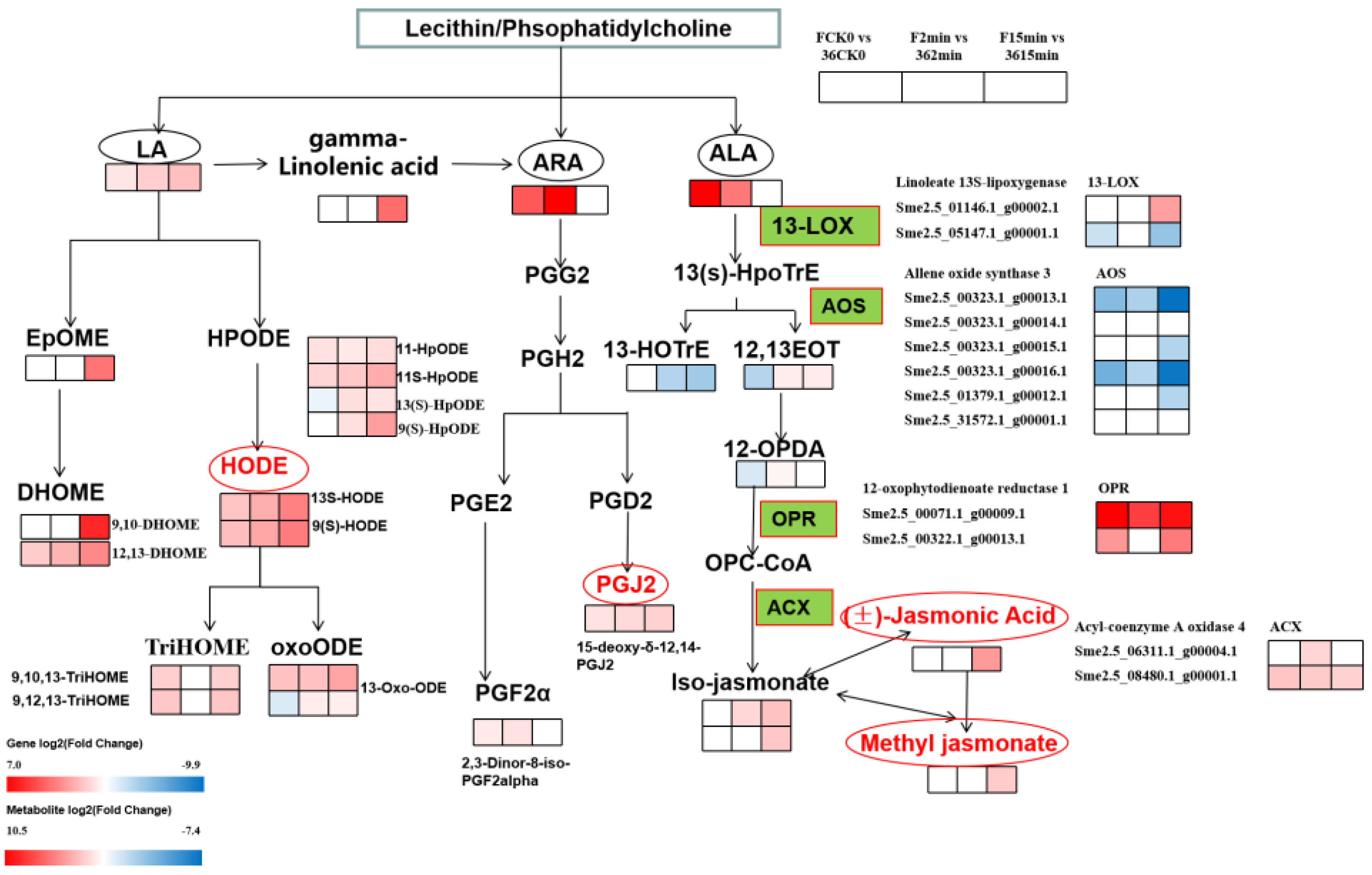
3.5.2. Expression of DEGs and DMEs Related to Lipid Metabolism
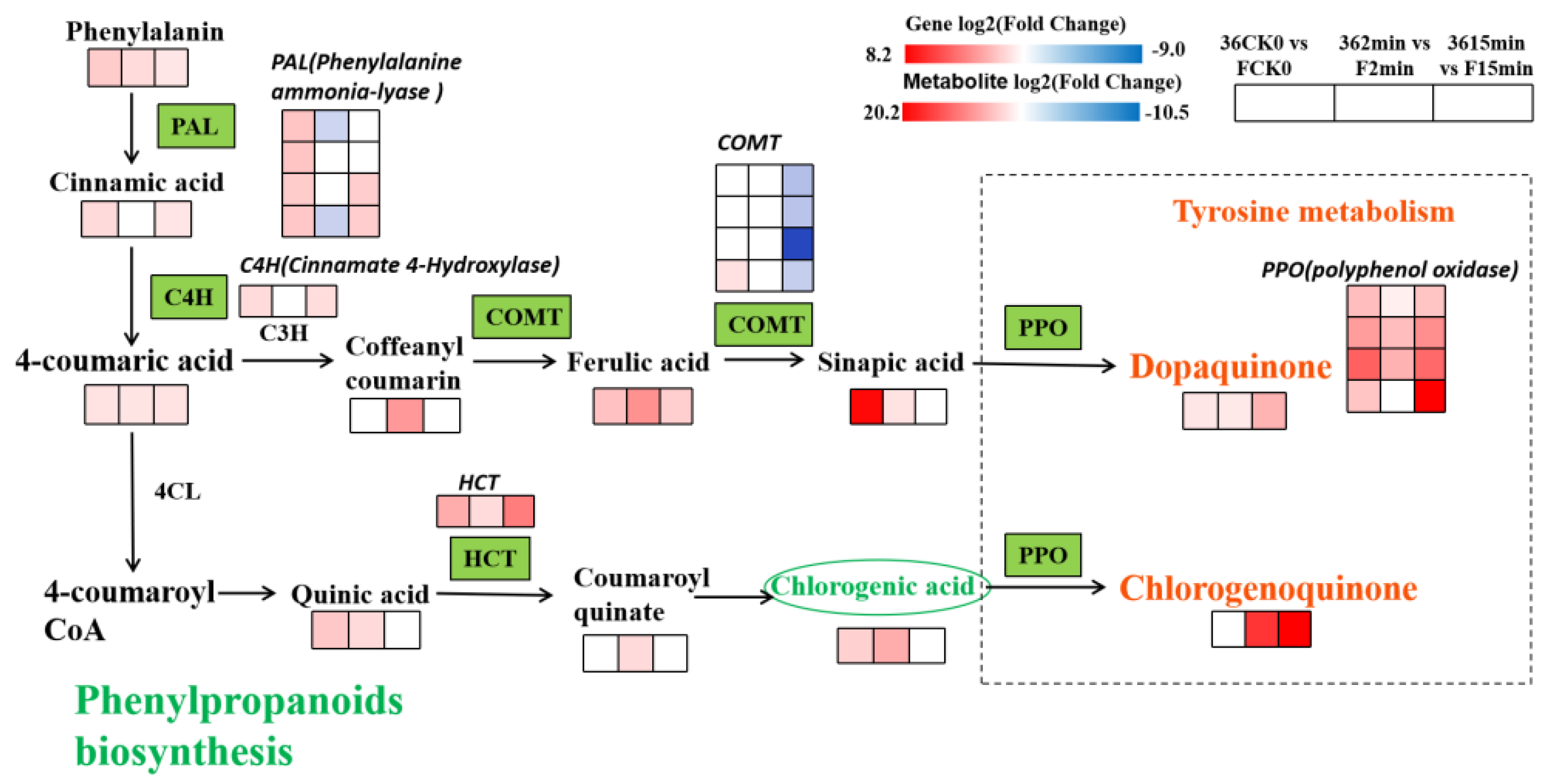
3.5.3. Expression of DEGs and DMEs Related to Phenylpropane Biosynthesis
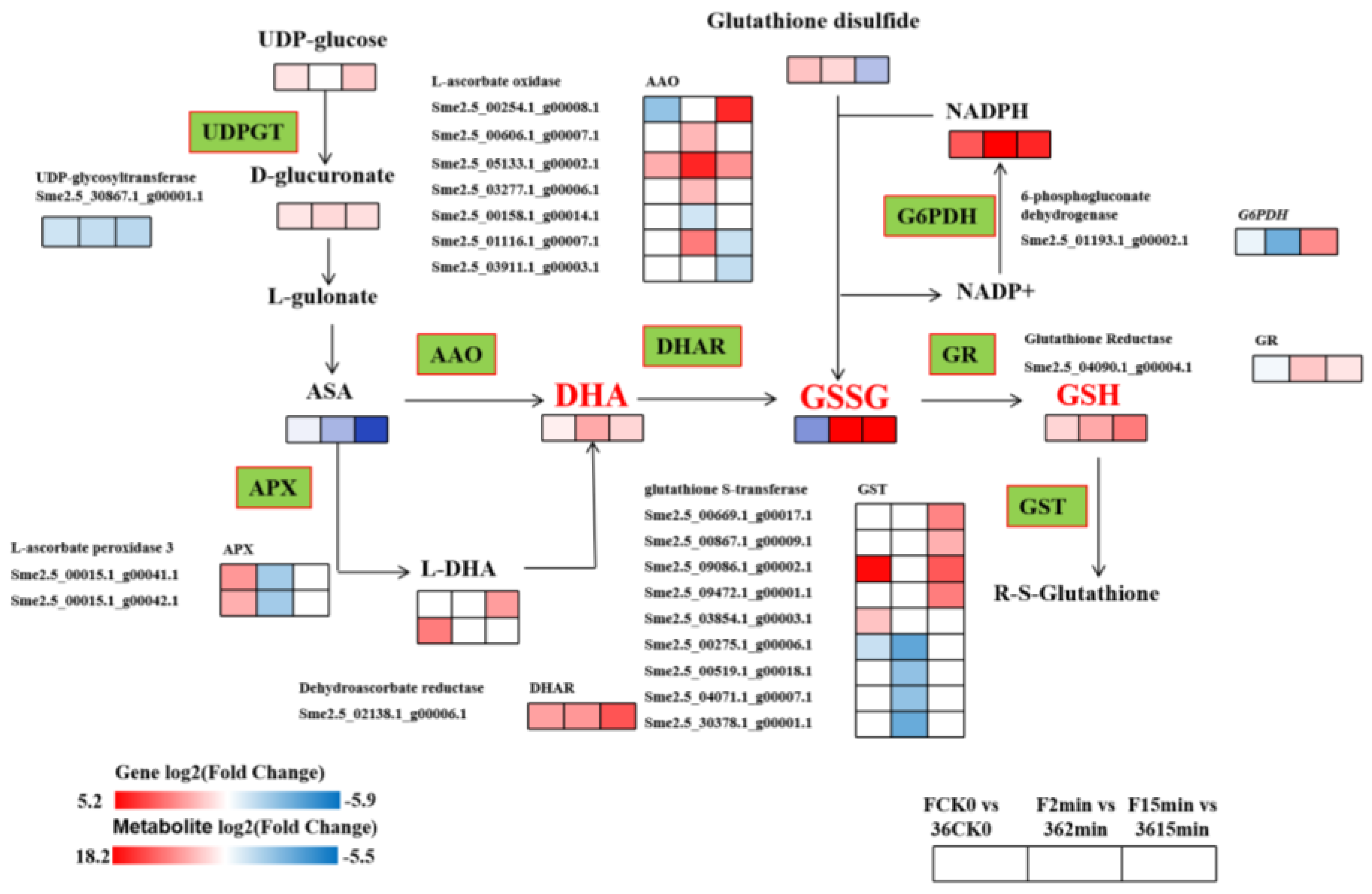
3.5.4. Expression of DEGs and DMEs Related to Glutathione Metabolism
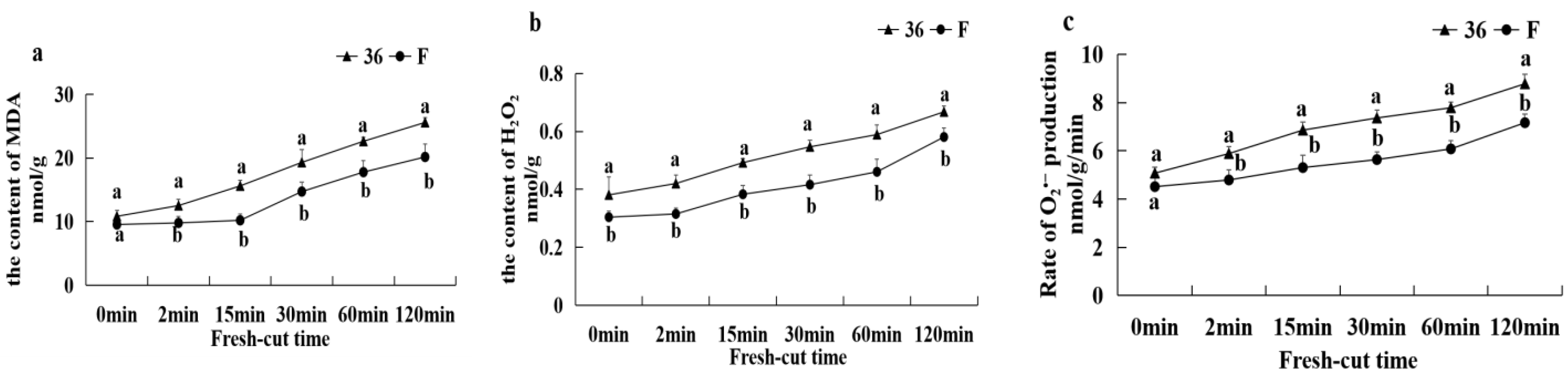
3.5.5. Malondialdehyde Content, H2O2 Content, and O2− Production Rate
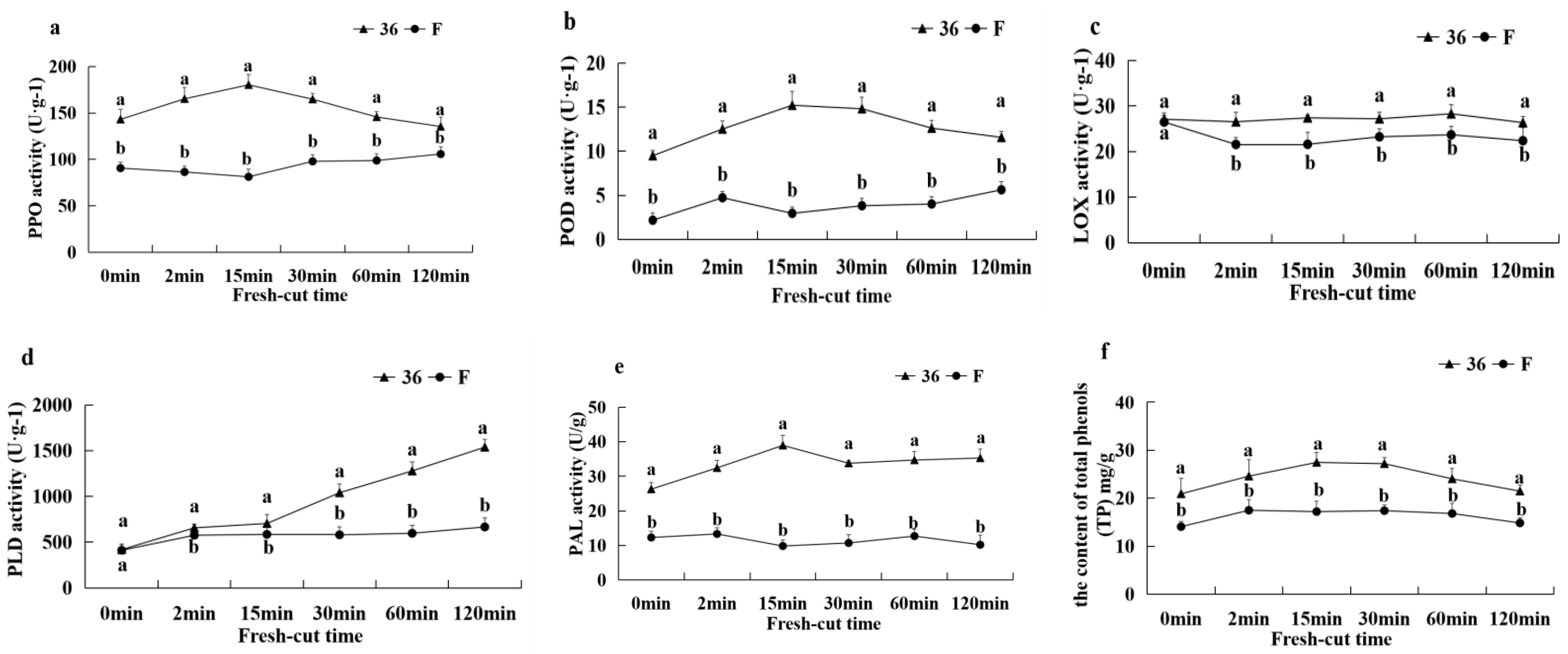
3.5.6. Activities of PPO, LOX, PLD, POD, PAL, and TP Content
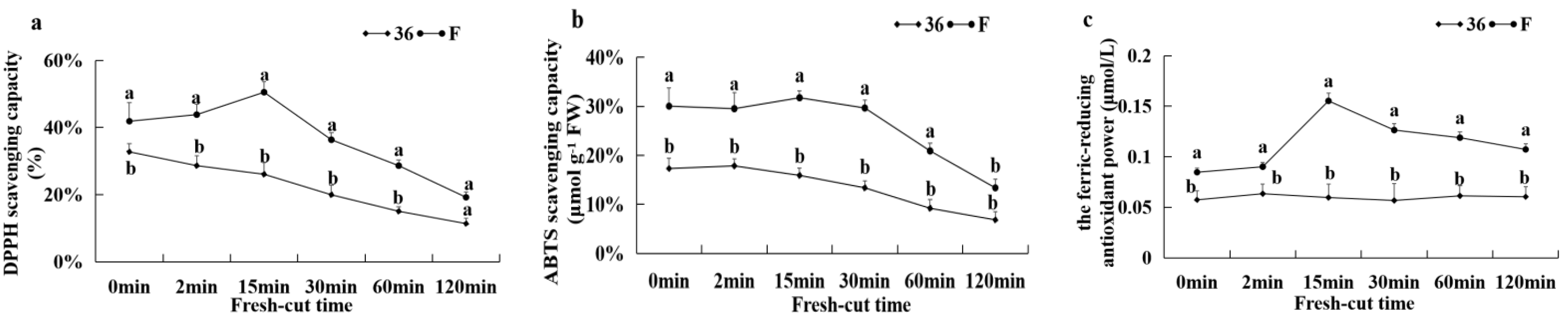
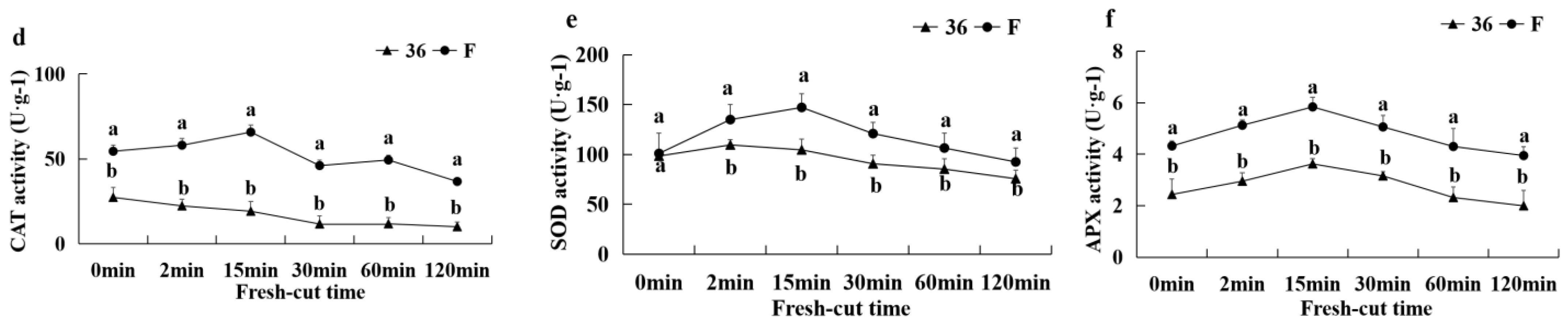
3.5.7. Antioxidant Capacity and the Activities of CAT, APX, and SOD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, A.; Sharma, V.; Jain, B.T.; Kaushik, P. Heterosis breeding in eggplant (Solanum melongena L.): Gains and provocations. Plants 2020, 9, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, B.B.; Gautam, S.; Sharma, A. Purification and characterisation of polyphenol oxidase (PPO) from eggplant (Solanum melongena). Food Chem. 2012, 134, 1855–1861. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Long, Q.; Gao, F.; Han, C.; Jin, P.; Zheng, Y. Effect of cutting styles on quality and antioxidant activity in fresh-cut pitaya fruit. Postharvest Biol. Technol. 2017, 124, 1–7. [Google Scholar] [CrossRef]
- Bustos, M.C.; Mazzobre, M.F.; Buera, M.P. Stabilization of refrigerated avocado pulp: Effect of Allium and Brassica extracts on enzymatic browning. LWT Food Sci. Technol. 2015, 61, 89–97. [Google Scholar] [CrossRef]
- Ali, S.; Khan, A.S.; Malik, A.U.; Shaheen, T.; Shahid, M. Pre-storage methionine treatment inhibits postharvest enzymatic browning of cold stored ‘Gola’ litchi fruit. Postharvest Biol. Technol. 2018, 140, 100–106. [Google Scholar] [CrossRef]
- Zhang, S.; Lin, H.; Lin, M.; Lin, Y.; Chen, Y.; Wang, H.; Lin, Y.; Shi, J. Lasiodiplodia theobromae (Pat.) Griff. & Maubl. reduced energy status and ATPase activity and its relation to disease development and pericarp browning of harvested longan fruit. Food Chem. 2019, 275, 239–245. [Google Scholar] [CrossRef]
- Kong, X.; Wei, B.; Gao, Z.; Zhou, Y.; Shi, F.; Zhou, X.; Zhou, Q.; Ji, S. Changes in membrane lipid composition and function accompanying chilling injury in bell peppers. Plant Cell Physiol. 2018, 59, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Cao, Y.; Zhou, L.; Jiang, C.Z.; Feng, Y.; Wei, S. Effects of postharvest curing treatment on flesh colour and phenolic metabolism in fresh-cut potato products. Food Chem. 2015, 169, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Holthuis, J.C.; Menon, A.K. Lipid landscapes and pipelines in membrane homeostasis. Nature 2014, 510, 48–57. [Google Scholar] [CrossRef]
- Sun, H.; Zhou, X.; Zhou, Q.; Zhao, Y.; Kong, X.; Luo, M.; Ji, S. Disorder of membrane metabolism induced membrane instability plays important role in pericarp browning of refrigerated ‘Nanguo’ pears. Food Chem. 2020, 320, 126684. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lv, M.; Li, G.; Jiang, Y.; Fu, W.; Zhang, L.; Ji, S. Effect of intermittent warming on alleviation of peel browning of ‘Nanguo’ pears by regulation energy and lipid metabolisms after cold storage. Postharvest Biol. Technol. 2018, 142, 99–106. [Google Scholar] [CrossRef]
- Kaushik, P.; Gramazio, P.; Vilanova, S.; Raigón, M.D.; Prohens, J.; Plazas, M. Phenolics content, fruit flesh colour and browning in cultivated eggplant, wild relatives and interspecific hybrids and implications for fruit quality breeding. Food Res. Int. 2017, 102, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Docimo, T.; Francese, G.; De Palma, M.; Mennella, D.; Toppino, L.; Lo Scalzo, R.; Mennella, G.; Tucci, M. Insights in the fruit flesh browning mechanisms in Solanum melongena genetic lines with opposite postcut behavior. J. Agric. Food Chem. 2016, 64, 4675–4685. [Google Scholar] [CrossRef]
- Kan, J.; Xie, W.; Wan, B.; Huo, T.B.; Lin, X.P.; Liu, J.; Jin, C.H. Heat-induced tolerance to browning of fresh-cut lily bulbs (Lilium lancifolium Thunb.) under cold storage. J. Food Biochem. 2019, 43, e12816. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, A.; Shang, J.; Zhu, Z.; Li, Y.; Wu, X.; Zha, D. Study on browning mechanism of fresh-cut eggplant (Solanum melongena L.) based on metabolomics, enzymatic assays and gene expression. Sci. Rep. 2021, 11, 6937. [Google Scholar] [CrossRef]
- Proestos, C.; Lytoudi, K.; Mavromelanidou, O.K.; Zoumpoulakis, P.; Sinanoglou, V.J. Antioxidant capacity of selected plant extracts and their essential oils. Antioxidants 2013, 2, 11–22. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of ‘antioxidant power’: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Busatto, N.; Farneti, B.; Commisso, M.; Bianconi, M.; Iadarola, B.; Zago, E.; Ruperti, B.; Spinelli, F.; Zanella, A.; Velasco, R.; et al. Apple fruit superficial scald resistance mediated by ethylene inhibition is associated with diverse metabolic processes. Plant J. 2018, 93, 270–285. [Google Scholar] [CrossRef] [Green Version]
- Busatto, N.; Farneti, B.; Tadiello, A.; Vrhovsek, U.; Cappellin, L.; Biasioli, F.; Velasco, R.; Costa, G.; Costa, F. Target metabolite and gene transcription profiling during the development of superficial scald in apple (Malus x domestica Borkh). BMC Plant Biol. 2014, 14, 193. [Google Scholar] [CrossRef] [PubMed]
- Tao, N.; Wang, R.; Xu, X.; Dong, T.; Zhang, S.; Liang, M.; Wang, Q. Xanthosine is a novel anti-browning compound in potato identified by widely targeted metabolomic analysis and in vitro test. Postharvest Biol. Technol. 2021, 171, 111367. [Google Scholar] [CrossRef]
- Min, T.; Bao, Y.; Zhou, B.; Yi, Y.; Wang, L.; Hou, W.; Ai, Y.; Wang, H. Transcription profiles reveal the regulatory synthesis of phenols during the development of lotus rhizome (Nelumbo nucifera Gaertn). Int. J. Mol. Sci. 2019, 20, 2735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Tomás-Barberán, F.A.; Saltveit, M.E. Wound-induced phenolic accumulation and browning in lettuce (Lactuca sativa L.) leaf tissue is reduced by exposure to n-alcohols. Postharvest Biol. Technol. 2005, 37, 47–55. [Google Scholar] [CrossRef]
- Han, C.; Li, J.; Jin, P.; Li, X.; Wang, L.; Zheng, Y. The effect of temperature on phenolic content in wounded carrots. Food Chem. 2017, 215, 116–123. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Jiang, Y.; Sun, J.; Liu, H.; Song, L.; Duan, X. Effects of short-term anoxia treatment on browning of fresh-cut Chinese water chestnut in relation to antioxidant activity. Food Chem. 2012, 132, 1191–1196. [Google Scholar] [CrossRef]
- Karamać, M.; Gai, F.; Longato, E.; Meineri, G.; Janiak, M.A.; Amarowicz, R.; Peiretti, P.G. Antioxidant activity and phenolic composition of amaranth (Amaranthus caudatus) during plant growth. Antioxidants 2019, 8, 173. [Google Scholar] [CrossRef] [Green Version]
- Supapvanich, S.; Pimsaga, J.; Srisujan, P. Physicochemical changes in fresh-cut wax apple (Syzygium samarangenese [Blume] Merrill & LM Perry) during storage. Food Chem. 2011, 127, 912–917. [Google Scholar] [CrossRef]
- Pinto, E.; Lentheric, I.; Vendrell, M.; Larrigaudiere, C. Role of fermentative and antioxidant metabolisms in the induction of core browning in controlled-atmosphere stored pears. J. Sci. Food Agric. 2001, 81, 364–370. [Google Scholar] [CrossRef]
- Chew, O.; Whelan, J.; Millar, A.H. Molecular definition of the ascorbate-glutathione cycle in Arabidopsis mitochondria reveals dual targeting of antioxidant defenses in plants. J. Biol. Chem. 2003, 278, 46869–46877. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, O.; Torres, C.A.; Moya-León, M.A.; Opazo, M.C.; Razmilic, I. Roles of the ascorbate–glutathione cycle, pigments and phenolics in postharvest ‘sunscald’ development on ‘Granny Smith’ apples (Malus domestica Borkh.). Postharvest Biol. Technol. 2014, 87, 79–87. [Google Scholar] [CrossRef]
- Scotti-Campos, P.; Pais, I.P.; Partelli, F.L.; Batista-Santos, P.; Ramalho, J.C. Phospholipids profile in chloroplasts of Coffea spp. genotypes differing in cold acclimation ability. J. Plant Physiol. 2014, 171, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lin, H.; Lin, Y.; Zhang, S.; Chen, Y.; Jiang, X. The roles of metabolism of membrane lipids and phenolics in hydrogen peroxide-induced pericarp browning of harvested longan fruit. Postharvest Biol. Technol. 2016, 111, 53–61. [Google Scholar] [CrossRef]
- Cao, S.; Yang, Z.; Cai, Y.; Zheng, Y. Fatty acid composition and antioxidant system in relation to susceptibility of loquat fruit to chilling injury. Food Chem. 2011, 127, 1777–1783. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, Y.; Lin, H.; Ritenour, M.A.; Shi, J.; Zhang, S.; Chen, Y.; Wang, H. Hydrogen peroxide-induced pericarp browning of harvested longan fruit in association with energy metabolism. Food Chem. 2017, 225, 31–36. [Google Scholar] [CrossRef]
- Mavangira, V.; Sordillo, L.M. Role of lipid mediators in the regulation of oxidative stress and inflammatory responses in dairy cattle. Res. Vet. Sci. 2018, 116, 4–14. [Google Scholar] [CrossRef]
- Muñoz, P.; Munné-Bosch, S. Oxylipins in plastidial retrograde signaling. Redox Biol. 2020, 37, 101717. [Google Scholar] [CrossRef]
- Raphael, W.; Sordillo, L.M. Dietary polyunsaturated fatty acids and inflammation: The role of phospholipid biosynthesis. Int. J. Mol. Sci. 2013, 14, 21167–21188. [Google Scholar] [CrossRef] [Green Version]
- Widemann, E.; Smirnova, E.; Aubert, Y.; Miesch, L.; Heitz, T. Dynamics of jasmonate metabolism upon flowering and across leaf stress responses in Arabidopsis thaliana. Plants 2016, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Sayyari, M.; Babalar, M.; Kalantari, S.; Martínez-Romero, D.; Guillén, F.; Serrano, M.; Valero, D. Vapour treatments with methyl salicylate or methyl jasmonate alleviated chilling injury and enhanced antioxidant potential during postharvest storage of pomegranates. Food Chem. 2011, 124, 964–970. [Google Scholar] [CrossRef]
- Ali, M.B.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate and salicylic acid induced oxidative stress and accumulation of phenolics in Panax ginseng bioreactor root suspension cultures. Molecules 2007, 12, 607–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangprayoon, P.; Supapvanich, S.; Youryon, P.; Wongs-Aree, C.; Boonyaritthongchai, P. Efficiency of salicylic acid or methyl jasmonate immersions on internal browning alleviation and physicochemical quality of Queen pineapple cv. ‘Sawi’ fruit during cold storage. J. Food Biochem. 2019, 43, e13059. [Google Scholar] [CrossRef] [PubMed]
- Taki, N.; Sasaki-Sekimoto, Y.; Obayashi, T.; Kikuta, A.; Kobayashi, K.; Ainai, T.; Yagi, K.; Sakurai, N.; Suzuki, H.; Masuda, T.; et al. 12-oxo-phytodienoic acid triggers expression of a distinct set of genes and plays a role in wound-induced gene expression in Arabidopsis. Plant Physiol. 2005, 139, 1268–1283. [Google Scholar] [CrossRef] [Green Version]
- Dhanasekaran, A.; Kotamraju, S.; Karunakaran, C.; Kalivendi, S.V.; Thomas, S.; Joseph, J.; Kalyanaraman, B. Mitochondria superoxide dismutase mimetic inhibits peroxide-induced oxidative damage and apoptosis: Role of mitochondrial superoxide. Free Radic. Biol. Med. 2005, 39, 567–583. [Google Scholar] [CrossRef]
- Garg, T.K.; Chang, J.Y. 15-deoxy-delta 12, 14-prostaglandin J2 prevents reactive oxygen species generation and mitochondrial membrane depolarization induced by oxidative stress. BMC Pharmacol. 2004, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Shibata, T. 15-deoxy-Δ12, 14--Prostaglandin J(2) as an electrophilic mediator. Biosci. Biotechnol. Biochem. 2015, 79, 1044–1049. [Google Scholar] [CrossRef] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Xiao, K.; Zhang, A.; Zhu, W.; Zhang, H.; Tan, F.; Huang, Q.; Wu, X.; Zha, D. Metabolomic Analysis, Combined with Enzymatic and Transcriptome Assays, to Reveal the Browning Resistance Mechanism of Fresh-Cut Eggplant. Foods 2022, 11, 1174. https://doi.org/10.3390/foods11081174
Liu X, Xiao K, Zhang A, Zhu W, Zhang H, Tan F, Huang Q, Wu X, Zha D. Metabolomic Analysis, Combined with Enzymatic and Transcriptome Assays, to Reveal the Browning Resistance Mechanism of Fresh-Cut Eggplant. Foods. 2022; 11(8):1174. https://doi.org/10.3390/foods11081174
Chicago/Turabian StyleLiu, Xiaohui, Kai Xiao, Aidong Zhang, Weimin Zhu, Hui Zhang, Feng Tan, Qianru Huang, Xuexia Wu, and Dingshi Zha. 2022. "Metabolomic Analysis, Combined with Enzymatic and Transcriptome Assays, to Reveal the Browning Resistance Mechanism of Fresh-Cut Eggplant" Foods 11, no. 8: 1174. https://doi.org/10.3390/foods11081174