
Shifts in the Bacterial Community Related to Quality Properties of Vacuum-Packaged Peeled Potatoes during Storage

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction and PCR Amplification
2.3. Illumina MiSeq Sequencing and Processing of Sequencing Data
2.4. Microbial Growth Analysis
2.5. Visual Quality Assessment
2.6. Texture Analysis
2.7. Determination of pH and Organic Acids
2.8. E-Nose
2.9. Determination of VOCs Using Gas Chromatography-Mass Spectrometry (GC-MS)
2.10. Statistical Analysis
3. Results and Discussion
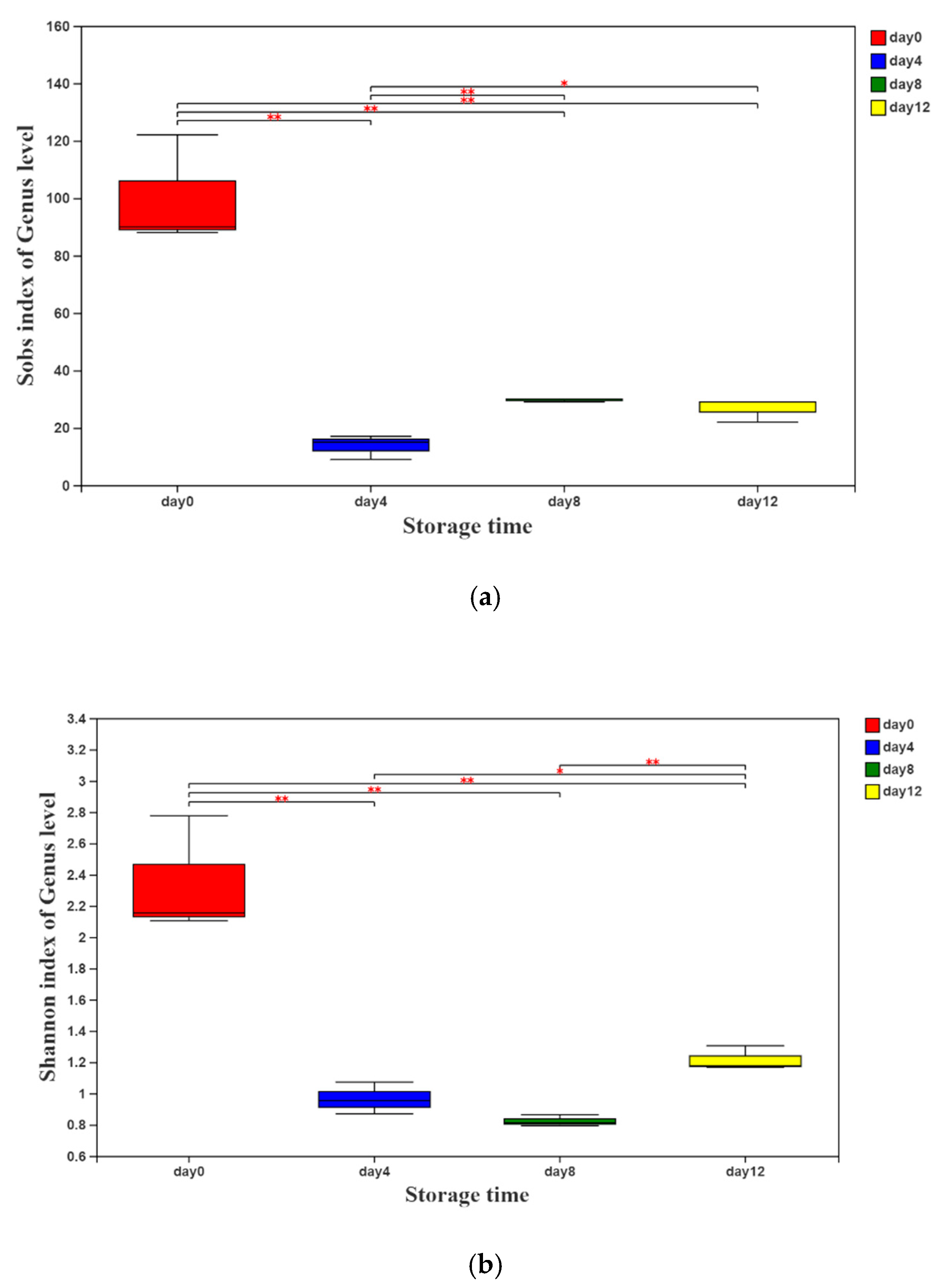
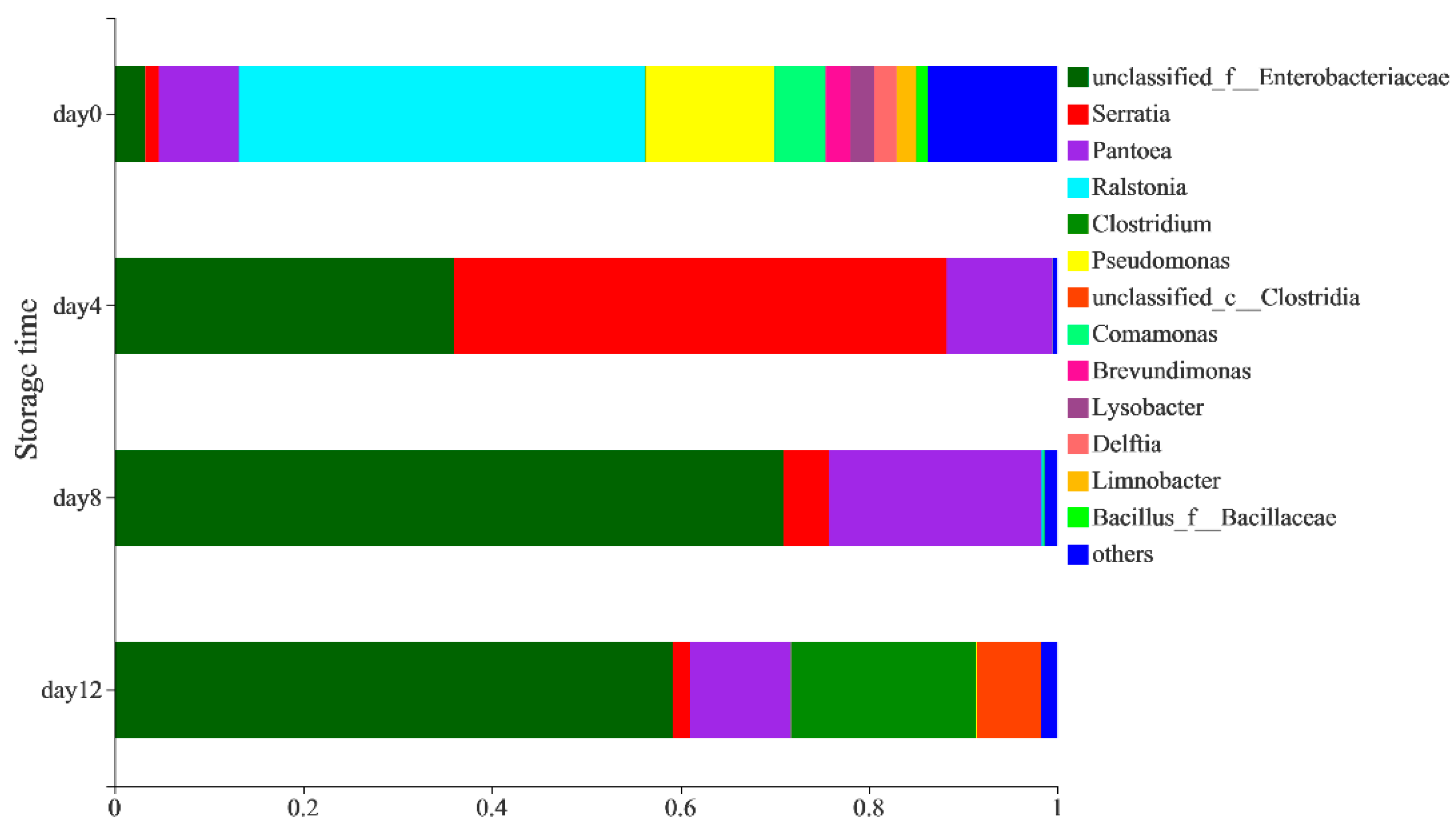
3.1. Bacterial Community
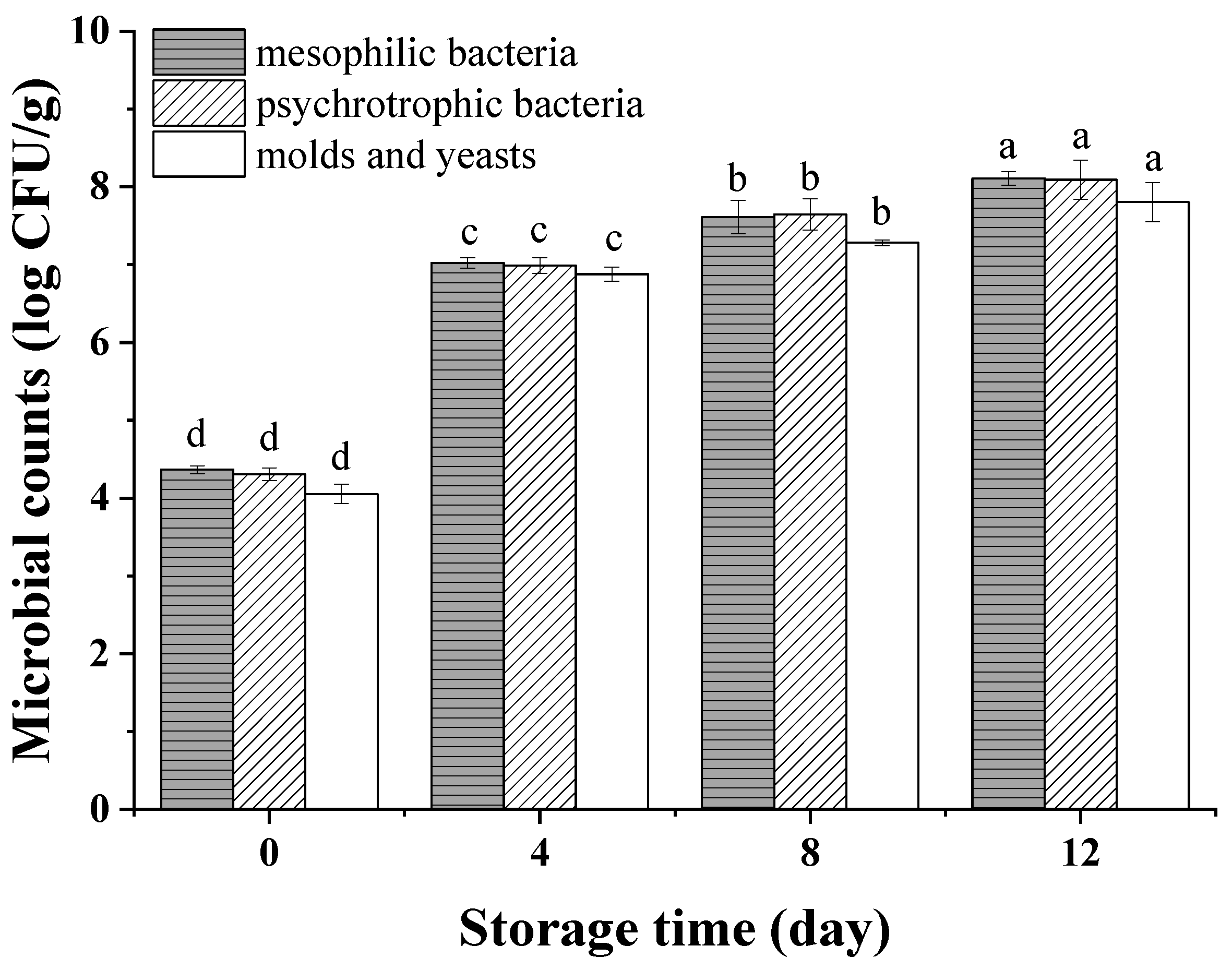
3.2. Microbial Quality
3.3. pH and Organic Acids
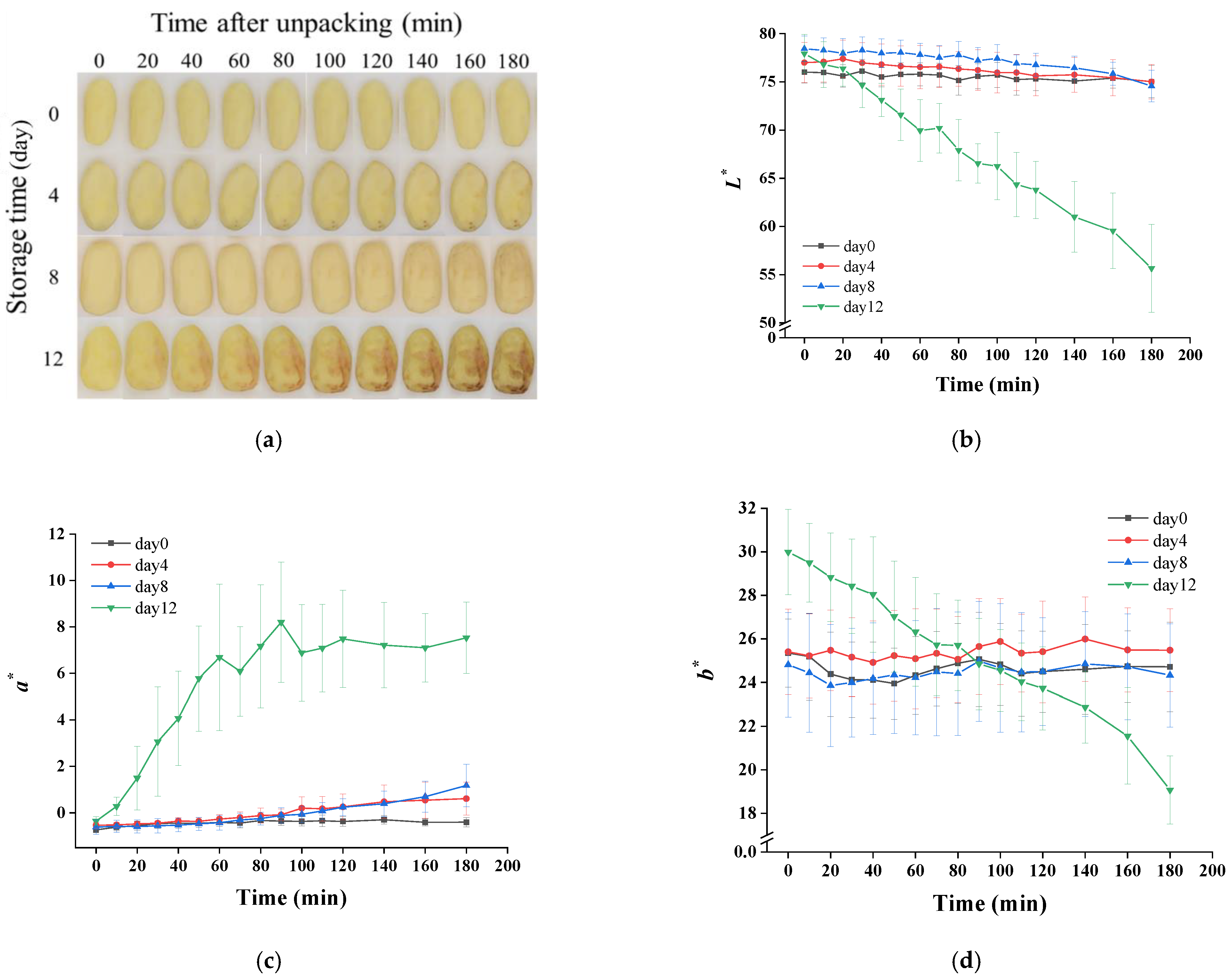
3.4. Visual Quality and Hardness
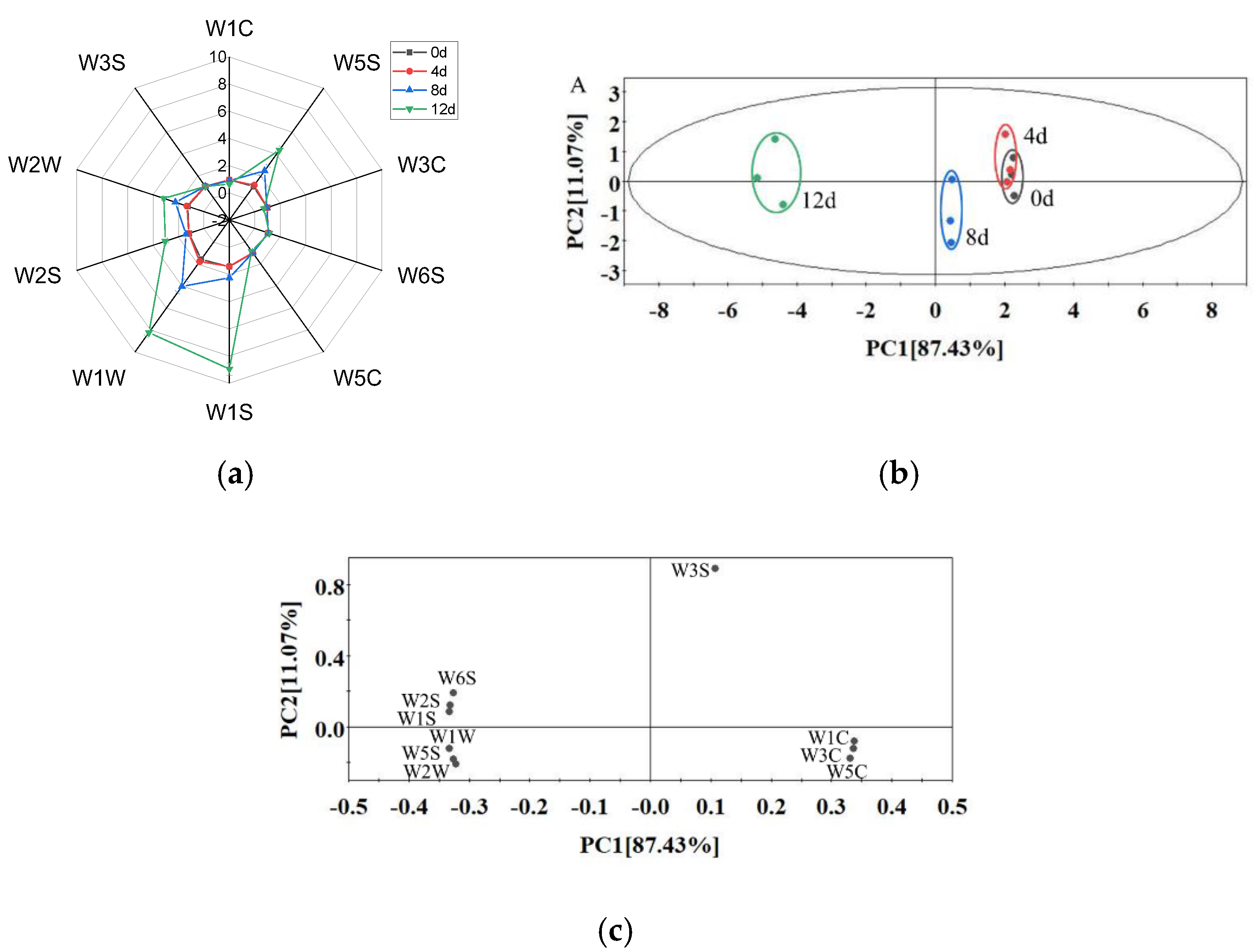
3.5. E-Nose
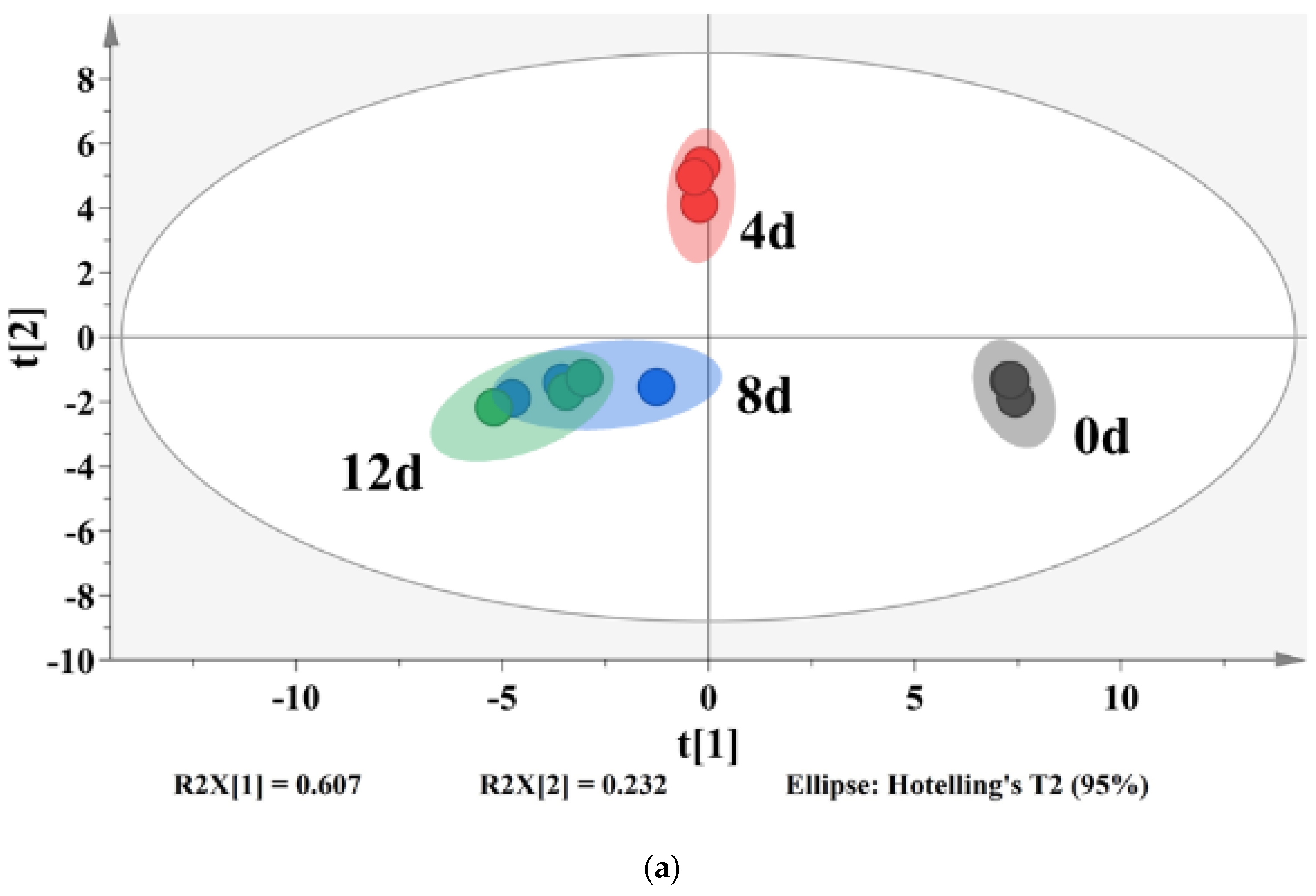
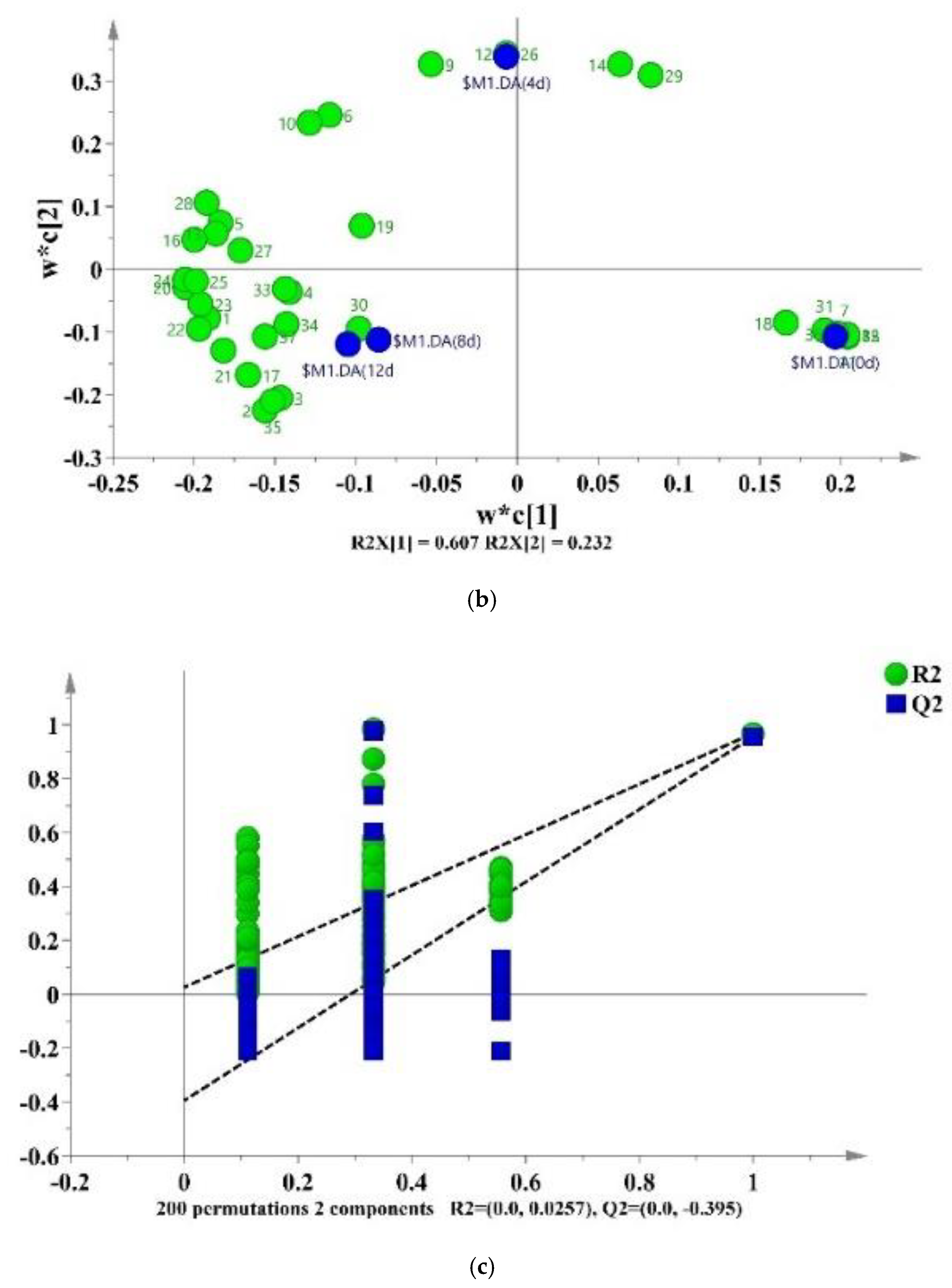
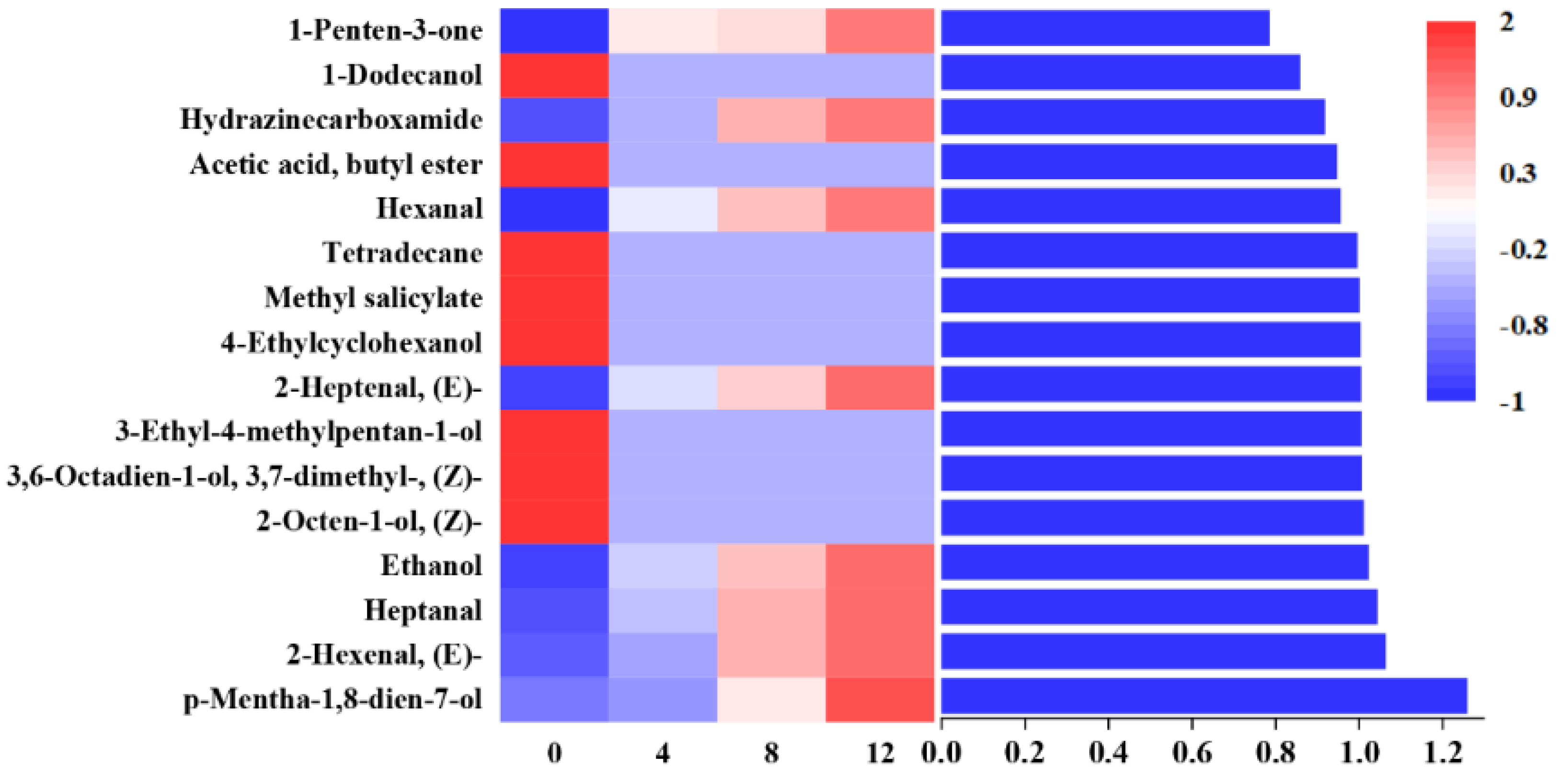
3.6. VOCs
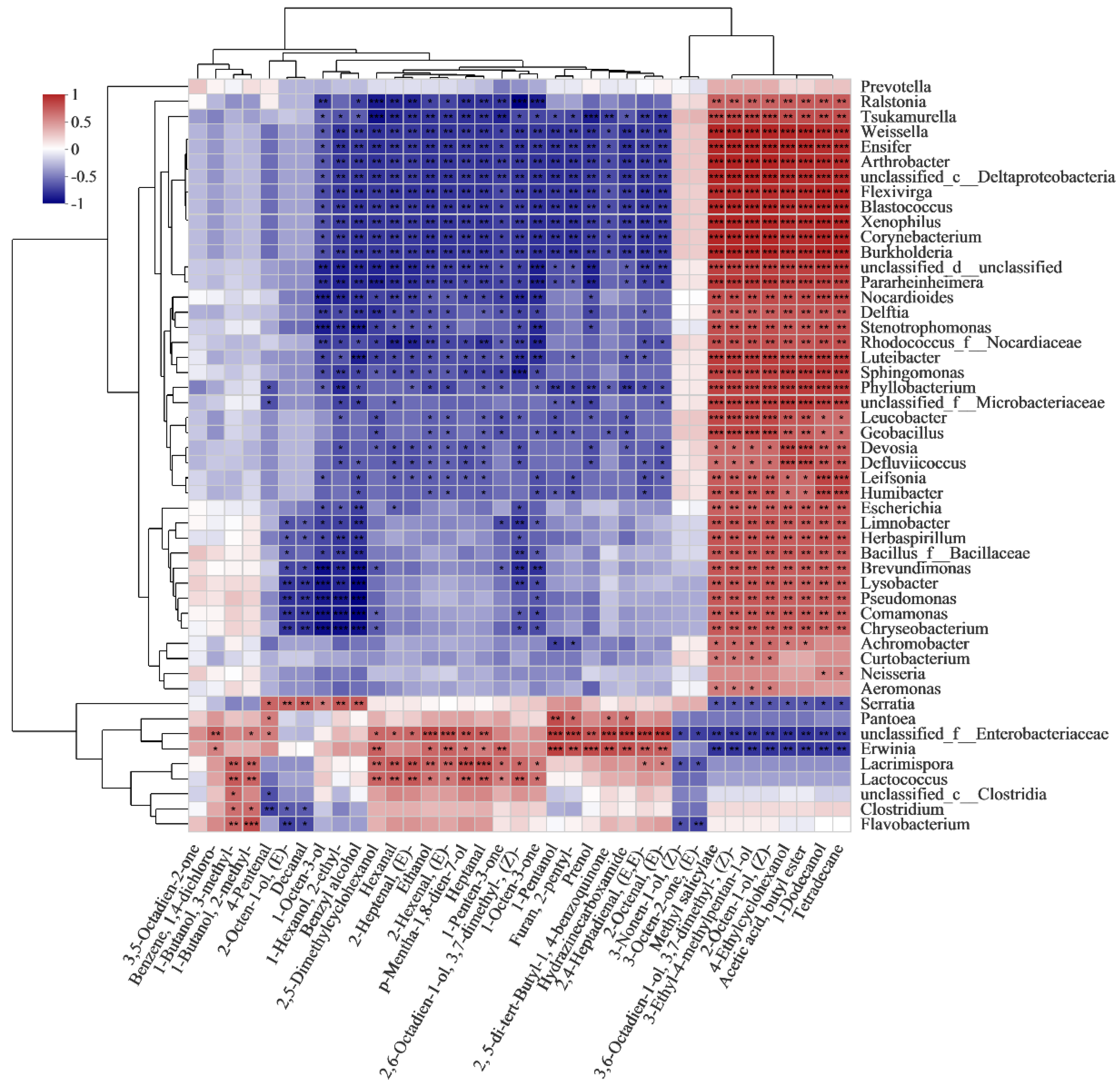
3.7. Correlation Analysis between Bacterial Genera and VOCs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montouto-Grana, M.; Cabanas-Arias, S.; Porto-Fojo, S.; Lourdes Vazquez-Oderiz, M.; Angeles Romero-Rodriguez, M. Sensory characteristics and consumer acceptance and purchase intention toward fresh-cut potatoes. J. Food Sci. 2012, 77, S40–S46. [Google Scholar] [CrossRef] [PubMed]
- Tudela, J.A.; Cantos, E.; Espin, J.C.; Tomas-Barberan, F.A.; Gil, M.I. Induction of antioxidant flavonol biosynthesis in fresh-cut potatoes. Effect of domestic cooking. J. Agric. Food Chem. 2002, 50, 5925–5931. [Google Scholar] [CrossRef] [PubMed]
- Bobo-Garcia, G.; Arroqui, C.; Merino, G.; Virseda, P. Antibrowning compounds for minimally processed potatoes: A review. Food Rev. Int. 2020, 36, 529–546. [Google Scholar] [CrossRef]
- Tsikrika, K.; Tzima, K.; Rai, D.K. Recent advances in anti-browning methods in minimally processed potatoes-A review. J. Food Process. Preserv. 2022, 46, e16298. [Google Scholar] [CrossRef]
- Moon, K.M.; Kwon, E.B.; Lee, B.; Kim, C.Y. Recent trends in controlling the enzymatic browning of fruit and vegetable products. Molecules 2020, 25, 2754. [Google Scholar] [CrossRef]
- Denoya, G.I.; Vaudagna, S.R.; Polenta, G. Effect of high pressure processing and vacuum packaging on the preservation of fresh-cut peaches. LWT 2015, 62, 801–806. [Google Scholar] [CrossRef]
- Min, T.; Liu, E.-C.; Xie, J.; Yi, Y.; Wang, L.-M.; Ai, Y.-W.; Wang, H.-X. Effects of vacuum packaging on enzymatic browning and ethylene response factor (ERF) gene expression of fresh-cut lotus root. HortScience 2019, 54, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Min, T.; Liu, E.-C.; Xie, J.; Yi, Y.; Wang, L.-M.; Ai, Y.-W.; Wang, H.-X. Effects of vacuum packaging on NAC gene expression in fresh-cut lotus root. J. Am. Soc. Hort. Sci. 2020, 145, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Dite Hunjek, D.; Pranjic, T.; Repajic, M.; Levaj, B. Fresh-cut potato quality and sensory: Effect of cultivar, age, processing, and cooking during storage. J. Food Sci. 2020, 85, 2296–2309. [Google Scholar] [CrossRef]
- Ge, L.; Lai, H.; Huang, Y.; Wang, Y.; Li, Y.; Zhu, S.; Shi, Q.; Li, H.; Zhu, Y.; Zhao, N. Comparative evaluation of package types in alleviating textural softening and package-swelling of Paocai during storage: Insight into microbial invasion, cell wall pectinolysis and alteration in sugar and organic acid profiles. Food Chem. 2021, 365, 130489. [Google Scholar] [CrossRef]
- Tudela, J.A.; Hernandez, J.A.; Gil, M.I.; Espin, J.C. L-Galactono-γ-lactone dehydrogenase activity and vitamin C content in fresh-cut potatoes stored under controlled atmospheres. J. Agric. Food Chem. 2003, 51, 4296–4302. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Hong, S.I.; Kim, D. Microbiological and visual quality of fresh-cut cabbage as affected by packaging treatments. Food Sci. Biotechnol. 2011, 20, 229–235. [Google Scholar] [CrossRef]
- Zudaire, L.; Vinas, I.; Abadias, M.; Lafarga, T.; Bobo, G.; Simo, J.; Aguilo-Aguayo, I. Effects of long-term controlled atmosphere storage, minimal processing, and packaging on quality attributes of calcots (Allium cepa L.). Food Sci. Technol. Int. 2020, 26, 403–412. [Google Scholar] [CrossRef]
- Gonzalez-Aguilar, G.A.; Ayala-Zavala, J.F.; Ruiz-Cruz, S.; Acedo-Felix, E.; Diaz-Cinco, M.E. Effect of temperature and modified atmosphere packaging on overall quality of fresh-cut bell peppers. Lebensm. Wiss. Und-Technol. Food Sci. Technol. 2004, 37, 817–826. [Google Scholar] [CrossRef]
- Pothakos, V.; Stellato, G.; Ercolini, D.; Devlieghere, F.; Griffiths, M.W.J.A.; Microbiology, E. Processing environment and ingredients are both sources of Leuconostoc gelidum, which emerges as a major spoiler in ready-to-eat meals. Appl. Environ. Microbiol. 2015, 81, 3529–3541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vihavainen, E.J.; Murros, A.E.; Bjrkroth, K.J. Leuconostoc spoilage of vacuum-packaged vegetable sausages. J. Food Prot. 2008, 71, 2312–2315. [Google Scholar] [CrossRef] [PubMed]
- Rocha, A.M.; Coulon, E.C.; Morais, A.M. Effects of vacuum packaging on the physical quality of minimally processed potatoes. Food Serv. Technol. 2003, 3, 81–88. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura de Sena Aquino, A.C.; Azevedo, M.S.; Baggio Ribeiro, D.H.; Oliveira Costa, A.C.; Amante, E.R. Validation of HPLC and CE methods for determination of organic acids in sour cassava starch wastewater. Food Chem. 2015, 172, 725–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paillart, M.J.M.; van der Vossen, J.M.B.M.; Levin, E.; Lommen, E.; Otma, E.C.; Snels, J.C.M.A.; Woltering, E.J. Bacterial population dynamics and sensorial quality loss in modified atmosphere packed fresh-cut iceberg lettuce. Postharvest Biol. Technol. 2017, 124, 91–99. [Google Scholar] [CrossRef]
- Solomon, H.M.; Rhodehamel, E.J.; Kautter, D.A. Growth and toxin production by clostridium botulinum in sliced raw potatoes under vacuum with and without sulfite. J. Food Prot. 1994, 57, 878–881. [Google Scholar] [CrossRef]
- Avci, A.; Kamiloglu, A.; Donmez, S. Efficient production of acetone butanol ethanol from sole fresh and rotten potatoes by various Clostridium strains. Biomass Convers. Biorefin. 2021, 189, 1–9. [Google Scholar]
- Lauridsen, L.; Knochel, S. Microbiological stability and diversity in raw pre-peeled potatoes packed in different atmospheres. Eur. Food Res. Technol. 2003, 217, 421–426. [Google Scholar] [CrossRef]
- Paillart, M.; Vossen, J.M.B.M.v.d.; Levin, E.; Otma, E.; Lommen, E.; Snels, J.; Woltering, E.J. Organic acids produced by lactic acid bacteria (Leuconostoc sp.) contribute to sensorial quality loss in modified-atmosphere-packed fresh-cut iceberg lettuce. Acta Hortic. 2016, 1141, 289–296. [Google Scholar] [CrossRef]
- Oms-Oliu, G.; Aguilo-Aguayo, I.; Martin-Belloso, O.; Soliva-Fortuny, R. Effects of pulsed light treatments on quality and antioxidant properties of fresh-cut mushrooms (Agaricus bisporus). Postharvest Biol. Technol. 2010, 56, 216–222. [Google Scholar] [CrossRef]
- Waimaleongora-Ek, P.; Corredor, A.J.H.; No, H.K.; Prinyawiwatkul, W.; King, J.M.; Janes, M.E.; Sathivel, S. Selected quality characteristics of fresh-cut sweet potatoes coated with chitosan during 17-day refrigerated storage. J. Food Sci. 2008, 73, S418–S423. [Google Scholar] [CrossRef]
- Zhai, Y.; Perez-Diaz, I.M.; Diaz, J.T. Viability of commercial cucumber fermentation without nitrogen or air purging. Trends Food Sci. Technol. 2018, 81, 185–192. [Google Scholar] [CrossRef]
- Pothimon, R.; Podjanee, U.; Krusong, W.; Thompson, A.K.; Massa, S. Inhibition of Pantoea agglomerans contamination of fresh-cut jackfruit by exposure to weak organic acid vapors. LWT 2021, 139, 110586. [Google Scholar] [CrossRef]
- Corbo, M.R.; del Nobile, M.A.; Sinigaglia, M. A novel approach for calculating shelf life of minimally processed vegetables. Int. J. Food Microbiol. 2006, 106, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Jacques, M.A.; Morris, C.E. Bacterial population dynamics and decay on leaves of different ages of ready-to-use broad-leaved endive. Int. J. Food Sci. Tech. 2010, 30, 221–236. [Google Scholar] [CrossRef]
- Ragaert, P.; Devlieghere, F.; Debevere, J. Role of microbiological and physiological spoilage mechanisms during storage of minimally processed vegetables. Postharvest Biol. Technol. 2007, 44, 185–194. [Google Scholar] [CrossRef]
- Robbs, P.G.; Bartz, J.A.; McFie, G.; Hodge, N.C. Causes of decay of fresh-cut celery. J. Food Sci. 1996, 61, 444–448. [Google Scholar] [CrossRef]
- Xiong, T.; Li, J.; Liang, F.; Wang, Y.; Guan, Q.; Xie, M. Effects of salt concentration on Chinese sauerkraut fermentation. LWT 2016, 69, 169–174. [Google Scholar] [CrossRef]
- Huang, S.J.; Lin, S.Y.; Wang, T.T.; Hsu, F.C. Combining acetic acid and ethanol as an anti-browning treatment for lettuce butt discoloration through repression of the activity and expression of phenylalanine ammonia lyase. Postharvest Biol. Technol. 2020, 164, 111151. [Google Scholar] [CrossRef]
- Shabuer, G.; Ishida, K.; Pidot, S.J.; Roth, M.; Dahse, H.-M.; Hertweck, C. Plant pathogenic anaerobic bacteria use aromatic polyketides to access aerobic territory. Science 2015, 350, 670–674. [Google Scholar] [CrossRef]
- Supapvanich, S.; Tucker, G.A. Physicochemical changes in fresh-cut Honeydew melon fruit during storage. Afr. J. Agric. Res. 2011, 6, 2737–2742. [Google Scholar]
- Zhang, B.Y.; Samapundo, S.; Pothakos, V.; Surengil, G.; Devlieghere, F. Effect of high oxygen and high carbon dioxide atmosphere packaging on the microbial spoilage and shelf-life of fresh-cut honeydew melon. Int. J. Food Microbiol. 2013, 166, 378–390. [Google Scholar] [CrossRef] [PubMed]
- El-Hendawy, H.H.; Osman, M.E.; Ramadan, H.A. Pectic enzymes produced in vitro and in vivo by Erwinia spp. isolated from carrot and pepper in Egypt. J. Phytopathol. Phytopathol. 2002, 150, 431–438. [Google Scholar] [CrossRef]
- Chen, H.Z.; Zhang, M.; Bhandari, B.; Guo, Z. Evaluation of the freshness of fresh-cut green bell pepper (Capsicum annuum var. grossum) using electronic nose. LWT 2018, 87, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Jacxsens, L.; Devlieghere, F.; Ragaert, P.; Vanneste, E.; Debevere, J. Relation between microbiological quality, metabolite production and sensory quality of equilibrium modified atmosphere packaged fresh-cut produce. Int. J. Food Microbiol. 2003, 83, 263–280. [Google Scholar] [CrossRef]
- Chen, H.Z.; Zhang, M.; Guo, Z. Discrimination of fresh-cut broccoli freshness by volatiles using electronic nose and gas chromatography-mass spectrometry. Postharvest Biol. Technol. 2019, 148, 168–175. [Google Scholar] [CrossRef]
- Dresow, J.F.; Boehm, H. The influence of volatile compounds of the flavour of raw, boiled and baked potatoes: Impact of agricultural measures on the volatile components. Landbauforsch. Volkenrode 2009, 59, 309–337. [Google Scholar]
- Beaulieu, J.C. Within-season volatile and quality differences in stored fresh-cut cantaloupe cultivars. J. Agric. Food Chem. 2005, 53, 8679–8687. [Google Scholar] [CrossRef]
- Lamikanra, O.; Richard, O.A. Effect of storage on some volatile aroma compounds in fresh-cut cantaloupe melon. J. Agric. Food Chem. 2002, 50, 4043–4047. [Google Scholar] [CrossRef]
- Costello, B.P.J.D.L.; Evans, P.; Ewen, R.J.; Gunson, H.E.; Spencer-Phillips, P.T.N. Gas chromatography-mass spectrometry analyses of volatile organic compounds from potato tubers inoculated with Phytophthora infestans or Fusarium coeruleum. Plant Pathol. 2010, 50, 489–496. [Google Scholar] [CrossRef]
- Petersen, M.A.; Poll, L.; Larsen, L.M. Comparison of volatiles in raw and boiled potatoes using a mild extraction technique combined with GC odour profiling and GC-MS. Food Chem. 1998, 61, 461–466. [Google Scholar] [CrossRef]
- Waterer, D.R.; Pritchard, M.K. Monitoring of volatiles: A technique for detection of soft rot (Erwinia carotovora) in potato tubers. Can. J. Plant Pathol. 1984, 6, 165–171. [Google Scholar] [CrossRef]
- Schütz, S.; Weißbecker, B.; Koch, U.T.; Hummel, H.E. Detection of volatiles released by diseased potato tubers using a biosensor on the basis of intact insect antennae. Biosens. Bioelectron. 1999, 14, 221–228. [Google Scholar] [CrossRef]
- Vollbrecht, D.; Radler, F. Die bildung höherer alkohole bei aminosäuremangelmutanten von saccharomyces cerevisiae III. höhere alkohole als nebenprodukte der aminosäuresynthese. Zent. Für Bakteriol. Parasitenkd. Infekt. Und Hygiene. Zweite Nat. Abt. Allg. Landwirtsch. Und Tech. Mikrobiol. 1975, 130, 238–244. [Google Scholar] [CrossRef]
- Ragaert, P.; Devlieghere, F.; Devuyst, E.; Dewulf, J.; van Langenhove, H.; Debevere, J. Volatile metabolite production of spoilage micro-organisms on a mixed-lettuce agar during storage at 7 degrees C in air and low oxygen atmosphere. Int. J. Food Microbiol. 2006, 112, 162–170. [Google Scholar] [CrossRef]
- Ioannidis, A.G.; Kerckhof, F.M.; Drif, Y.R.; Vanderroost, M.; Boon, N.; Ragaert, P.; de Meulenaer, B.; Devlieghere, F. Characterization of spoilage markers in modified atmosphere packaged iceberg lettuce. Int. J. Food Microbiol. 2018, 279, 1–13. [Google Scholar] [CrossRef]
- Lopez-Galvez, F.; Ragaert, P.; Hague, M.A.; Eriksson, M.; van Labeke, M.C.; Devlieghere, F. High oxygen atmospheres can induce russet spotting development in minimally processed iceberg lettuce. Postharvest Biol. Technol. 2015, 100, 168–175. [Google Scholar] [CrossRef]
- Rao, Y.; Tao, Y.; Chen, X.; She, X.; Qian, Y.; Li, Y.; Du, Y.; Xiang, W.; Li, H.; Liu, L. The characteristics and correlation of the microbial communities and flavors in traditionally pickled radishes. LWT 2020, 118, 108804. [Google Scholar] [CrossRef]
- Tiwari, S.; Goswami, U.; Kate, A.; Modhera, B.; Tripathi, M.K.; Mohapatra, D. Biological relevance of VOCs emanating from red onions infected with Erwinia (Pectobacterium) carotovora under different storage conditions. Postharvest Biol. Technol. 2022, 184, 111761. [Google Scholar] [CrossRef]
- Ercolini, D.; Russo, F.; Nasi, A.; Ferranti, P.; Villani, F. Mesophilic and psychrotrophic bacteria from meat and their spoilage potential in vitro and in beef. Appl. Environ. Microbiol. 2009, 75, 1990–2001. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zhang, F.; Zhang, C.; Yang, L.; Fan, G.; Xu, Y.; Sun, B.; Li, X. Dynamic microbial succession of Shanxi aged vinegar and its correlation with flavor metabolites during different stages of acetic acid fermentation. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Morales, P.; Feliu, I.; Fernandez-Garcia, E.; Nunez, M. Volatile compounds produced in cheese by Enterobacteriaceae strains of dairy origin. J. Food Prot. 2004, 67, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Garde, S.; Carbonell, M.; Fernández-García, E.; Medina, M.; Nuñez, M. Volatile compounds in Hispánico cheese manufactured using a mesophilic starter, a thermophilic starter, and bacteriocin-producing Lactococcus lactis subsp. lactis INIA 415. J. Agric. Food Chem. 2002, 50, 6752–6757. [Google Scholar] [CrossRef] [PubMed]
- Gallegos, J.; Arce, C.; Jordano, R.; Arce, L.; Medina, L.M. Target identification of volatile metabolites to allow the differentiation of lactic acid bacteria by gas chromatography-ion mobility spectrometry. Food Chem. 2017, 220, 362–370. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Storage Time (Day) | pH | Lactic Acid (mg/g FW) | Acetic Acid (mg/g FW) | Hardness (N) | Browning Time (min) |
|---|---|---|---|---|---|
| 0 | 5.91 ± 0.05 a | 0.04 ± 0.01 d | 2.06 ± 0.07 c | 37.76 ± 5.53 a | 31.88 ± 6.31 b |
| 4 | 5.73 ± 0.04 b | 0.24 ± 0.04 c | 2.20 ± 0.08 c | 37.53 ± 3.62 a | 35.50 ± 2.84 a |
| 8 | 5.67 ± 0.06 bc | 0.45 ± 0.03 b | 3.39 ± 0.65 b | 37.13 ± 3.96 a | 20.35 ± 3.14 c |
| 12 | 5.60 ± 0.08 c | 0.79 ± 0.09 a | 4.87 ± 0.06 a | 31.55 ± 6.70 b | 5.71 ± 1.29 d |
| Group (NO.) | Volatile Compounds a | Retention Index b | The Content of Volatile Organic Compounds in Different Storage Times (μg/kg) c | |||
|---|---|---|---|---|---|---|
| 0 Day | 4 Day | 8 Day | 12 Day | |||
| Alcohols | ||||||
| 1 | Ethanol | - | - | 1517.10 ± 382.57 | 2795.72 ± 757.98 | 3843.27 ± 1004.55 |
| 2 | 1-Butanol, 3-methyl- | 732.64 | 80.46 ± 3.32 | 61.94 ± 5.31 | 190.90 ± 13.3 | 236.17 ± 22.47 |
| 3 | 1-Butanol, 2-methyl- | 736.09 | 76.96 ± 7.02 | 60.19 ± 8.35 | 181.93 ± 32.18 | 246.07 ± 61.24 |
| 4 | 1-Pentanol | 764.38 | 52.70 ± 1.02 | 79.57 ± 2.77 | 124.41 ± 34.94 | 80.63 ± 8.14 |
| 5 | Prenol | 771.74 | 29.81 ± 6.57 | 52.00 ± 2.83 | 55.88 ± 10.13 | 51.61 ± 0.23 |
| 6 | 1-Octen-3-ol | 979.72 | 307.61 ± 9.78 | 500.66 ± 27.33 | 366.60 ± 48.56 | 436.94 ± 29.16 |
| 7 | 4-Ethylcyclohexanol | 992.22 | 9.95 ± 1.42 | - | - | - |
| 8 | 3-Ethyl-4-methylpentan-1-ol | 1019.97 | 6.72 ± 0.44 | - | - | - |
| 9 | 1-Hexanol, 2-ethyl- | 1027.86 | 130.56 ± 9.05 | 303.64 ± 27.78 | 167.53 ± 13.04 | 165.62 ± 5.27 |
| 10 | Benzyl alcohol | 1032.39 | 26.21 ± 9.04 | 87.59 ± 18.30 | 54.72 ± 12.72 | 65.00 ± 4.75 |
| 11 | 2-Octen-1-ol, (Z)- | 1066.09 | 5.55 ± 0.02 | - | - | - |
| 12 | 2-Octen-1-ol, (E)- | 1069.78 | - | 20.19 ± 1.62 | - | - |
| 13 | 2,5-Dimethylcyclohexanol | 1093.29 | 11.12 ± 1.65 | 37.59 ± 6.66 | 35.81 ± 11.93 | 46.57 ± 5.82 |
| 14 | 3-Nonen-1-ol, (Z)- | 1152.67 | 6.01 ± 0.50 | 18.33 ± 1.03 | - | - |
| 15 | 3,6-Octadien-1-ol, 3,7-dimethyl-, (Z)- | 1239.06 | 4.99 ± 0.31 | - | - | - |
| 16 | 2,6-Octadien-1-ol, 3,7-dimethyl-, (Z)- | 1248.75 | 22.63 ± 0.10 | 48.16 ± 1.16 | 45.75 ± 1.34 | 60.51 ± 2.85 |
| 17 | p-Mentha-1,8-dien-7-ol | 1305.40 | 2.01 ± 0.43 | 28.49 ± 1.45 | 48.57 ± 2.00 | 71.81 ± 1.93 |
| 18 | 1-Dodecanol | 1475.17 | 45.25 ± 34.70 | - | - | - |
| The total content of alcohols (μg/kg) | 841.52 ± 32.81 | 2815.46 ± 385.70 | 4067.82 ± 892.95 | 5304.19 ± 1075.33 | ||
| Aldehydes | ||||||
| 19 | 4-Pentenal | 751.28 | - | 19.60 ± 3.48 | 37.00 ± 3.59 | - |
| 20 | Hexanal | 801.71 | 165.48 ± 7.30 | 1505.03 ± 78.26 | 2070.49 ± 543.93 | 2632.31 ± 87.91 |
| 21 | 2-Hexenal, (E)- | 851.48 | - | 39.32 ± 12.80 | 122.19 ± 24.57 | 165.66 ± 50.46 |
| 22 | Heptanal | 901.47 | - | 18.13 ± 0.47 | 39.28 ± 11.57 | 49.46 ± 3.41 |
| 23 | 2-Heptenal, (E)- | 956.01 | 11.23 ± 0.75 | 178.43 ± 17.41 | 269.39 ± 84.64 | 389.63 ± 52.24 |
| 24 | 2,4-Heptadienal, (E, E)- | 995.28 | 12.61 ± 4.72 | 126.22 ± 9.29 | 191.98 ± 48.02 | 186.38 ± 19.20 |
| 25 | 2-Octenal, (E)- | 1057.55 | 12.65 ± 1.42 | 152.55 ± 8.74 | 249.89 ± 73.72 | 214.86 ± 22.23 |
| 26 | Decanal | 1205.13 | - | 19.56 ± 1.73 | - | - |
| The total content of aldehydes (μg/kg) | 201.97 ± 11.08 | 2058.83 ± 118.58 | 2980.22 ± 785.10 | 3638.29 ± 21.61 | ||
| Ketones | ||||||
| 27 | 1-Penten-3-one | 683.26 | 15.15 ± 13.43 | 47.63 ± 21.40 | 49.33 ± 4.55 | 62.38 ± 15.28 |
| 28 | 1-Octen-3-one | 976.29 | 43.33 ± 2.45 | 109.84 ± 11.14 | 99.48 ± 13.54 | 117.10 ± 11.62 |
| 29 | 3-Octen-2-one, (E)- | 1037.08 | 12.95 ± 3.75 | 30.54 ± 5.59 | - | - |
| 30 | 3,5-Octadien-2-one | 1068.29 | 42.17 ± 1.74 | 45.09 ± 10.65 | 71.59 ± 15.95 | 49.86 ± 15.61 |
| The total content of ketones (μg/kg) | 113.60 ± 11.39 | 233.09 ± 10.67 | 220.41 ± 28.23 | 229.35 ± 30.40 | ||
| Esters | ||||||
| 31 | Acetic acid, butyl ester | 814.23 | 8.20 ± 3.51 | - | - | - |
| 32 | Methyl salicylate | 1191.07 | 9.72 ± 0.91 | - | - | - |
| The total content of esters (μg/kg) | 17.92 ± 2.89 | - | - | - | ||
| Furans | ||||||
| 33 | Furan, 2-pentyl- | 989.12 | 37.65 ± 6.46 | 73.43 ± 6.78 | 130.56 ± 32.65 | 71.25 ± 15.11 |
| Hydrocarbons | ||||||
| 34 | Hydrazinecarboxamide | - | 789.47 ± 1.57 | 910.03 ± 34.95 | 1110.74 ± 70.22 | 1180.93 ± 365.58 |
| 35 | Benzene, 1,4-dichloro- | 1013.07 | 37.88 ± 1.28 | 34.76 ± 2.35 | 74.06 ± 12.30 | 66.69 ± 5.98 |
| 36 | Tetradecane | 1399.22 | 6.98 ± 1.99 | - | - | - |
| 37 | 2, 5-di-tert-Butyl-1, 4-benzoquinone | 1459.44 | 117.87 ± 16.29 | 135.48 ± 26.11 | 186.16 ± 22.54 | 167.97 ± 22.58 |
| The total content of hydrocarbons (μg/kg) | 952.20 ± 19.19 | 1080.26 ± 14.40 | 1370.96 ± 100.07 | 1415.60 ± 393.82 | ||
| The total content of VOCs (μg/kg) | 2164.85 ± 27.65 | 6261.08 ± 484.63 | 8769.97 ± 1768.05 | 10658.68 ± 1467.09 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zhao, W.; Ma, Y.; Liang, H.; Wang, D.; Zhao, X. Shifts in the Bacterial Community Related to Quality Properties of Vacuum-Packaged Peeled Potatoes during Storage. Foods 2022, 11, 1147. https://doi.org/10.3390/foods11081147
Li Z, Zhao W, Ma Y, Liang H, Wang D, Zhao X. Shifts in the Bacterial Community Related to Quality Properties of Vacuum-Packaged Peeled Potatoes during Storage. Foods. 2022; 11(8):1147. https://doi.org/10.3390/foods11081147
Chicago/Turabian StyleLi, Zudi, Wenting Zhao, Yue Ma, Hao Liang, Dan Wang, and Xiaoyan Zhao. 2022. "Shifts in the Bacterial Community Related to Quality Properties of Vacuum-Packaged Peeled Potatoes during Storage" Foods 11, no. 8: 1147. https://doi.org/10.3390/foods11081147