
Characterisation of Lactobacillus plantarum of Dairy-Product Origin for Probiotic Chèvre Cheese Production

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Potential Probiotic Bacteria from Dairy and Fermented Foods
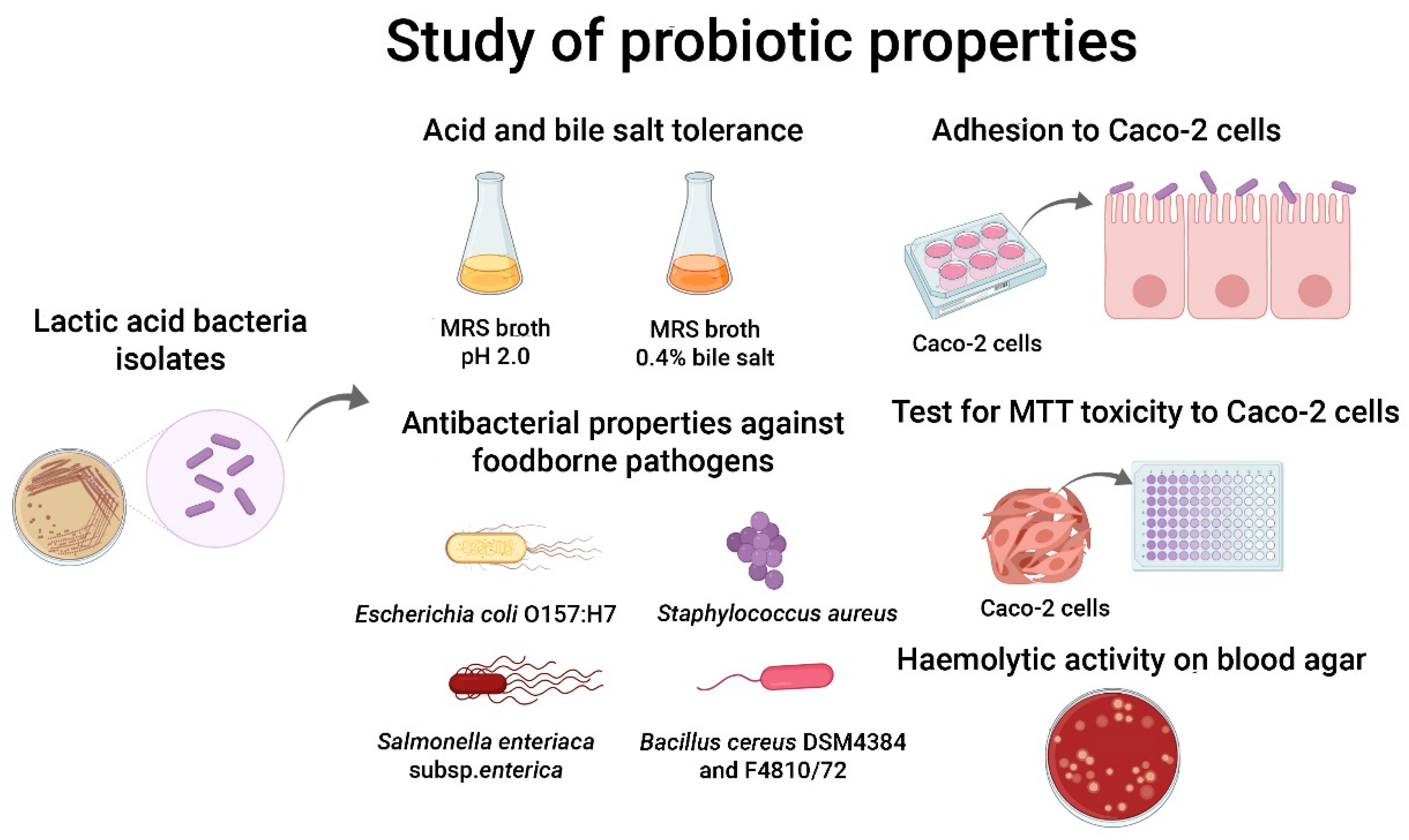
2.2. Investigation of Probiotic Properties of Isolates
2.2.1. Acid Tolerance
2.2.2. Bile Salt Tolerance
2.2.3. Haemolytic Activity Test
2.2.4. Adherence to Caco-2 Cells
2.2.5. Antibacterial Activity Test
2.2.6. Test for Toxicity of Bacterial Isolates to Caco-2 Cells
2.3. Characterisation and Identification of Bacterial Isolates
2.4. Preparation of Probiotic Culture
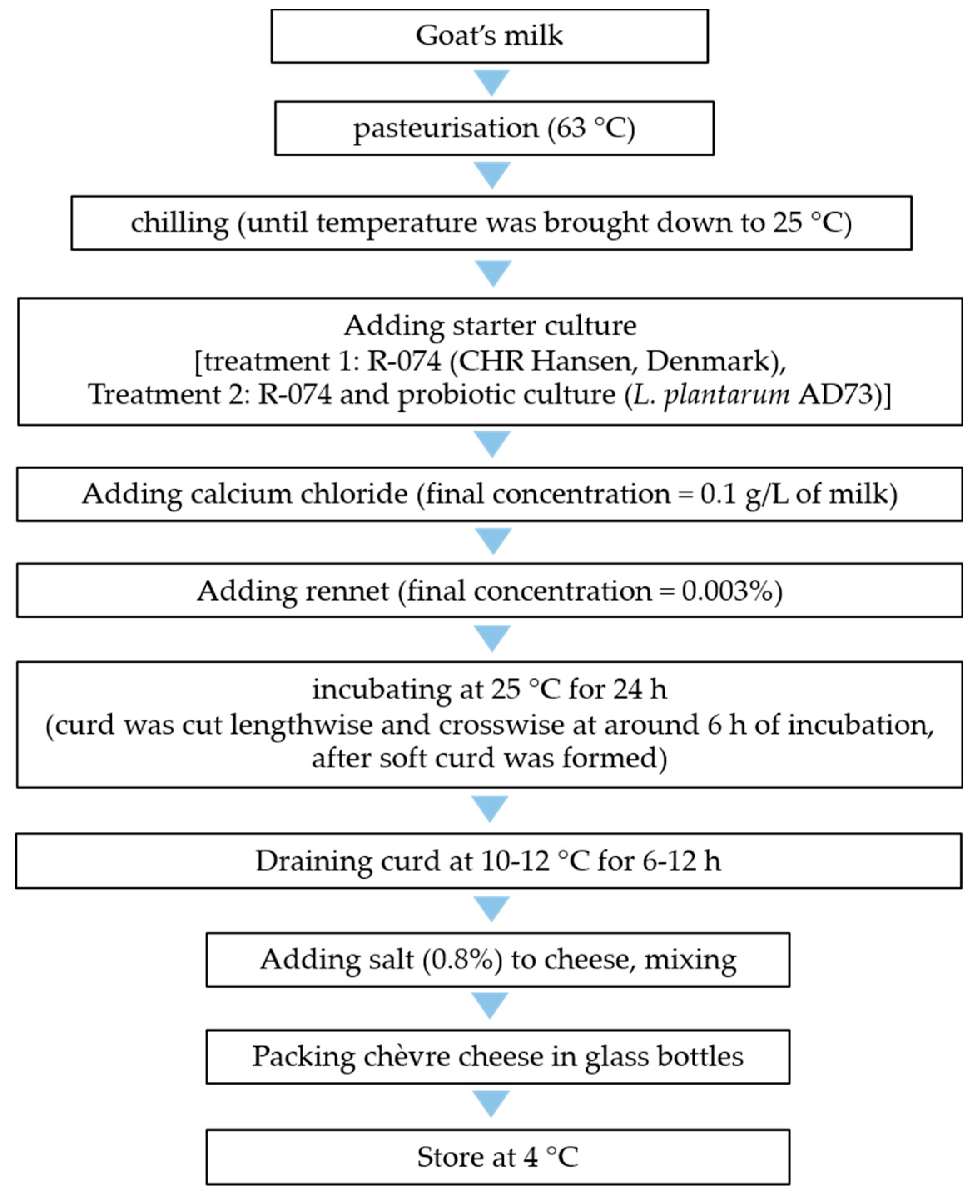
2.5. Production of Probiotic Chèvre Cheese
2.6. Moisture Content and pH Analysis
2.7. Microbiological Analysis of Chèvre Cheese
2.8. Statistical Analysis
3. Results
3.1. Isolation of Potential Probiotic Bacteria from Dairy and Fermented Foods
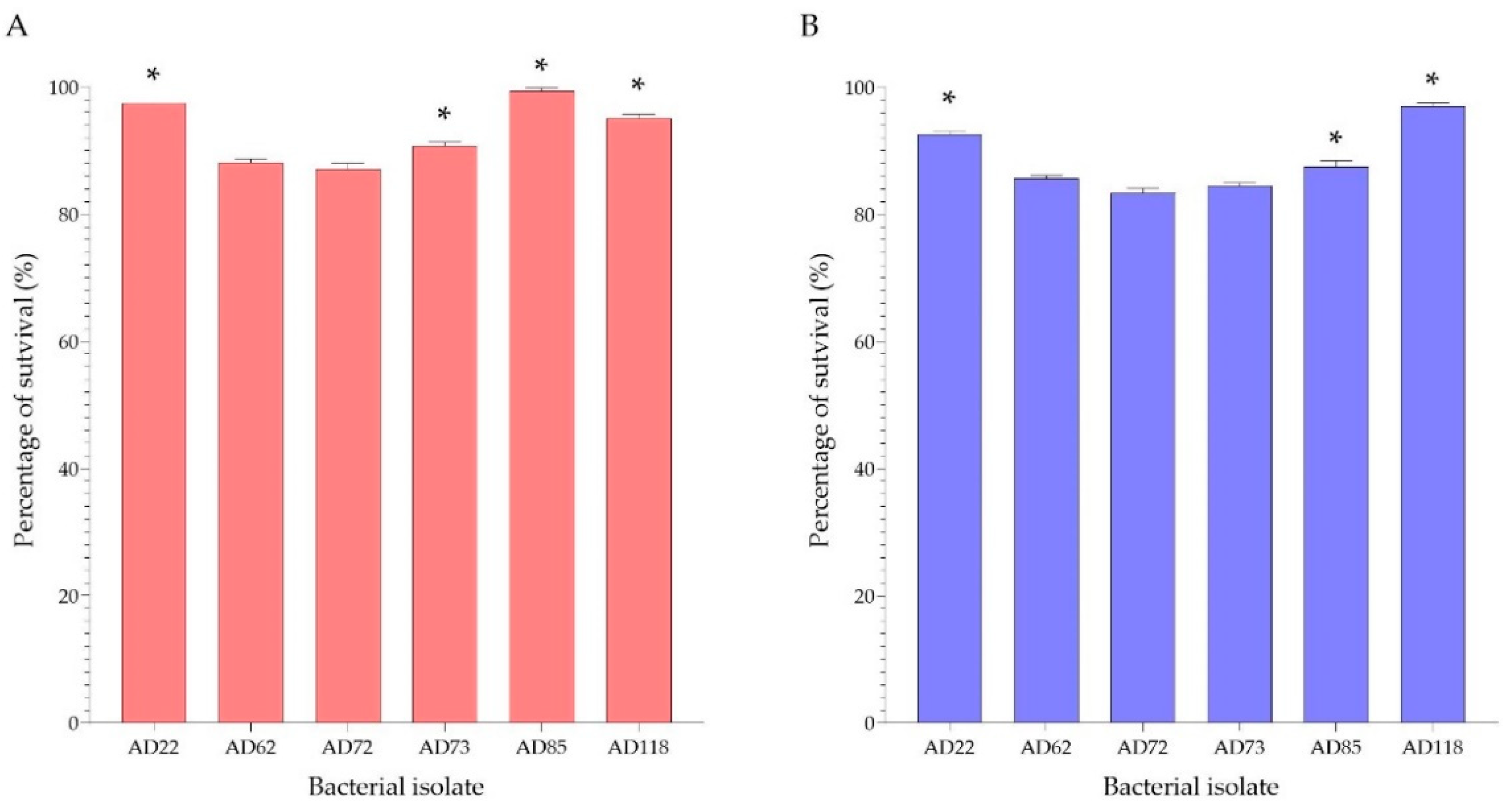
3.2. Acid and Bile Salt Tolerance
3.3. Carbohydrate Fermentation Profiles of Potential Probiotic Isolates
3.4. Antibacterial Activity Test
3.5. Adhesion of Bacterial Isolates on Caco-2 Epithelial Cells
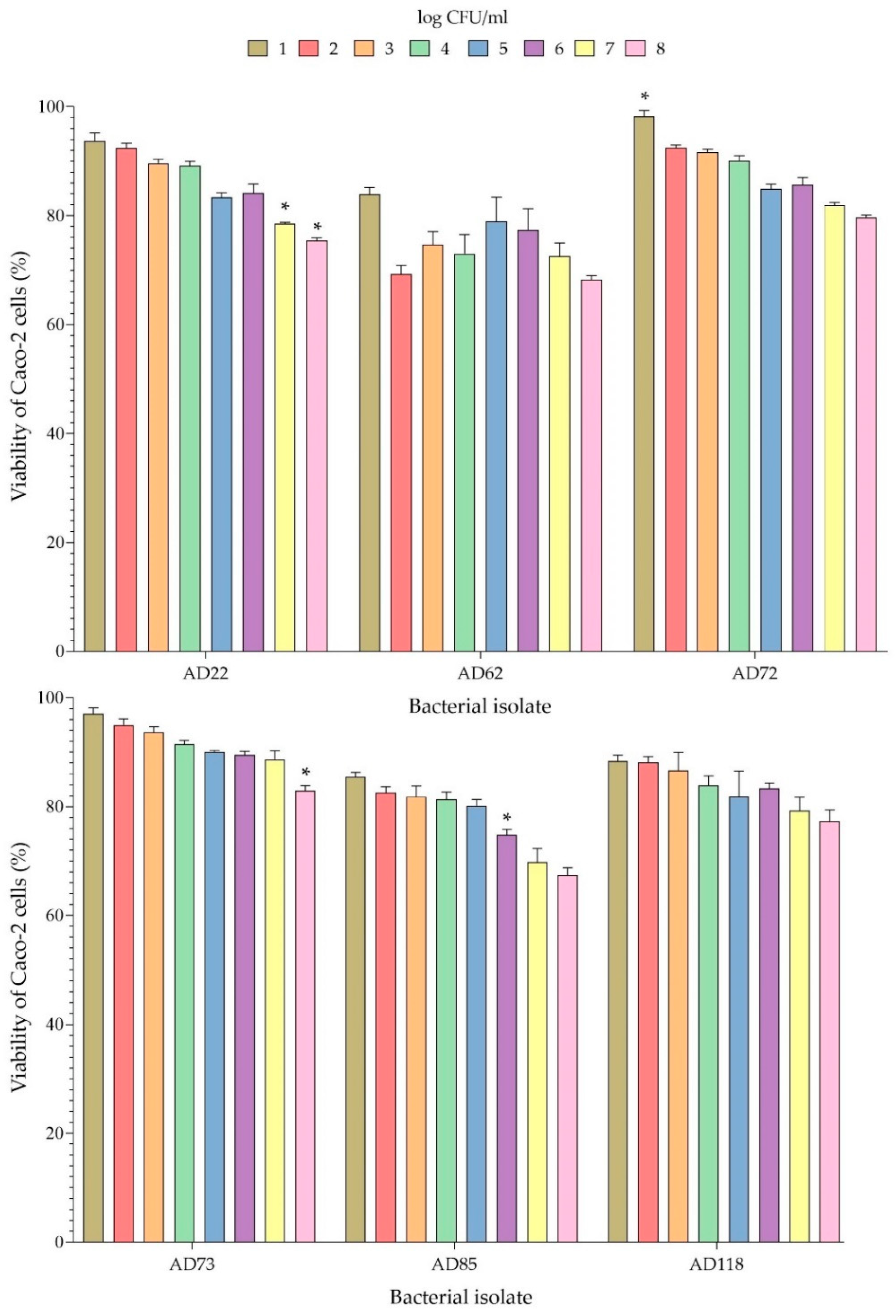
3.6. Cytotoxicity to Caco-2 Cells
3.7. Use of Selected Probiotic Culture for Production of Probiotic Chèvre Cheese
3.7.1. Selection and Preparation of Probiotic Bacteria
3.7.2. Production of Chèvre Cheese
3.8. Microbiological Analysis of Chèvre Cheese
4. Discussion
4.1. Isolation of Potential Probiotic Bacteria, Morphological, Biochemical and Identification of Isolates
4.2. Acid and Bile Salt Tolerance
4.3. Analysis of Properties of Probiotics of Bacterial Isolates with High Probiotic Potentials
4.4. Use of Probiotic Culture in Chèvre Cheese Production
4.5. Survival of Probiotic and Shelf-Life Evaluation of Chèvre Cheese
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.; Cook, B.; Roux, S. The challenges in commercialisation of probiotic API manufacturing. Microbiol. Aust. 2020, 41, 82–85. [Google Scholar]
- Granato, D.; Branco, G.F.; Nazzaro, F.; Cruz, A.G.; Faria, J.A.F. Functional foods and nondairy probiotic food development: Trends, concepts, and products. Compr. Rev. Food Sci. Food Saf. 2010, 9, 292–302. [Google Scholar]
- Stanton, C.; Ross, R.P.; Fitzgerald, G.F.; Van Sinderen, D. Fermented functional foods based on probiotics and their biogenic metabolites. Curr. Opin. Biotechnol. 2005, 16, 198–203. [Google Scholar] [PubMed]
- FAO/WHO. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; FAO/WHO: Rome, Italy, 2006; pp. 1–56. Available online: https://www.fao.org/publications/card/en/c/7c102d95-2fd5-5b22-8faf-f0b2e68dfbb6/ (accessed on 27 February 2022).
- Sanders, M.E.; Guarner, F.; Guerrant, R.; Holt, P.R.; Quigley, E.M. An update on the use and investigation of probiotics in health and disease. Gut 2013, 62, 787–796. [Google Scholar] [PubMed]
- Fontana, L.; Brito, M.B.; Diaz, J.P.; Quezada, S.M.; Gil, A. Sources, isolation, characterization and evaluation of probiotics. Br. J. Nutr. 2013, 109, 35–50. [Google Scholar]
- Pelletier, X.; Laure, B.S.; Donazzolo, Y. Hydrogen excretion upon ingestion of dairy products in lactose-intolerant male subjects: Importance of the live flora. Eur. J. Clin. Nutr. 2001, 55, 509–512. [Google Scholar]
- Woodard, G.A.; Encarnacion, B.; Downey, J.R.; Peraza, J.; Chong, K. Probiotics improve outcomes after Rouxen-Y gastric bypass surgery: A prospective randomized trial. J. Gastrointest. Surg. 2009, 13, 1198–1204. [Google Scholar] [PubMed]
- Karska, W.B.; Bazo, M.; Smoragiewicz, W. Antibacterial activity of Lactobacillus acidophilus and Lactobacillus casei against Methicillin-Resistant Staphylococcus aureus (MRSA). Microbiol. Res. 2010, 165, 674–686. [Google Scholar]
- Rafter, J.; Bennett, M.; Caderni, G.; Clune, Y.; Hughes, R. Dietary synbiotics reduce cancer risk factors in polypectomized and colon cancer patients. Am. J. Clin. Nutr. 2007, 85, 488–496. [Google Scholar]
- Liong, M.T.; Shah, N.P. Acid and bile tolerance and cholesterol removal ability of lactobacilli strains. J. Dairy Sci. 2005, 88, 55–66. [Google Scholar]
- Reid, G.; Sanders, M.E.; Gaskins, H.R.; Gibson, G.R.; Mercenier, A.; Rastall, R.; Roberfroid, M.; Rowland, I.; Cherbut, C.; Klaenhammer, T.R. New scientific paradigms for probiotics and prebiotics. J. Clin. Gastroenterol. 2003, 37, 105–118. [Google Scholar] [PubMed] [Green Version]
- Martins, F.S.; Miranda, I.C.; Rosa, C.A.; Nicoli, J.R.; Neves, M.J. Effect of the trehalose levels on the screening of yeast as probiotic by in vivo and in vitro assays. Braz. J. Microbiol. 2008, 39, 50–55. [Google Scholar]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Genes and molecules of lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar]
- Boonaert, C.J.; Rouxhet, P.G. Surface of lactic acid bacteria: Relationships between chemical composition and physicochemical properties. Appl. Environ. Microbiol. 2000, 66, 2548–2554. [Google Scholar] [PubMed] [Green Version]
- Duary, R.K.; Rajput, Y.S.; Batish, V.K.; Grover, S. Assessing the adhesion of putative indigenous probiotic lactobacilli to human colonic epithelial cells. Indian J. Med. Res. 2011, 134, 664. [Google Scholar]
- Rivera, E.Y.; Gallardo, N.Y. Non-dairy probiotic products. Food Microbiol. 2010, 27, 1–11. [Google Scholar]
- Almeida, M.H.B.; Zoellner, S.S.; Cruz, A.G.; Moura, M.R.L.; Carvalho, L.M.J.; Santana, A.S. Potentially probiotic acid yoghurt. Int. J. Dairy Technol. 2008, 61, 178–182. [Google Scholar]
- Lavermicocca, P. Highlights on new food research. Dig. Liver Dis. 2006, 38, 295–299. [Google Scholar]
- Boyle, R.J.; Tang, M.L. The role of probiotics in the management of allergic disease. Clin. Exp. Allergy 2006, 36, 568–576. [Google Scholar]
- Senok, A.C.; Ismaeel, A.Y.; Botta, G.A. Probiotics: Facts and myths. Clin. Microbiol. Infect. 2005, 11, 958–966. [Google Scholar]
- Shah, N.P. Functional cultures and health benifits. Int. Dairy J. 2007, 17, 1262–1277. [Google Scholar]
- Angmo, K.; Kumari, A.; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LWT-Food Sci. Technol. 2016, 66, 428–435. [Google Scholar]
- Rajoka, M.S.R.; Mehwish, H.M.; Siddiq, M.; Haobin, Z.; Zhu, J.; Yan, L.; Shao, D.; Xu, X.; Shi, J. Identification, characterization, and probiotic potential of Lactobacillus rhamnosus isolated from human milk. LWT-Food Sci. Technol. 2017, 84, 271–280. [Google Scholar]
- Mulaw, G.; Sisay, T.T.; Muleta, D.; Tesfaye, A. In vitro evaluation of probiotic properties of lactic acid bacteria isolated from some traditionally fermented Ethiopian food products. Int. J. Microbiol. 2019, 2019, 7179514. [Google Scholar] [PubMed] [Green Version]
- Finlay, W.J.J.; Logan, N.A.; Sutherland, A.D. Semiautomated metabolic staining assay for Bacillus cereus emetic toxin. Appl. Environ. Microbiol. 1999, 65, 1811–1812. [Google Scholar]
- Umthong, S.; Phuwapraisirisan, P.; Puthong, S.; Chanchao, C. In vitro antiproliferative activity of partially purified Trigona laeviceps propolis from Thailand on human cancer cell lines. BMC Complement Altern. Med. 2011, 11, 37. [Google Scholar]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Uroic, K.; Nikolic, M.; Kos, B.; Pavunc, A.L.; Beganovic, J.; Lukic, J.; Jovic, B.; Filipic, B.; Miljkovic, M.; Golic, N.; et al. Probiotic properties of lactic acid bacteria isolated from Croatian fresh soft cheese and Serbian white pickled cheese 2014. Food Technol. Biotechnol. 2014, 52, 232–241. [Google Scholar]
- Carunchiawhetstine, M.E.; Karagul, Y.Y.; Avsar, Y.K.; Drake, M.A. Identification and quantification of character aroma components in fresh chevre-style goat cheese. J. Food Sci. 2003, 68, 2441–2447. [Google Scholar]
- Madalozzo, E.S.; Sauer, E.; Nagata, N. Determination of fat, protein and moisture in ricotta cheese by near infrared spectroscopy and multivariate calibration. J. Food Sci. Technol. 2015, 52, 1649–1655. [Google Scholar]
- Mukdsi, M.C.A.; Haro, C.; González, S.N.; Medina, R.B. Functional goat milk cheese with feruloyl esterase activity. J. Funct. Foods. 2013, 5, 801–809. [Google Scholar]
- Ouwehand, A.C.; Salminen, S.J. The health effects of cultured milk products with viable and non-viable bacteria. Int. Dairy J. 1998, 8, 749–758. [Google Scholar]
- Pyar, H.; Peh, K.K. Characterization and identification of Lactobacillus acidophilus using biolog rapid identification system. Int. J. Pharm. Pharm. Sci. 2014, 6, 189–193. [Google Scholar]
- Haghshenas, B.; Yousef, N.; Ali, A.; Norhafizah, A.; Dayang, R.; Rozita, R.; Abolfazl, B.; Ahmad, Y.K. Isolation and characterization of probiotics from dairies. Iran. J. Microbiol. 2017, 9, 234–243. [Google Scholar] [PubMed]
- Husebye, E. The pathogenesis of gastrointestinal bacterial overgrowth. Chemotherapy 2005, 51, 1–22. [Google Scholar]
- Yokota, A.; Veenstra, M.; Kurdi, P.; van Veen, H.W.; Konings, W.N. Cholate resistance in Lactococcus lactis is mediated by an ATP-dependent multispecific organic anion transporter. J. Bacteriol. 2000, 182, 5196–5201. [Google Scholar] [PubMed] [Green Version]
- Saadatzadeh, A.; Fazeli, M.R.; Jamalifar, H.; Dinarvand, R. Probiotic properties of lyophilized cell free extract of Lactobacillus casei. Jundishapur J. Nat. Pharm. Prod. 2013, 8, 131–137. [Google Scholar] [PubMed] [Green Version]
- Lee, Y.K.; Puong, K.Y.; Ouwehand, A.C.; Salminen, S. Displacement of bacterial pathogens from mucus and Caco-2 cell surface by lactobacilli. J. Med. Microbiol. 2003, 52, 925–930. [Google Scholar]
- Muriana, P.M.; Klaenhammer, T.R. Purification and partial characterization of lactacin F, a bacteriocin produced by Lactobacillus acidophilus 11088. Appl. Environ. Microbiol. 1991, 57, 114–121. [Google Scholar]
- Van Reenen, C.A.; Dicks, L.M.T.; Chikindas, M.L. Isolation, purification and partial characterization of plantaricin 423, a bacteriocin produced by Lactobacillus plantarum. J. Appl. Microbiol. 1998, 84, 1131–1137. [Google Scholar] [PubMed] [Green Version]
- De Kwaadsteniet, M.; Todorov, S.D.; Knoetze, H.; Dicks, L.M.T. Characterization of a 3944 Da bacteriocin, produced by Enterococcus Mundtii ST15, with activity against Gram-positive and Gram-negative bacteria. Int. J. Food Microbiol. 2005, 105, 433–444. [Google Scholar] [PubMed]
- Zendo, T.; Masanori, F.; Kyoko, U.; Tomoko, H.; Jiro, N.; Kenji, S. Identification of the lantibiotic nisin Q, a new natural nisin variant produced by Lactococcus Lactis 61-14 isolated from a river in Japan. Biosci. Biotechnol. Biochem. 2003, 67, 1616–1619. [Google Scholar] [PubMed] [Green Version]
- Caggia, C.; De Angelis, M.; Pitino, I.; Pino, A.; Randazzo, C.L. Probiotic features of Lactobacillus strains isolated from Ragusano and Pecorino Siciliano cheeses. Food Microbiol. 2015, 50, 109–117. [Google Scholar] [PubMed]
- Paolillo, R.; Carratelli, C.R.; Sorrentino, S.; Mazzola, N.; Rizzo, A. Immunomodulatory effects of Lactobacillus plantarum on human colon cancer cells. Int. Immunopharmacol. 2009, 9, 1265–1271. [Google Scholar]
- International Organization for Standardization. Biological Evaluation of Medical Devices; British Standards Institution: London, UK, 2009; pp. 1–42. [Google Scholar]
- Bourne, M. Food Texture and Viscosity: Concept and Measurement; Academic Press: London, UK, 2002; pp. 107–187. [Google Scholar]
- Vinderola, C.G.; Reinheimer, J.A. Enumeration of Lactobacillus casei in the presence of L. acidophilus, bifidobacteria and lactic starter bacteria in fermented dairy products. Int. Dairy J. 2000, 10, 271–275. [Google Scholar]
- Ross, R.P.; Fitzgerald, G.; Collins, K.; Stanton, C. Cheese delivering biocultures—Probiotic cheese. Aust. J. Dairy Technol. 2002, 57, 71. [Google Scholar]
- Addis, E.; Fleet, G.H.; Cox, J.M.; Kolak, D.; Leung, T. The growth, properties and interactions of yeasts and bacteria associated with the maturation of Camembert and blue-veined Cheeses. Int. J. Food Microbiol. 2001, 69, 25–36. [Google Scholar] [PubMed]
- Romano, P.; Capece, A.; Jespersen, L. Taxonomic and ecological diversity of food and beverage yeasts. In Yeasts Food Beverages; Springer: Heidelberg, Germany, 2006; pp. 13–53. [Google Scholar]
- Fleet, G.H. Yeast spoilage of foods and beverages. Yeasts 2011, 1, 53–63. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Category | Sample | No. of Sample | Source |
|---|---|---|---|
| Milk and dairy | Milk kefir | 1 | Russia |
| Raw goat’s milk | 5 | Chiang Mai, Thailand | |
| Fermented food | Pla-som (fermented fish) | 3 | Pa-Yao, Thailand |
| Tua-nao (alkaline fermented soybean) | 4 | Chiang Mai, Thailand | |
| Pickled garlic | 2 | Chiang Mai, Thailand | |
| Pickled cabbage | 3 | Chiang Mai, Thailand | |
| Miang (fermented tea leaf product) | 4 | Chiang Mai, Thailand | |
| kimchi | 3 | Chiang Mai, Thailand |
| Source | No. of Isolate | Gram Stain Reaction and Cell Morphology | Catalase Production | |
|---|---|---|---|---|
| Fermented foods | Kimchi | 29 | Gram-positive, rod-shaped | negative |
| Pickled cabbage | 29 | Gram-positive, rod-shaped | negative | |
| 13 | Gram-positive, short rod-shaped | negative | ||
| Miang | 13 | Gram-positive, rod-shaped | negative | |
| 17 | Gram-positive, short rod-shaped | negative | ||
| 7 | Gram-positive, spherical-shaped | negative | ||
| Tua-nao | 8 | Gram-positive, rod-shaped | negative | |
| 12 | Gram-positive, short rod-shaped | negative | ||
| 12 | Gram-positive, spherical-shaped | negative | ||
| Pickled garlic | 5 | Gram-positive, rod shaped | negative | |
| 8 | Gram-positive, spherical-shaped | negative | ||
| Pla-som | 17 | Gram-positive, rod-shaped | negative | |
| 25 | Gram-positive, short rod-shaped | negative | ||
| Milk | Raw goat’s milk | 38 | Gram-positive, rod-shaped | negative |
| Dairy product | Milk kefir | 8 | Gram-positive, rod-shaped | negative |
| Total | 241 | |||
| Isolate | Source | Closest Species/Strain | % Identity | Accession Number of Closet Relative | Accession Number * |
|---|---|---|---|---|---|
| AD22 | Kimchi | Lactobacillus plantarum strain JCM 1149 | 100.00 | NR_115605.1 | OM807265 |
| AD62 | Tua-nao | Lactobacillus fermentum strain CIP 102980 | 99.80 | NR_104927.1 | OM807266 |
| AD72 | Milk kefir | Lactobacillus plantarum strain JCM 1149 | 100.00 | NR_115605.1 | OM807267 |
| AD73 | Milk kefir | Lactobacillus plantarum strain JCM 1149 | 99.93 | NR_115605.1 | OM807268 |
| AD85 | Miang | Lactobacillus fermentum strain CIP 102980 | 99.80 | NR_104927.1 | OM807269 |
| AD118 | Raw goat’s milk | Lactobacillus fermentum strain CIP 102980 | 99.93 | NR_104927.1 | OM807270 |
| Test | AD22 | AD62 | AD72 | AD73 | AD85 | AD118 |
|---|---|---|---|---|---|---|
| Control | - | - | - | - | - | - |
| Glycerol | - | + | - | - | - | - |
| Erythritol | - | - | - | - | - | - |
| D-Arabinose | - | - | - | - | - | - |
| L-Arabinose | + | + | + | + | + | + |
| D-Ribose | + | + | + | + | + | + |
| D-Xylose | - | - | - | - | - | - |
| L-Xylose | - | - | - | - | - | - |
| D-Adonitol | - | - | - | - | - | - |
| Methyl β-D-glucopyranoside | - | - | - | - | - | - |
| D-Galactose | + | + | + | + | + | - |
| D-Glucose | + | + | + | + | + | + |
| D-Fructose | + | + | + | + | + | + |
| D-Mannose | + | + | + | + | + | + |
| L-Sorbose | - | - | - | - | - | - |
| L-Rhamnose | - | - | - | - | - | - |
| Dulcitol | - | - | - | - | - | - |
| Inositol | - | - | - | - | - | - |
| D-Mannitol | - | - | + | + | - | - |
| D-Sorbitol | - | - | - | - | - | - |
| Methyl α-D-mannopyranoside | - | - | + | + | - | - |
| Methyl α-D-glucoside | - | - | - | - | - | - |
| N-Acetyl glucosamine | - | - | + | + | - | - |
| Amygdalin | - | - | + | + | - | - |
| Arbutin | - | - | + | + | - | - |
| Esculin ferric citrate salicin | - | - | + | + | - | + |
| D-Cellobiose | - | - | + | + | - | - |
| D-Maltose | - | - | + | + | - | - |
| Arbutin | + | + | + | + | + | + |
| D-Melibiose | + | + | + | + | + | - |
| D-Melibiose | + | + | + | + | + | - |
| D-Saccharose | + | + | + | + | + | + |
| D-Trehalose | - | - | + | + | - | - |
| Inulin | - | - | - | - | - | - |
| D-Melezitose | - | - | + | + | - | - |
| D-Raffinose | + | + | + | + | + | + |
| Amidon | - | - | - | - | - | - |
| Glycogen | - | - | - | - | - | - |
| Xylitol | - | - | - | - | - | - |
| Gentobiose | - | - | + | + | - | - |
| D-Turanose | - | - | + | + | - | - |
| D-Lyxose | - | - | - | - | - | - |
| Tagatose | - | - | - | - | - | - |
| D-Focose | - | - | - | - | - | - |
| D-Lucose | - | - | - | - | - | - |
| L-Fucose | - | - | - | - | - | - |
| D-Arabitol | - | - | - | - | - | - |
| Potassium gluconate | - | - | + | + | - | + |
| Potassium 2-keto gluconate | - | - | - | - | - | - |
| Potassium 5-keto gluconate | - | - | - | - | - | - |
| Isolate | Inhibition Zone (mm) | ||||
|---|---|---|---|---|---|
| B. cereus DSM4384 | B. cereus F4810/72 | Staphylococcus aureus | Salmonella enterica subsp. enterica | E. coli O157:H7 | |
| AD22 | 12.3 ± 0.07 | 12.0 ± 0.20 * | 12.5 ± 0.07 | 12.9 ± 0.03 * | 11.9 ± 0.05 |
| AD62 | 12.2 ± 0.06 | 15.0 ± 0.10 * | 12.2 ± 0.05 * | 12.1 ± 0.03 | 12.3 ± 0.10 * |
| AD72 | 14.3 ± 0.09 * | 14.0 ± 0.15 * | 12.7 ± 0.02 | 11.0 ± 0.05 | 13.5 ± 0.05 * |
| AD73 | 13.0 ± 0.05 * | 11.3 ± 0.10 * | 13.5 ± 0.03 * | 11.9 ± 0.04 | 12.0 ± 0.05 |
| AD85 | 14.6 ± 0.07 * | 14.6 ± 0.10 * | 11.7 ± 0.02 * | 12.4 ± 0.06 * | 13.1 ± 0.06 * |
| AD118 | 11.8 ± 0.09 * | 11.6 ± 0.04 * | 12.5 ± 0.02 | 11.2 ± 0.05 | 12.0 ± 0.05 |
| Penicillin | 21.0 ± 0.05 | 20.0 ± 0.03 | 22.0 ± 0.05 | ND | ND |
| Polymyxin B | ND | ND | ND | 16.0 ± 0.07 | 15.0 ± 0.07 |
| Isolate | Inhibition Zone (mm) | ||||
|---|---|---|---|---|---|
| B. cereus DSM4384 | B. cereus F4810/72 | Staphylococcus aureus | Salmonella enterica subsp. enterica | E. coli O157:H7 | |
| AD22 | 11.0 ± 0.02 | 9.0 ± 0.05 * | 7.0 ± 0.01 | 12.0 ± 0.03 | 9.0 ± 0.10 * |
| AD62 | 14.0 ± 0.14 * | 13.5 ± 0.14 * | 6.0 ± 0.02 * | 10.0 ± 0.14 * | 9.5 ± 0.07 * |
| AD72 | 13.0 ± 0.14 * | 10.5 ± 0.07 * | 8.5 ± 0.07 | 12.0 ± 0.14 | 11.0 ± 0.14 * |
| AD73 | 11.0 ± 0.05 | 12.0 ± 0.12 * | 9.5 ± 0.07 * | 13.0 ± 0.10 * | 8.0 ± 0.01 |
| AD85 | 12.0 ± 0.05 * | 11.0 ± 0.15 * | 8.5 ± 0.07 | 8.0 ± 0.14 * | 7.0 ± 0.12 * |
| AD118 | 8.0 ± 0.07 * | 7.5 ± 0.07 * | 7.0 ± 0.06 | 9.5 ± 0.07 * | 8.0 ± 0.05 |
| Penicillin | 21.0 ± 0.05 | 20.0 ± 0.03 | 22.0 ± 0.05 | ND | ND |
| Polymyxin B | ND | ND | ND | 16.0 ± 0.07 | 15.0 ± 0.07 |
| Characterisation | Chèvre Cheese Made with Starter Culture * | Chèvre Cheese Made with Starter Culture * and Probiotic |
|---|---|---|
| Raw goat’s milk | 12 kg | 12 kg |
| Whey volume | 7.19 L | 7.19 L |
| Weight of cheese | 2.185 kg | 2.010 kg |
| Percent yield of cheese (by weight of goat’s milk) | 18.20% | 16.75% |
| Moisture content | 64.53 ± 0.62% ** | 61.33 ± 0.68% ** |
| Colour *** | white | white |
| Texture of cheese *** | firm, homogenous and smooth texture | firm but not completely homogenous texture (lumps found in curd) |
| Day | pH | Total Viable Count (log CFU/g) | Lactic Acid Bacteria * (log CFU/g) | Yeast and Mould Count ** (log CFU/g) | Detection of Deviated Smell from Day 0 *** |
|---|---|---|---|---|---|
| 0 | 4.42 | 9.84 ± 0.02 | 10.03 ± 0.02 | 2.31 ± 0.01 | NA |
| 2 | 4.38 | 9.81 ± 0.20 | 9.40 ± 0.03 | 2.27 ± 0.25 | ND |
| 4 | 4.36 | 8.82 ± 0.02 | 8.47 ± 0.03 | 3.17 ± 0.02 | ND |
| 6 | 4.35 | 8.38 ± 0.02 | 7.92 ± 0.05 | 4.53 ± 0.02 | ND |
| 8 | 4.33 | 8.37 ± 0.04 | 7.01 ± 0.03 | 5.23 ± 0.02 | ND |
| 10 | 4.30 | 7.99 ± 0.11 | 6.92 ± 0.01 | 6.02 ± 0.01 | D (alcoholic smell) |
| 12 | 4.25 | 8.52 ± 0.05 | 6.86 ± 0.01 | 6.35 ± 0.05 | D (alcoholic smell) |
| 14 | 4.28 | 6.39 ± 0.04 | 6.61 ± 0.01 | 6.61 ± 0.01 | D (alcoholic smell) |
| Day | pH | Total Viable Count (log CFU/g) | Number of Lactic Acid Bacteria (log CFU/g) | Yeast and Mould Count ** (log CFU/g) | Detection of Deviated Smell from Day 0 *** | ||
|---|---|---|---|---|---|---|---|
| Total Lactic Acid Bacteria | LAB Starters | L. plantarum AD73 * | |||||
| 0 | 4.36 | 9.81 ± 0.02 | 10.49 ± 0.01 | 10.49 ± 0.01 | 8.63 ± 0.04 | 2.16 ± 0.28 | NA |
| 2 | 4.36 | 9.76 ± 0.04 | 9.28 ± 0.04 | 9.22 ± 0.04 | 8.41 ± 0.08 | 2.57 ± 0.23 | ND |
| 4 | 4.35 | 8.65 ± 0.03 | 9.19 ± 0.05 | 8.61 ± 0.03 | 7.83 ± 0.13 | 3.12 ± 0.04 | ND |
| 6 | 4.35 | 8.59 ± 0.04 | 8.33 ± 0.01 | 8.18 ± 0.01 | 7.77 ± 0.05 | 4.00 ± 0.01 | ND |
| 8 | 4.36 | 8.39 ± 0.08 | 8.00 ± 0.03 | 7.81 ± 0.03 | 7.56 ± 0.05 | 5.64 ± 0.02 | ND |
| 10 | 4.37 | 8.15 ± 0.09 | 7.91 ± 0.02 | 7.70 ± 0.02 | 7.49 ± 0.08 | 5.98 ± 0.01 | D (alcoholic smell) |
| 12 | 4.37 | 8.33 ± 0.03 | 7.01 ± 0.03 | 6.81 ± 0.01 | 6.58 ± 0.09 | 6.65 ± 0.02 | D (alcoholic smell) |
| 14 | 4.35 | 6.76 ± 0.03 | 6.82 ± 0.01 | 6.56 ± 0.04 | 6.46 ± 0.02 | 6.79 ± 0.03 | D (alcoholic smell) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngamsomchat, A.; Kaewkod, T.; Konkit, M.; Tragoolpua, Y.; Bovonsombut, S.; Chitov, T. Characterisation of Lactobacillus plantarum of Dairy-Product Origin for Probiotic Chèvre Cheese Production. Foods 2022, 11, 934. https://doi.org/10.3390/foods11070934
Ngamsomchat A, Kaewkod T, Konkit M, Tragoolpua Y, Bovonsombut S, Chitov T. Characterisation of Lactobacillus plantarum of Dairy-Product Origin for Probiotic Chèvre Cheese Production. Foods. 2022; 11(7):934. https://doi.org/10.3390/foods11070934
Chicago/Turabian StyleNgamsomchat, Achirawit, Thida Kaewkod, Maytiya Konkit, Yingmanee Tragoolpua, Sakunnee Bovonsombut, and Thararat Chitov. 2022. "Characterisation of Lactobacillus plantarum of Dairy-Product Origin for Probiotic Chèvre Cheese Production" Foods 11, no. 7: 934. https://doi.org/10.3390/foods11070934