
Ozone-Microbubble-Washing with Domestic Equipment: Effects on the Microstructure, and Lipid and Protein Oxidation of Muscle Foods

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Treatments
2.3. Assessment of Muscle Microstructure
2.4. Quantification of Thiobarbituric Acid Reactive Substances (TBARS) Values
2.5. Quantification of Glyoxal (GO) and Methylglyoxal (MGO)
2.5.1. Sample Pretreatment
2.5.2. UPLC-MS/MS Analysis
2.6. Quantification of Free Amino Acids
2.6.1. Sample Pretreatment
2.6.2. HPLC Analysis
2.7. Identification of Oxidation Sites on Proteins
2.7.1. Protein Extraction
2.7.2. Protein Digestion
2.7.3. High-Resolution Mass Spectrometry (HR-MS) Analysis
2.8. Statistical Analysis
3. Results and Discussion
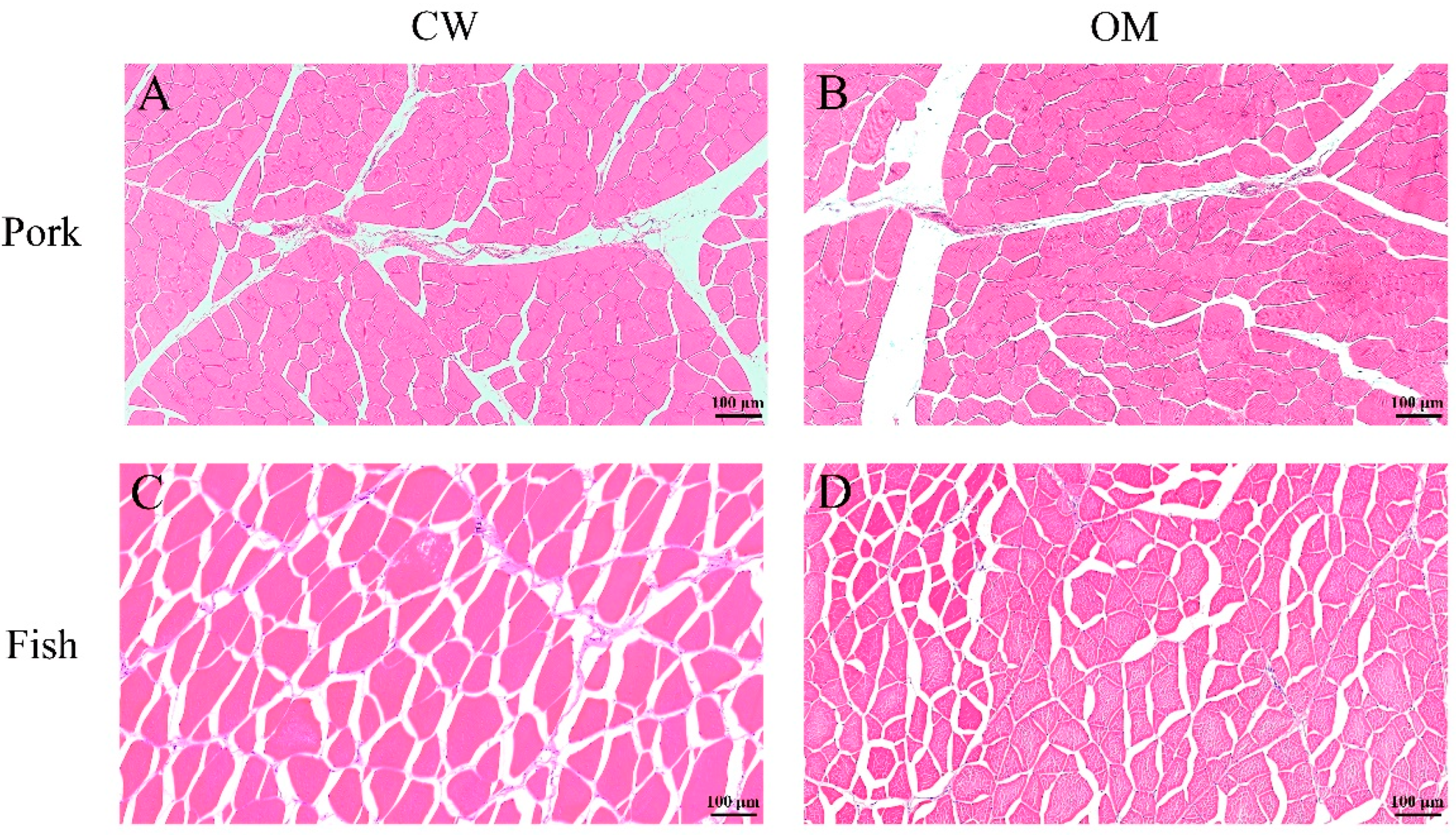
3.1. Effects of OM Treatment on Microstructure of Muscle Tissue
3.2. Effects of OM Treatment on Lipid Oxidation
3.2.1. TBARS Values
3.2.2. GO and MGO
3.3. Effects of OM Treatment on Protein Oxidation
3.3.1. Free Amino Acids
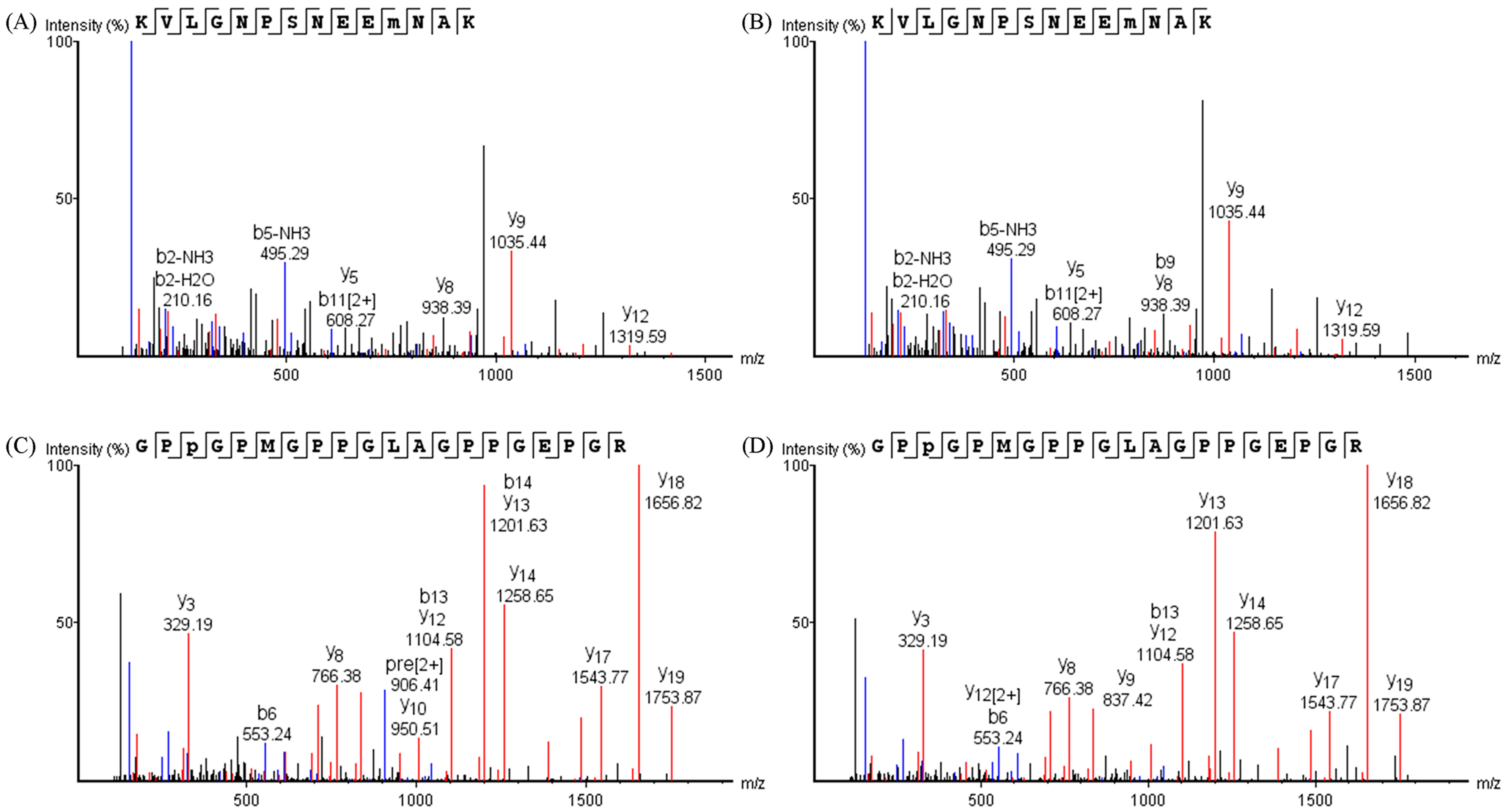
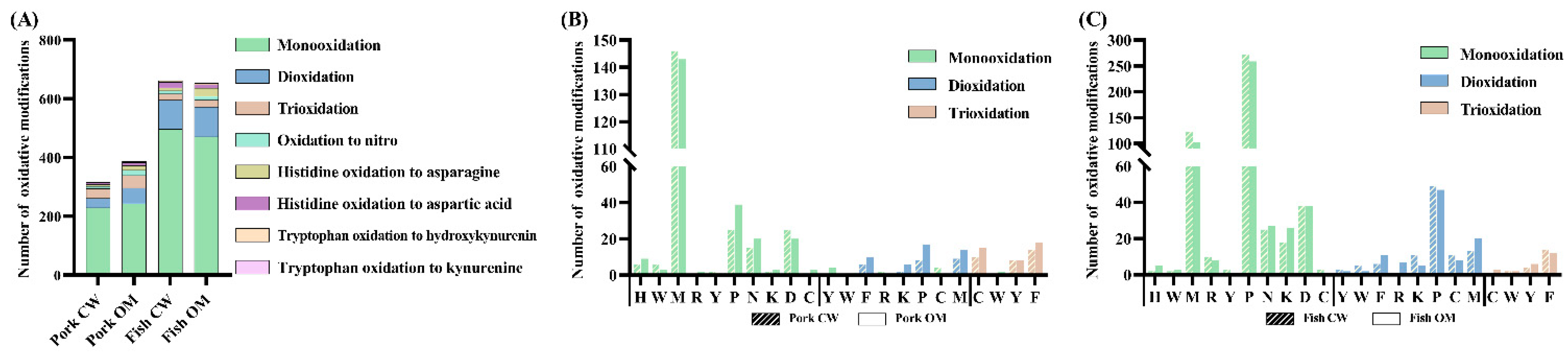
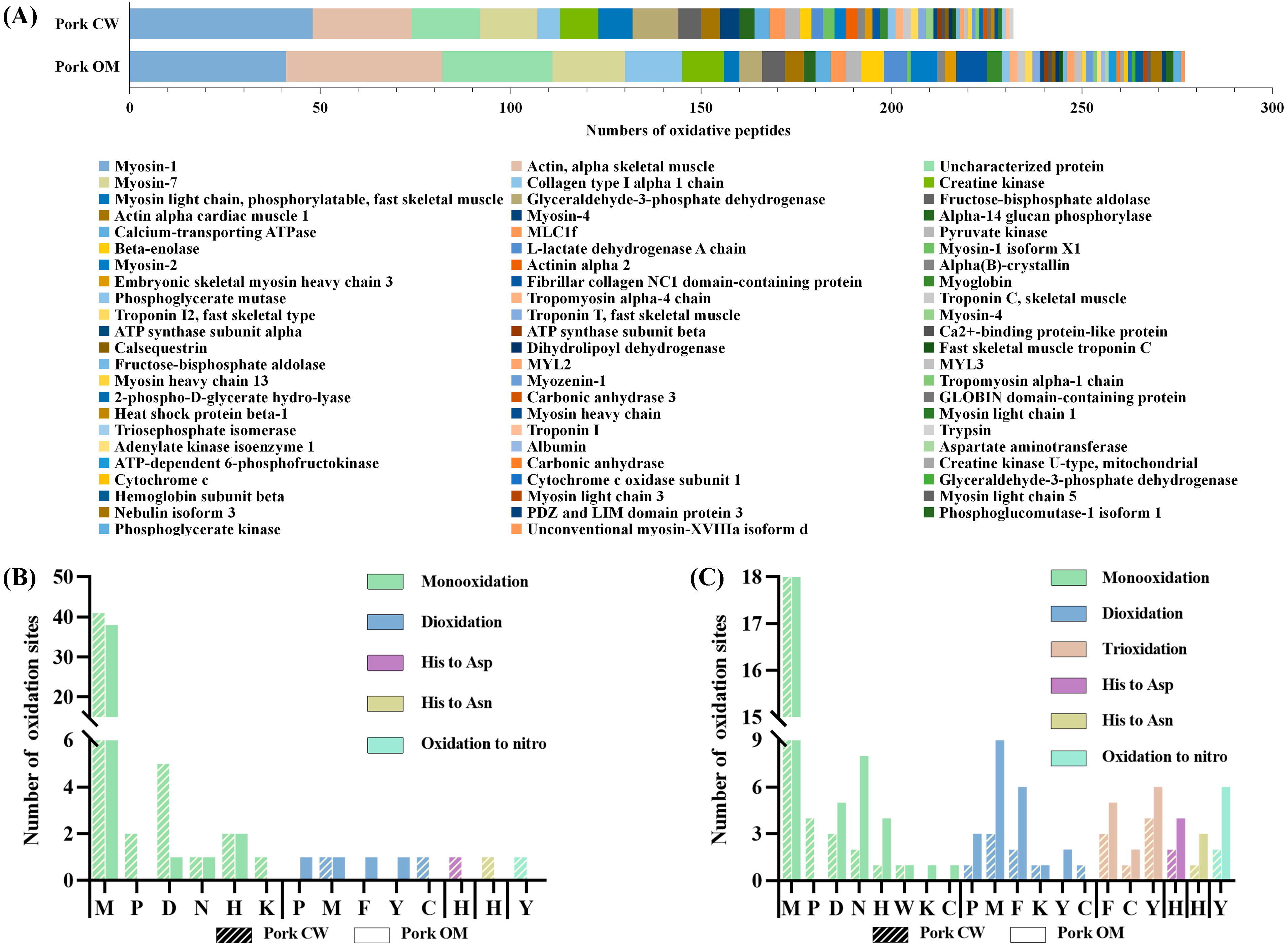
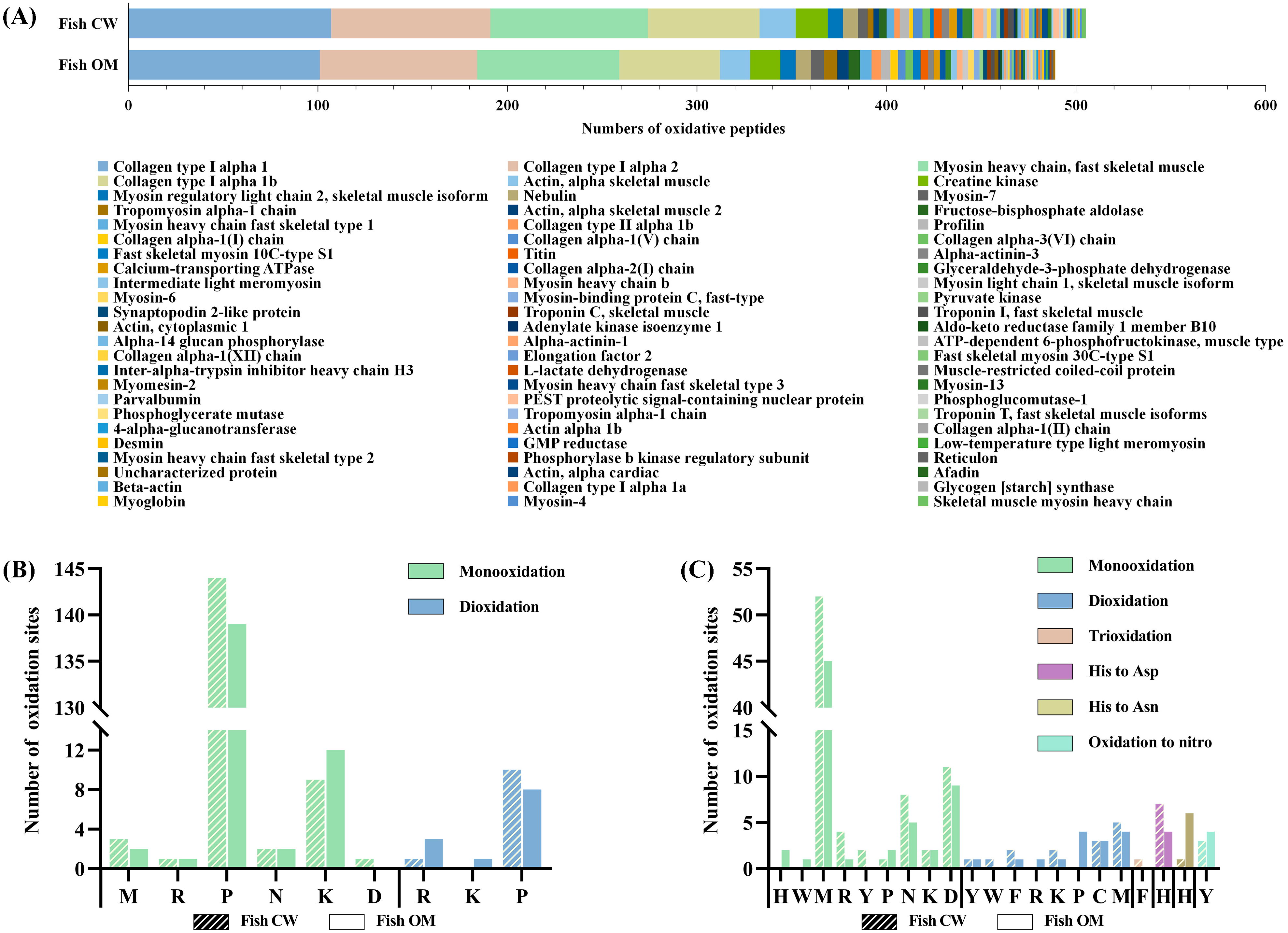
3.3.2. Oxidation Sites on Proteins
- Types of oxidative modifications
- Types and properties of oxidatively modified proteins of pork samples
- Types and properties of oxidatively modified proteins of fish samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elvis, A.M.; Ekta, J.S. Ozone therapy: A clinical review. J. Nat. Sci. Biol. Med. 2011, 2, 66–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, Z.; Barbara, R. Inactivation of microbes by ozone in the food industry: A review. Afr. J. Food Sci. 2021, 15, 113–120. [Google Scholar] [CrossRef]
- Cattel, F.; Giordano, S.; Bertiond, C.; Lupia, T.; Corcione, S.; Scaldaferri, M.; Angelone, L.; De Rosa, F.G. Ozone therapy in COVID-19: A narrative review. Virus Res. 2021, 291, 198207. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Zhang, F.; Hu, Y.; Feng, C.; Wu, H. Ozonation in water treatment: The generation, basic properties of ozone and its practical application. Rev. Chem. Eng. 2017, 33, 49–89. [Google Scholar] [CrossRef]
- Khanashyam, A.C.; Shanker, M.A.; Kothakota, A.; Mahanti, N.K.; Pandiselvam, R. Ozone Applications in Milk and Meat Industry. Ozone Sci. Eng. 2022, 44, 50–65. [Google Scholar] [CrossRef]
- Horvitz, S.; Arancibia, M.; Arroqui, C.; Chonata, E.; Vírseda, P. Effects of Gaseous Ozone on Microbiological Quality of Andean Blackberries (Rubus glaucus Benth). Foods 2021, 10, 2039. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, X.; Li, L.; Hao, S.; Wei, Y.; Cen, J.; Lin, H. Chemical, Microbiological, Color and Textural Changes in Nile Tilapia (Oreochromis niloticus) Fillets Sterilized by Ozonated Water Pretreatment During Frozen Storage. J. Food Processing Preserv. 2016, 41, e12746. [Google Scholar] [CrossRef]
- Mohammadi, H.; Mazloomi, S.M.; Eskandari, M.H.; Aminlari, M.; Niakousari, M. The effect of ozone on aflatoxin M1, oxidative stability, carotenoid content and the microbial count of milk. Ozone Sci. Eng. 2017, 39, 447–453. [Google Scholar] [CrossRef]
- Schrader, K.K.; Davidson, J.W.; Rimando, A.M.; Summerfelt, S.T. Evaluation of ozonation on levels of the off-flavor compounds geosmin and 2-methylisoborneol in water and rainbow trout Oncorhynchus mykiss from recirculating aquaculture systems. Aquac. Eng. 2010, 43, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Giacosa, S.; Río Segade, S.; Gai, G.; Dogliani, E.; Gerbi, V.; Cocolin, L.; et al. Minimizing the environmental impact of cleaning in winemaking industry by using ozone for cleaning-in-place (CIP) of wine bottling machine. J. Clean. Prod. 2019, 233, 582–589. [Google Scholar] [CrossRef]
- Liu, C.; Chen, C.; Jiang, A.; Zhang, Y.; Zhao, Q.; Hu, W. Effects of aqueous ozone treatment on microbial growth, quality, and pesticide residue of fresh-cut cabbage. Food Sci. Nutr. 2021, 9, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Okpala, C. Physicochemical changes in minimal ozone-treated fresh shrimp stored on ice tandem with microbiological efficacy of treatment. Niger. Food J. 2015, 33, 74–87. [Google Scholar]
- Okpala, C.O.R. Investigation of quality attributes of ice-stored Pacific white shrimp (Litopenaeus vannamei) as affected by sequential minimal ozone treatment. LWT Food Sci. Technol. 2014, 57, 538–547. [Google Scholar] [CrossRef]
- Li, C.; Xie, Y.; Guo, Y.; Cheng, Y.; Yu, H.; Qian, H.; Yao, W. Effects of ozone-microbubble treatment on the removal of residual pesticides and the adsorption mechanism of pesticides onto the apple matrix. Food Control 2021, 120, 107548. [Google Scholar] [CrossRef]
- Lee, U.; Joo, S.; Klopfenstein, N.B.; Kim, M.-S. Efficacy of washing treatments in the reduction of post-harvest decay of chestnuts (Castanea crenata ‘Tsukuba’) during storage. Can. J. Plant Sci. 2016, 96, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Ikeura, H.; Hamasaki, S.; Tamaki, M. Effects of ozone microbubble treatment on removal of residual pesticides and quality of persimmon leaves. Food Chem. 2013, 138, 366–371. [Google Scholar] [CrossRef]
- Okpala, C.O.R. Changes in some proximate, colour and textural characteristics of ozone-processed shrimp: Combined effects of increasing ozone discharge and iced storage. Iran. J. Fish. Sci. 2017, 16, 625–638. [Google Scholar]
- Okpala, C.O.R.; Bono, G.; Geraci, M.L.; Sardo, G.; Vitale, S.; Schaschke, C.J. Lipid oxidation kinetics of ozone-processed shrimp during iced storage using peroxide value measurements. Food Biosci. 2016, 16, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Yun, Y.; Mi, J.; Luo, J.; Jin, M.; Nie, G.; Zhou, Q. Effects of faba bean on growth performance and fillet texture of Yellow River carp, Cyprinus carpio haematopterus. Aquac. Rep. 2020, 17, 100379. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, F.; Zhang, T.; Cui, M.; Peng, B.; Zhang, Y.; Wang, S. Simultaneous Determination of Seven α-Dicarbonyl Compounds in Milk and Milk Products Based on an LC–MS/MS Method with Matrix-Matched Calibration. Food Anal. Methods 2022. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, H.; McClements, D.J.; Nie, S.; Shen, M.; Li, C.; Huang, Y.; Chen, J.; Zeng, M.; Xie, M. Effect of fatty acids and triglycerides on the formation of lysine-derived advanced glycation end-products in model systems exposed to frying temperature. RSC Adv. 2019, 9, 15162–15170. [Google Scholar] [CrossRef] [Green Version]
- Goh, K.M.; Lai, O.M.; Abas, F.; Tan, C.P. Effects of sonication on the extraction of free-amino acids from moromi and application to the laboratory scale rapid fermentation of soy sauce. Food Chem. 2017, 215, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Ngapo, T.M.; Vachon, L. Biogenic amine concentrations and evolution in “chilled” Canadian pork for the Japanese market. Food Chem. 2017, 233, 500–506. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Dann, G.P.; Shi, T.; Wang, L.; Gao, X.; Su, D.; Nicora, C.D.; Shukla, A.K.; Moore, R.J.; Liu, T.; et al. Simple sodium dodecyl sulfate-assisted sample preparation method for LC-MS-based proteomics applications. Anal. Chem. 2012, 84, 2862–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Mao, J.-l.; Yao, H.; Aubourg, S.P. Label-free based proteomics analysis of protein changes in frozen whiteleg shrimp (Litopenaeus vannamei) pre-soaked with sodium trimetaphosphate. Food Res. Int. 2020, 137, 109455. [Google Scholar] [CrossRef]
- Yurina, L.; Vasilyeva, A.; Indeykina, M.; Bugrova, A.; Biryukova, M.; Kononikhin, A.; Nikolaev, E.; Rosenfeld, M. Ozone-induced damage of fibrinogen molecules: Identification of oxidation sites by high-resolution mass spectrometry. Free Radic. Res. 2019, 53, 430–455. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Sun, W.; Xiong, G.; Shi, L.; Jiao, C.; Wu, W.; Li, X.; Qiao, Y.; Liao, L.; Ding, A.; et al. Effects of HVEF treatment on microbial communities and physicochemical properties of catfish fillets during chilled storage. LWT 2020, 131, 109667. [Google Scholar] [CrossRef]
- Chen, L.; Bao, P.; Wang, Y.; Hu, Y.; Fang, H.; Yang, H.; Zhang, B.; He, B.; Zhou, C. Improving quality attributes of refrigerated prepared pork chops by injecting l-arginine and l-lysine solution. LWT 2022, 153, 112423. [Google Scholar] [CrossRef]
- Nakyinsige, K.; Sazili, A.Q.; Aghwan, Z.A.; Zulkifli, I.; Goh, Y.M.; Abu Bakar, F.; Sarah, S.A. Development of microbial spoilage and lipid and protein oxidation in rabbit meat. Meat Sci. 2015, 108, 125–131. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Chaijan, M.; Panpipat, W. Pre-neutralized crude palm oil as natural colorant and bioactive ingredient in fish sausage prepared from tilapia (Oreochromis niloticus). LWT 2021, 135, 110289. [Google Scholar] [CrossRef]
- Giménez, B.; Graiver, N.; Giannuzzi, L.; Zaritzky, N. Treatment of beef with gaseous ozone: Physicochemical aspects and antimicrobial effects on heterotrophic microflora and listeria monocytogenes. Food Control 2021, 121, 107602. [Google Scholar] [CrossRef]
- Avery, N.C.; Bailey, A.J. Enzymic and non-enzymic cross-linking mechanisms in relation to turnover of collagen: Relevance to aging and exercise. Scand. J. Med. Sci. Sports 2005, 15, 231–240. [Google Scholar] [CrossRef] [PubMed]
- De Mendonça Silva, A.M.; Gonçalves, A.A. Effect of aqueous ozone on microbial and physicochemical quality of Nile tilapia processing. J. Food Processing Preserv. 2017, 41, e13298. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, Z.; He, Z.; Wang, Z.; Qin, F.; Zeng, M.; Chen, J. Effect of Freeze-Thaw Cycles on the Oxidation of Protein and Fat and Its Relationship with the Formation of Heterocyclic Aromatic Amines and Advanced Glycation End Products in Raw Meat. Molecules 2021, 26, 1264. [Google Scholar] [CrossRef]
- Zhao, G.; Bai, X.; Tian, W.; Ru, A.; Li, J.; Wang, H.; Wang, K.; Xiao, K.; Zhu, C.; Li, H. The effect of shower time, electrolyte treatment, and electrical stimulation on meat quality of cattle longissimus thoracis muscle in cold weather. Meat Sci. 2022, 184, 108664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, X.; Chen, W.; Zhou, P. Conformation stability, in vitro digestibility and allergenicity of tropomyosin from shrimp (Exopalaemon modestus) as affected by high intensity ultrasound. Food Chem. 2018, 245, 997–1009. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Detection, identification, and quantification of oxidative protein modifications. J. Biol. Chem. 2019, 294, 19683–19708. [Google Scholar] [CrossRef] [Green Version]
- Requena, J.R.; Fu, M.X.; Ahmed, M.U.; Jenkins, A.J.; Lyons, T.J.; Baynes, J.W.; Thorpe, S.R. Quantification of malondialdehyde and 4-hydroxynonenal adducts to lysine residues in native and oxidized human low-density lipoprotein. Biochem. J. 1997, 322, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.K.; Graham, N.J.D. Oxidation of Amino Acids, Peptides and Proteins by Ozone: A Review. Ozone Sci. Eng. 2010, 32, 81–90. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, L.; Zhang, J.; Shi, J.; Wang, Y.; Wang, S. Adverse Effects of Thermal Food Processing on the Structural, Nutritional, and Biological Properties of Proteins. Annu. Rev. Food Sci. Technol. 2021, 12, 259–286. [Google Scholar] [CrossRef]
- Hellwig, M. Analysis of Protein Oxidation in Food and Feed Products. J. Agric. Food Chem. 2020, 68, 12870–12885. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, Q.; Hong, H.; Luo, Y. Tracking structural modifications and oxidative status of myofibrillar proteins from silver carp (Hypophthalmichthys molitrix) fillets treated by different stunning methods and in vitro oxidizing conditions. Food Chem. 2021, 365, 130510. [Google Scholar] [CrossRef] [PubMed]
- Mitra, B.; Lametsch, R.; Akcan, T.; Ruiz-Carrascal, J. Pork proteins oxidative modifications under the influence of varied time-temperature thermal treatments: A chemical and redox proteomics assessment. Meat Sci. 2018, 140, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Estévez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Li, Y.; Yang, L.; Wu, S.; Chen, J.; Lin, H. Structural, functional, rheological, and biological properties of the swim bladder collagen extracted from grass carp (Ctenopharyngodon idella). LWT 2022, 153, 112518. [Google Scholar] [CrossRef]
- Wang, L.; Yang, B.; Wang, R.; Du, X. Extraction of pepsin-soluble collagen from grass carp (Ctenopharyngodon idella) skin using an artificial neural network. Food Chem. 2008, 111, 683–686. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Modification | Amino Acid Residue | Position | Formula | Mass Shift (Da) |
|---|---|---|---|---|
| Monooxidation | HWMRYPNKDC | any | O | 15.99 |
| Dioxidation | YWFRKPCM | any | O(2) | 31.99 |
| Trioxidation | CWYF | any | O(3) | 47.98 |
| Tryptophan oxidation to kynurenine | W | any | C(−1)O | 3.99 |
| Oxidation to nitro | YW | any | H(−1)NO(2) | 44.99 |
| Tryptophan oxidation to hydroxykynurenin | W | any | C(−1)O(2) | 19.99 |
| Histidine oxidation to Aspartic Acid | H | any | H(−1)C(−2)N(−1)O | −22.03 |
| Histidine oxidation to Asparagine | H | any | H(−2)C(−2)N(−2)O(2) | −23.02 |
| Compounds | Pork | Fish | ||
|---|---|---|---|---|
| CW | OM | CW | OM | |
| TBRAS value | ND | ND | ND | ND |
| GO | 23.21 ± 1.90 | 17.13 ± 2.73 * | 27.60 ± 0.37 | 26.93 ± 0.55 |
| MGO | 72.23 ± 4.77 | 55.23 ± 7.58 * | 100.55 ± 4.18 | 118.93 ± 3.99 * |
| Free Amino Acid | Pork | Fish | ||
|---|---|---|---|---|
| CW | OM | CW | OM | |
| Asp | 13.28 ± 2.26 | 12.91 ± 0.15 | 10.44 ± 0.10 | 11.01 ± 0.94 |
| Ser | 39.58 ± 1.47 | 37.34 ± 0.70 | 34.73 ± 1.58 | 28.67 ± 1.61 * |
| Glu | 50.77 ± 4.40 | 56.11 ± 1.43 | 20.98 ± 0.96 | 24.61 ± 3.04 |
| Gly | 46.84 ± 1.10 | 53.01 ± 0.80 * | 484.23 ± 16.79 | 531.03 ± 9.21 * |
| His | 20.12 ± 1.24 | 19.17 ± 0.35 | 1121.46 ± 22.93 | 1145.86 ± 24.65 |
| Arg | 36.52 ± 6.61 | 42.91 ± 6.78 | 102.73 ± 6.02 | 116.61 ± 9.07 |
| Thr | 1735.14 ± 76.58 | 1736.48 ± 5.25 | ND | ND |
| Ala | 307.21 ± 9.41 | 354.44 ± 2.11 * | 176.12 ± 13.00 | 291.46 ± 31.39 * |
| Pro | 24.33 ± 3.17 | 66.95 ± 1.26 * | 138.46 ± 6.94 | 125.32 ± 13.42 |
| Cys | 30.28 ± 4.78 | 19.36 ± 0.39 * | ND | ND |
| Tyr | 23.12 ± 3.36 | 18.81 ± 0.44 | 4.45 ± 1.44 | 4.88 ± 0.76 |
| Val | 27.74 ± 6.66 | 25.20 ± 5.60 | 21.51 ± 2.13 | 22.19 ± 2.47 |
| Met | 12.38 ± 1.19 | 7.12 ± 0.20 * | 8.74 ± 1.16 | 8.51 ± 1.07 |
| Lys | 23.89 ± 2.39 | 13.76 ± 0.15 * | 139.4 ± 14.09 | 146.34 ± 17.03 |
| Ile | 17.38 ± 2.04 | 11.58 ± 0.29 * | 29.81 ± 2.49 | 23.75 ± 2.34 * |
| Leu | 30.55 ± 2.27 | 27.53 ± 0.48 | 44.29 ± 3.08 | 44.74 ± 3.53 |
| Phe | 14.67 ± 1.32 | 12.64 ± 0.50 | 15.39 ± 1.16 | 13.49 ± 0.57 |
| Total FAA | 2453.78 ± 98.51 | 2515.33 ± 7.25 | 2352.74 ± 87.92 | 2538.46 ± 108.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pian, M.-H.; Dong, L.; Yu, Z.-T.; Wei, F.; Li, C.-Y.; Fan, D.-C.; Li, S.-J.; Zhang, Y.; Wang, S. Ozone-Microbubble-Washing with Domestic Equipment: Effects on the Microstructure, and Lipid and Protein Oxidation of Muscle Foods. Foods 2022, 11, 903. https://doi.org/10.3390/foods11070903
Pian M-H, Dong L, Yu Z-T, Wei F, Li C-Y, Fan D-C, Li S-J, Zhang Y, Wang S. Ozone-Microbubble-Washing with Domestic Equipment: Effects on the Microstructure, and Lipid and Protein Oxidation of Muscle Foods. Foods. 2022; 11(7):903. https://doi.org/10.3390/foods11070903
Chicago/Turabian StylePian, Mei-Hui, Lu Dong, Zhen-Ting Yu, Fan Wei, Chun-Yang Li, Dan-Cai Fan, Shi-Jie Li, Yan Zhang, and Shuo Wang. 2022. "Ozone-Microbubble-Washing with Domestic Equipment: Effects on the Microstructure, and Lipid and Protein Oxidation of Muscle Foods" Foods 11, no. 7: 903. https://doi.org/10.3390/foods11070903