
Nutritional Quality of Four Novel Porridge Products Blended with Edible Cricket (Scapsipedus icipe) Meal for Food

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
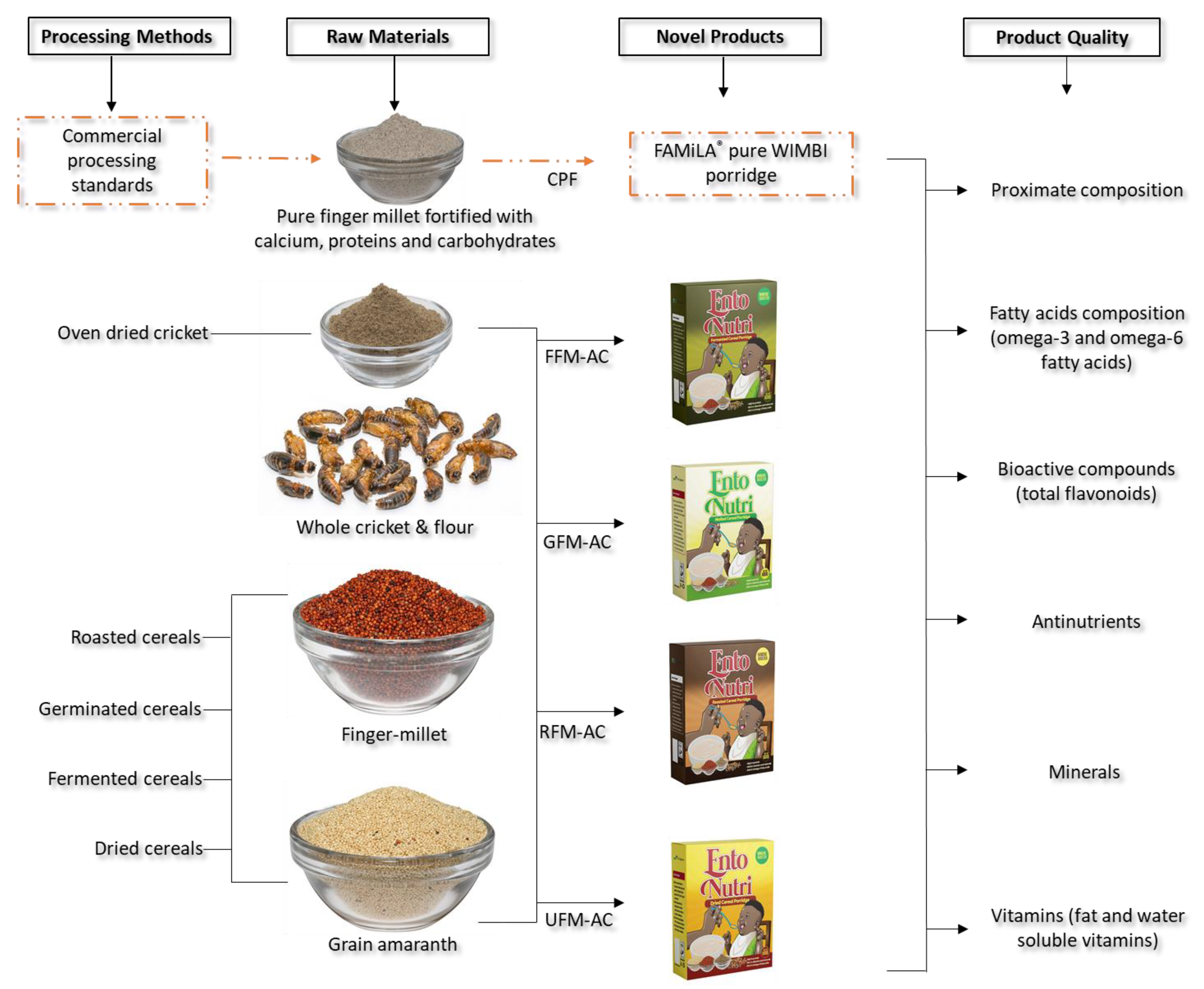
2.1. Raw Materials
2.2. Preparation of Raw Materials
2.2.1. Preparation of Crickets
2.2.2. Processing of Finger Millet and Amaranth Grains
2.3. Porridge Flour Formulations
2.4. Analysis of Proximate Composition
2.5. Determination of Mineral Composition
2.6. Assessment of Fatty Acids
2.6.1. Folch Extraction Method
2.6.2. Fatty Acid Determination
2.7. Determination of Water-Soluble Vitamins
2.8. Determination of Fat-Soluble Vitamins
2.9. Assessment of Phytic Acid, Tannins, and Flavonoids
2.10. Determination of Molar Ratios
2.11. Statistical Analysis
3. Results
3.1. Proximate Composition
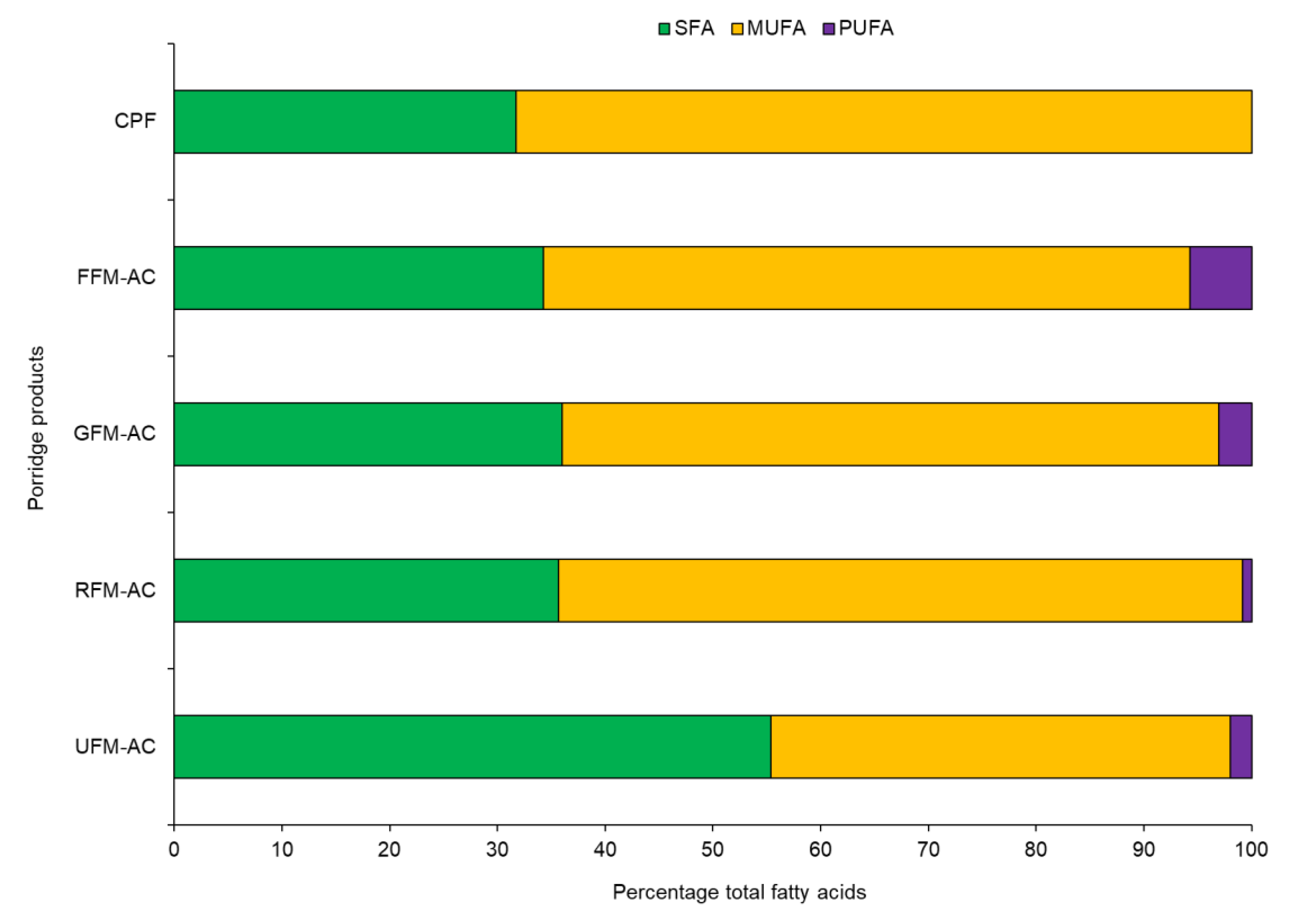
3.2. Fatty Acids
3.3. Vitamin Content
3.4. Mineral Content and Molar Ratios
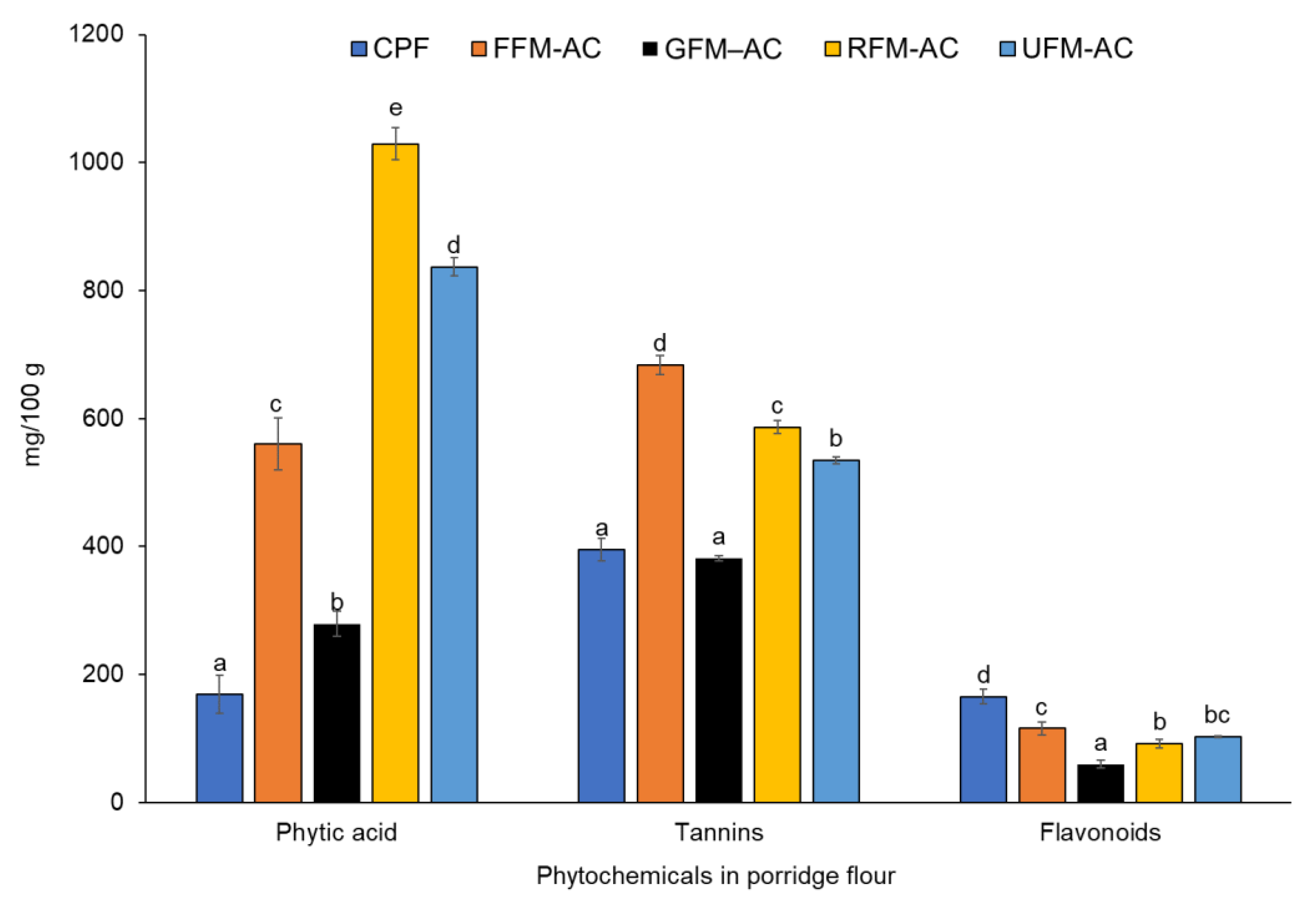
3.5. Phytic Acid, Tannins, and Flavonoids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wanjala, G.; Onyango, A.; Makayoto, M.; Onyango, C. Indigenous Technical Knowledge and Formulations of Thick (Ugali) and Thin (Uji) Porridges Consumed in Kenya. Afr. J. Food Sci. 2016, 10, 385–396. [Google Scholar] [CrossRef]
- Békés, F.; Schoenlechner, R.; Tömösközi, S. Ancient wheats and pseudocereals for possible use in cereal-grain dietary intolerances. In Cereal Grains; Wrigley, C., Ed.; Woodhead Publishing: Amsterdam, The Netherlands, 2017; pp. 353–389. [Google Scholar] [CrossRef]
- Liceaga, A.M. Processing insects for use in the food and feed industry. Curr. Opin. Insect. Sci. 2021, 48, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Nyangena, D.N.; Mutungi, C.; Imathiu, S.; Kinyuru, J.; Affognon, H.; Ekesi, S.; Nakimbugwe, D.; Fiaboe, K.K. Effects of Traditional Processing Techniques on the Nutritional and Microbiological Quality of Four Edible Insect Species Used for Food and Feed in East Africa. Foods 2020, 9, 574. [Google Scholar] [CrossRef] [PubMed]
- Fombong, F.T.; Van Der Borght, M.; Vanden Broeck, J. Influence of Freeze-Drying and Oven-Drying Post Blanching on the Nutrient Composition of the Edible Insect Ruspolia Differens. Insects 2017, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Dobermann, D.; Field, L.M.; Michaelson, L.V. Impact of Heat Processing on the Nutritional Content of Gryllus Bimaculatus (Black Cricket). Nutr. Bull. 2019, 44, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Mutungi, C.; Irungu, F.G.; Nduko, J.; Mutua, F.; Affognon, H.; Nakimbugwe, D.; Ekesi, S.; Fiaboe, K.K.M. Postharvest Processes of Edible Insects in Africa: A Review of Processing Methods, and the Implications for Nutrition, Safety and New Products Development. Crit. Rev. Food Sci. Nutr. 2019, 59, 276–298. [Google Scholar] [CrossRef] [Green Version]
- Ssepuuya, G.; Nakimbugwe, D.; De Winne, A.; Smets, R.; Claes, J.; Van Der Borght, M. Effect of Heat Processing on the Nutrient Composition, Colour, and Volatile Odour Compounds of the Long-Horned Grasshopper Ruspolia differens Serville. Food Res. Int. 2020, 129, 108831. [Google Scholar] [CrossRef]
- Mmari, M.W.; Kinyuru, J.N.; Laswai, H.S.; Okoth, J.K. Traditions, Beliefs and Indigenous Technologies in Connection with the Edible Longhorn Grasshopper Ruspolia differens (Serville 1838) in Tanzania. J. Ethnobiol. Ethnomed. 2017, 13, 60. [Google Scholar] [CrossRef] [Green Version]
- Onyango, C.; Okoth, M.W.; Mbugua, S.K. Effect of Drying Lactic Fermented Uji (an East African Sour Porridge) on Some Carboxylic Acids. J. Sci. Food Agric. 2000, 80, 1854–1858. [Google Scholar] [CrossRef]
- WHO; FAO. WHO|Vitamin and Mineral Requirements in Human Nutrition. World Health Organization WHO/FAO (2004), 2nd ed.; WHO: Geneva, Switzerland, 2004; Available online: https://apps.who.int/iris/handle/10665/42716 (accessed on 13 February 2022).
- WHO; FAO; UNU. Joint FAO/WHO/UNU Expert Consultation on Protein and Amino Acid Requirements in Human Nutrition (2002: Geneva, Switzerland); Food and Agriculture Organization of the United Nations, World Health Organization & United Nations University: Geneva, Switzerland, 2007. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 21st ed.; Latimer, G.W.J., Ed.; AOAC International: New York, NY, USA, 2019. [Google Scholar]
- Ochieng, J.; Schreinemachers, P.; Ogada, M.; Dinssa, F.F.; Barnos, W.; Mndiga, H. Adoption of improved amaranth varieties and good agricultural practices in East Africa. Land Use Policy 2019, 83, 187–194. [Google Scholar] [CrossRef]
- Yang, R.Y.; Keding, G.B. Nutritional Contributions of Important African Indigenous Vegetables. African Indigenous Vegetables in Urban Agriculture; Earthscan: London, UK, 2009; pp. 105–143. [Google Scholar]
- Petr, J.; Michalik, I.; Tlaskalova, H.; Capouchova, I.; Famera, O.; Urminska, D.; Tukova, L.; Knoblochova, H. Extension of the spectra of plant products for the diet in celiac disease. Czech J. Food Sci. 2003, 21, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Macharia-Mutie, C.W.; van de Wiel, A.M.; Moreno-Londono, A.M.; Mwangi, A.M.; Brouwer, I.D. Sensory acceptability and factors predicting the consumption of grain amaranth in Kenya. Ecol. Food Nutr. 2011, 50, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Karamać, M.; Gai, F.; Longato, E.; Meineri, G.; Janiak, M.A.; Amarowicz, R.; Peiretti, P.G. Antioxidant Activity and Phenolic Composition of Amaranth (Amaranthus caudatus) during Plant Growth. Antioxidants 2019, 8, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavez-Jauregui, R.N.; Silva, M.E.M.P.; Arěas, J.A.G. Extrusion cooking process for amaranth. J. Food Sci. 2000, 65, 1009–1015. [Google Scholar] [CrossRef]
- Olaniyi, J.O. Evaluation of yield and quality performance of grain amaranth varieties in the southwestern Nigeria. Res. J. Agron. 2007, 1, 42–45. [Google Scholar]
- Kolawole, E.L.; Sarah, O.A. Growth and yield performance of Amaranthus cruentus influenced by planting density and poultry manure application. Not. Bot. Horti Agrobot. Cluj-Napoca. 2009, 37, 195–199. [Google Scholar]
- Kim, H.K.; Kim, M.J.; Cho, H.Y.; Kim, E.-K.; Shin, D.H. Antioxidative and anti-diabetic effects of amaranth (Amaranthus esculantus) in streptozotocin-induced diabetic rats. Cell Biochem. Funct. 2006, 24, 195–214. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, M.J.; Shin, D.H. Improvement of lipid profile by amaranth (Amaranthus esculantus) supplementation in streptozotocin-induced diabetic rats. Ann. Nutr. Metab. 2006, 50, 277–281. [Google Scholar] [CrossRef]
- Bario, D.A.; Añón, M.C. Potential antitumor properties of a protein isolate obtained from the seeds of Amaranthus mantegazzianus. Eur. J. Nutr. 2010, 49, 73–82. [Google Scholar] [CrossRef]
- Aderibigbe, O.R.; Ezekiel, O.O.; Owolade, S.O.; Korese, J.K.; Sturm, B.; Hensel, O. Exploring the potentials of underutilized grain amaranth (Amaranthus spp.) along the value chain for food and nutrition security: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 656–669. [Google Scholar] [CrossRef]
- Okoth, J.K.; Ochola, S.; Gikonyo, N.K.; and Makokha, A.O. Efficacy of amaranth sorghum grains porridge in rehabilitating moderately acute malnourished children in a low-resource setting in Kenya: A randomized controlled trial. Integr. Food Nutr. Metal. 2017, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Cheseto, X.; Baleba, S.B.S.; Tanga, C.M.; Kelemu, S.; Torto, B. Chemistry and Sensory Characterization of a Bakery Product Prepared with Oils from African Edible Insects. Foods 2020, 9, 800. [Google Scholar] [CrossRef] [PubMed]
- Thermo Fisher Scientific. Determination of Water–and Fat–Soluble Vitamins by HPLC, Knowledge Creation Diffusion Utilization; Thermo Fisher Scientific: Waltham, MA, USA, 2010; pp. 1–23. [Google Scholar]
- Bhatnagar-Panwar, M.; Bhatnagar-Mathur, P.; VijayAnand Bhaaskarla, V.; Reddy Dumbala, S.; Sharma, K.K. Rapid, Accurate and Routine HPLC Method for Large-Scale Screening of pro-Vitamin A Carotenoids in Oilseeds. J. Plant Biochem. Biotechnol. 2015, 24, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Megazyme. Megazyme—Phytic Acid (Phytate)/Total Phosphoru Assay Kit Procedure; Megazyme: Wicklow, Ireland, 2017; Volume 17. [Google Scholar]
- Saxena, V.; Mishra, G.; Saxena, A.; Vishwakarma, K.K. A Comparative Study on Quantitative Estimation of Tannins in Terminalia Chebula, Terminalia Belerica, Terminalia Arjuna and Saraca Indica Using Spectrophotometer. Asian J. Pharm. Clin. Res. 2013, 6 (Suppl. 3), 148–149. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Norhaizan, M.E.; Nor Faizadatul Ain, A.W. Determination of Phytate, Iron, Zinc, Calcium Contents and Their Molar Ratios in Commonly Consumed Raw and Prepared Food in Malaysia. Malays. J. Nutr. 2009, 15, 213–222. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Anigo, K.; Ameh, D.; Ibrahim, S.; Danbauchi, S. Nutrient Composition of Complementary Food Gruels Formulated from Malted Cereals, Soybeans and Groundnut for Use in North-Western Nigeria. Afr. J. Food Sci. 2010, 4, 65–72. [Google Scholar]
- Dewey, K.G. The Challenge of Meeting Nutrient Needs of Infants and Young Children during the Period of Complementary Feeding: An Evolutionary Perspective. J. Nutr. 2013, 143, 2050–2054. [Google Scholar] [CrossRef] [Green Version]
- Murugu, D.K.; Onyango, A.N.; Ndiritu, A.K.; Osuga, I.M.; Xavier, C.; Nakimbugwe, D.; Tanga, C.M. From Farm to Fork: Crickets as Alternative Source of Protein, Minerals, and Vitamins. Front. Nutr. 2021, 505, 704002. [Google Scholar] [CrossRef]
- Melgar-Lalanne, G.; Hern’andez-´Alvarez, A.; Salinas-Castro, A. Edible insects processing: Traditional and innovative technologies. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1166–1191. [Google Scholar] [CrossRef] [Green Version]
- Lillioja, S.; Neal, A.L.; Tapsell, L.; Jacobs, D.R. Whole grains, type 2 diabetes, coronary heart disease, and hypertension: Links to the aleurone preferred over indigestible fiber. BioFactors 2013, 39, 242–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onyango, C.A.; Ochanda, S.O.; Mwasaru, M.A.; Ochieng, J.K.; Mathooko, F.M.; Kinyuru, J.N. Effects of Malting and Fermentation on Anti-Nutrient Reduction and Protein Digestibility of Red Sorghum, White Sorghum and Pearl Millet. J. Food Res. 2013, 2, 41. [Google Scholar] [CrossRef]
- Min, Z.; Chen, H.; Li, J.; Pei, Y.; Liang, Y. Antioxidant properties of tartary buckwheat extracts as affected by different thermal processing methods. LWT Food Sci. Technol. 2010, 43, 181–185. [Google Scholar]
- Masarirambi, M.; Mavuso, V.; Songwe, V.; Nkambule, T.; and Mhazo, N. Indigenous Post-Harvest Handling and Processing of Traditional Vegetables in Swaziland: A Review. Afr. J. Agric. Res. 2010, 5, 3333–3341. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Luchuo, E.B.; Paschal, K.A.; Geraldine, N.; Kindong, N.P.; Nsah, Y.S.; Tanjeko, A.T. Malnutrition in Sub—Saharan Africa: Burden, causes and prospects. Pan Afr Med. J. 2013, 15, 1–20. [Google Scholar]
- Branca, F.; Demaio, A.; Udomkesmalee, E.; Baker, P.; Aguayo, V.M.; Barquera, S.; Dain, K.; Keir, L.; Lartey, A.; Mugambi, G. Dynamics of the double burden of malnutrition and the changing nutrition reality. Lancet 2020, 395, 65–74. [Google Scholar]
- Agbemafle, I.; Hadz, D.; Amagloh, F.K.; Zotor, F.B.; Reddy, M.B. Orange-Fleshed Sweet Potato and Edible Insects. Foods 2020, 9, 1225. [Google Scholar] [CrossRef]
- Magara, H.J.O.; Niassy, S.; Ayieko, M.A.; Mukundamago, M.; Egonyu, J.P.; Tanga, C.M.; Kimathi, E.K.; Ongere, J.O.; Fiaboe, K.K.M.; Hugel, S.; et al. Edible Crickets (Orthoptera) Around the World: Distribution, Nutritional Value, and Other Benefits—A Review. Front. Nutr. 2021, 7, 257. [Google Scholar] [CrossRef]
- Pranoto, Y.; Anggrahini, S.; Efendi, Z. Effect of Natural and Lactobacillus Plantarum Fermentation on In-Vitro Protein and Starch Digestibilities of Sorghum Flour. Food Biosci. 2013, 2, 46–52. [Google Scholar] [CrossRef]
- Osman, M.A. Effect of Traditional Fermentation Process on the Nutrient and Antinutrient Contents of Pearl Millet during Preparation of Lohoh. J. Saudi Soc. Agric. Sci. 2011, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Nnam, N.M. Chemical Evaluation of Multimixes Formulated from Some Local Staples for Use as Complementary Foods in Nigeria. Plant Foods Hum. Nutr. 2000, 55, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Onabanjo, O.O.; Oguntona, C.R.B.; Maziya-Dixon, B.; Olayiwola, I.O.; Oguntona, E.B.; Dixon, A.G.O. Nutritional Evaluation of Four Optimized Cassava-Based Complementary Foods. Afr. J. Food Sci. 2008, 2, 136–142. [Google Scholar]
- Assohoun, M.C.N.; Djeni, T.N.; Koussémon-Camara, M.; Brou, K. Effect of Fermentation Process on Nutritional Composition and Aflatoxins Concentration of Doklu, a Fermented Maize Based Food. Food Nutr. Sci. 2013, 4, 1120–1127. [Google Scholar] [CrossRef] [Green Version]
- Ikenebomah, M.; Kok, J.R.; Ingram, J. Processing and Fermentation of the African Locust Bean (Parkia Felocoidea) Can. Int. J. Food Sci. Technol. 2019, 17, 48–50. [Google Scholar]
- Codex Alimentarius. Guidelines for Development of Supplementary Foods for Older Infants and Children; FAO: Rome, Italy, 1991. [Google Scholar]
- Islam, M.S.; Castellucci, C.; Fiorini, R.; Greco, S.; Gagliardi, R.; Zannotti, A.; Giannubilo, S.R.; Ciavattini, A.; Frega, N.G.; Pacetti, D.; et al. Omega-3 Fatty Acids Modulate the Lipid Profile, Membrane Architecture, and Gene Expression of Leiomyoma Cells. J. Cell. Physiol. 2018, 233, 7143–7156. [Google Scholar] [CrossRef]
- Martínez Andrade, K.A.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [Green Version]
- Stark, K.D.; Van Elswyk, M.E.; Higgins, M.R.; Weatherford, C.A.; Salem, N. Global Survey of the Omega-3 Fatty Acids, Docosahexaenoic Acid and Eicosapentaenoic Acid in the Blood Stream of Healthy Adults. Prog. Lipid Res. 2016, 63, 132–152. [Google Scholar] [CrossRef]
- Gunaratne, A.W.; Makrides, M.; Collins, C.T. Maternal Prenatal and/or Postnatal n-3 Fish Oil Supplementation for Preventing Allergies in Early Childhood. Cochrane Database Syst. Rev. 2012, 9, 1–23. [Google Scholar] [CrossRef]
- Paucean, A.; Moldovan, O.P.; Mureșan, V.; Socaci, S.A.; Dulf, F.V.; Alexa, E.; Man, S.M.; Mureșan, A.E.; Muste, S. Folic Acid, Minerals, Amino-Acids, Fatty Acids and Volatile Compounds of Green and Red Lentils. Folic Acid Content Optimization in Wheat-Lentils Composite Flours. Chem. Cent. J. 2018, 12, 88. [Google Scholar] [CrossRef] [Green Version]
- Kannan, N.; Rao, A.S.; Nair, A. Microbial Production of Omega-3 Fatty Acids: An Overview. J. Appl. Microbiol. 2021, 131, 2114–2130. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Abe, S.; Hirakawa, Y.; Takagi, S. Roasting Effects on Fatty Acid Distributions of Triacylglycerols and Phospholipids in Sesame (Sesamum Indicum) Seeds. J. Sci. Food Agric. 2001, 81, 620–626. [Google Scholar] [CrossRef]
- Mariod, A.A.; Edris, Y.A.; Cheng, S.F.; Abdelwahab, S.I. Effect of Germination Periods and Conditions on Chemical Composition, Fatty Acids and Amino Acids of Two Black Cumin Seeds. Acta Sci. Pol. Technol. Aliment. 2012, 11, 401–410. [Google Scholar]
- Difo, H.V.; Onyike, E.; Ameh, D.A.; Ndidi, U.S.; Njoku, G.C. Chemical Changes during Open and Controlled Fermentation of Cowpea (Vigna Unguiculata) Flour. Int. J. Food Nutr. Saf. 2014, 5, 1–10. [Google Scholar]
- Tizazu, S.; Urga, K.; Abuye, C.; Retta, N. Improvement of Energy and Nutrient Density of Sorghumbased Complementary Foods Using Germination. Afr. J. Food Agric. Nutr. Dev. 2010, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Walther, B.; Schmid, A. Effect of Fermentation on Vitamin Content in Food; Elsevier Inc.: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Barrios-González, J. Solid-State Fermentation: Physiology of Solid Medium, Its Molecular Basis and Applications. Process Biochem. 2012, 47, 175–185. [Google Scholar] [CrossRef]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Somboonpanyakul, P. B Vitamins and Prebiotic Fructooligosaccharides of Cashew Apple Fermented with Probiotic Strains Lactobacillus Spp., Leuconostoc Mesenteroides and Bifidobacterium Longum. Process Biochem. 2018, 70, 9–19. [Google Scholar] [CrossRef]
- Tabaszewska, M.; Gabor, A.; Jaworska, G.; Drożdż, I. Effect of Fermentation and Storage on the Nutritional Value and Contents of Biologically-Active Compounds in Lacto-Fermented White Asparagus (Asparagus Officinalis L.). LWT Food Sci. Technol. 2018, 92, 67–72. [Google Scholar] [CrossRef]
- Zilic, S.; Delic, N.; Basic, Z.; Ignjatovic-Micic, D.; Jankovic, M.; Vancetovic, J. Effects of Alkaline Cooking and Sprouting on Bioactive Compounds, Their Bioavailability and Relation to Antioxidant Capacity of Maize Flour. J. Food Nutr. Res. 2015, 54, 155–164. [Google Scholar]
- Huang, X.; Cai, W.; Xu, B. Kinetic Changes of Nutrients and Antioxidant Capacities of Germinated Soybean (Glycine Max l.) and Mung Bean (Vigna Radiata L.) with Germination Time. Food Chem. 2014, 143, 268–276. [Google Scholar] [CrossRef]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.B. Fermentation and Germination Improve Nutritional Value of Cereals and Legumes through Activation of Endogenous Enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuliaş, A.; Vlase, G.; Vlase, T.; Oneţiu, D.; Doca, N.; Ledeţi, I. Thermal Degradation of B-Group Vitamins: B1, B2 and B6: Kinetic Study. J. Therm. Anal. Calorim. 2014, 118, 1033–1038. [Google Scholar] [CrossRef]
- Young, H.; Guk, I.; Myoung, T.; Sik, K.; Sik, D.; Hyun, J.; Joong, D.; Lee, J.; Ri, Y.; Sang, H. Chemical and Functional Components in Different Parts of Rough Rice (Oryza Sativa L.) before and after Germination. Food Chem. 2012, 134, 288–293. [Google Scholar] [CrossRef]
- Alamprese, C.; Ratti, S.; Rossi, M. Effects of Roasting Conditions on Hazelnut Characteristics in a Two-Step Process. J. Food Eng. 2009, 95, 272–279. [Google Scholar] [CrossRef]
- Stuetz, W.; Schlörmann, W.; Glei, M. B-Vitamins, Carotenoids and Tocopherols in Nuts. Food Chem. 2016, 221, 222–227. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Stockmann, R.; Ng, K.; Ajlouni, S. Revisiting Phytate-Element Interactions: Implications for Iron, Zinc and Calcium Bioavailability, with Emphasis on Legumes. Crit. Rev. Food Sci. Nutr. 2020, 62, 1696–1712. [Google Scholar] [CrossRef]
- Konietzny, U.; Greiner, R. PHYTIC ACID| Properties and Determination. Am. J. Med. Sci 2003, 317, 370–376. [Google Scholar]
- Castro-Alba, V.; Lazarte, C.E.; Perez-Rea, D.; Carlsson, N.G.; Almgren, A.; Bergenståhl, B.; Granfeldt, Y. Fermentation of Pseudocereals Quinoa, Canihua, and Amaranth to Improve Mineral Accessibility through Degradation of Phytate. J. Sci. Food Agric. 2019, 99, 5239–5248. [Google Scholar] [CrossRef] [Green Version]
- Inyang, C.U.; Zakari, U.M. Effect of Germination and Fermentation of Pearl Millet on Proximate Chemical and Sensory Properties of Instant “Fura”—A Nigerian Cereal Food. Pak. J. Nutr. 2008, 7, 9–12. [Google Scholar] [CrossRef]
- García-Mantrana, I.; Monedero, V.; Haros, M. Application of Phytases from Bifidobacteria in the Development of Cereal-Based Products with Amaranth. Eur. Food Res. Technol. 2014, 238, 853–862. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, S.S.; Salunke, D.K. Grain legumes, seeds and nuts: Rationale for fermentation. Fermented grains legumes, seeds and nuts: A global perspective. FAO Agric. Serv. Bull. 2002, 142, 1–32. [Google Scholar]
- Frontela, C.; García-Alonso, F.J.; Ros, G.; Martínez, C. Phytic Acid and Inositol Phosphates in Raw Flours and Infant Cereals: The Effect of Processing. J. Food Compos. Anal. 2008, 21, 343–350. [Google Scholar] [CrossRef]
- Greiner, R.; Konietzny, U. Phytase for Food Application Phytase for Food Application. J. Food Technol. Biotechnol. 2006, 44, 125–140. [Google Scholar]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adetuyi, F.O.; Ibrahim, T.A. Effect of Fermentation Time on the Phenolic, Flavonoid and Vitamin C Contents and Antioxidant Activities of Okra (Abelmoschus Esculentus) Seeds. Niger. Food J. 2014, 32, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of Fermentation on the Antioxidant Activity in Plant-Based Foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Chaaban, H.; Ioannou, I.; Chebil, L.; Slimane, M.; Gérardin, C.; Paris, C.; Charbonnel, C.; Chekir, L.; Ghoul, M. Effect of Heat Processing on Thermal Stability and Antioxidant Activity of Six Flavonoids. J. Food Process. Preserv. 2017, 41, 1–12. [Google Scholar] [CrossRef]
- Modgil, R.; Sood, P. Effect of Roasting and Germination on Carbohydrates and Anti-Nutritional Constituents of Indigenous and Exotic Cultivars of Pseudo-Cereal (Chenopodium). J. Life Sci. 2017, 9, 64–70. [Google Scholar] [CrossRef]
- Ali, M.A.M.; Tinay, A.H.; Tinay, E.l.; Abdalla, A.H. Effect of Fermentation on the in Vitro Protein Digestibility of Pearl Millet. Food Chem. 2003, 80, 51–54. [Google Scholar] [CrossRef]
- Shimelis, E.A.; Rakshit, S.K. Effect of Processing on Antinutrients and in Vitro Protein Digestibility of Kidney Bean (Phaseolus Vulgaris L.) Varieties Grown in East Africa. Food Chem. 2007, 103, 161–172. [Google Scholar] [CrossRef]
- Kunyanga, C.N.; Imungi, J.K.; Okoth, M.; Momanyi, C.; Biesalski, H.K.; Vadivel, V. Antioxidant and Antidiabetic Properties of Condensed Tannins in Acetonic Extract of Selected Raw and Processed Indigenous Food Ingredients from Kenya. J. Food Sci. 2011, 76, C560–C567. [Google Scholar] [CrossRef] [PubMed]
- Abdelhaleem, H.; Tinay, A.H.; El Mustafa, A.I.; Babiker, E.E. Effect of Fermentation, Malt-Pretreatment and Cooking on Antinutritional Factors and Protein Digestibility of Sorghum Cultivars. Pak. J. Nutr. 2008, 7, 335–341. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Products | Proximate Composition | ||||||
|---|---|---|---|---|---|---|---|
| Moisture (%) | Ash (g/100 g) | Fiber (g/100 g) | Protein (g/100 g) | Fat (g/100 g) | CHO (g/100 g) | Energy (kcal/100 g) | |
| CPF | 11.76 ± 0.25 e | 2.57 ± 0.04 ab | 4.79 ± 0.28 c | 8.55 ± 0.16 a | 2.14 ± 0.16 a | 81.94 ± 0.26 d | 381.28 ± 0.35 a |
| FFM–AC | 4.84 ± 0.25 b | 2.22 ± 0.15 a | 3.33 ± 0.29 a | 15.34 ± 0.17 b | 7.22 ± 0.06 b | 71.88 ± 0.29 c | 413.92 ± 0.80 c |
| GFM–AC | 5.92 ± 0.42 c | 2.88 ± 0.48 b | 5.34 ± 0.02 d | 16.12 ± 0.15 d | 8.19 ± 0.09 c | 67.46 ± 0.39 a | 408.12 ± 2.27 b |
| RFM–AC | 3.02 ± 0.22 a | 2.76 ± 0.05 ab | 3.88 ± 0.04 b | 15.54 ± 0.16 bc | 8.08 ± 0.28 c | 69.74 ± 0.13 b | 413.83 ± 1.44 c |
| UFM–AC | 7.44 ± 0.26 d | 2.83 ± 0.09 ab | 4.64 ± 0.11 c | 15.90 ± 0.28 cd | 8.31 ± 0.38 c | 68.31 ± 0.71 a | 411.68 ± 1.71 bc |
| Peak No. | tR (min) | Compound Name | ω-n(Δn) | CPF | FFM-AC | GFM-AC | RFM-AC | UFM-AC |
|---|---|---|---|---|---|---|---|---|
| 1 | 18.96 | Methyl Dodecanoate | C12:0 | - | 0.67 ± 0.09 | 0.27 ± 0.01 | 0.58 ± 0.05 | 0.35 ± 0.02 |
| 2 | 19.72 | Methyl 11-Methyldodecanoate | Iso-methyl-C12:0 | - | 0.05 ± 0.01 | 0.06 ± 0.00 | - | |
| 3 | 20.12 | Methyl Tridecanoate | C13:0 | - | 0.12 ± 0.01 | 0.06 ± 0.00 | 0.09 ± 0.01 | 0.66 ± 0.00 |
| 4 | 20.82 | Methyl 12-Methyltridecanoate | Iso-methyl-C13:0 | - | 0.21 ± 0.02 | 0.12 ± 0.00 | 0.07 ± 0.01 | 0.13 ± 0.01 |
| 5 | 21.22 | Methyl Tetradecanoate | C14:0 | 0.85 ± 0.04 | 14.72 ± 0.84 | 6.09 ± 0.10 | 12.04 ± 1.21 | 8.26 ± 0.97 |
| 6 | 21.78 | Methyl 4-Methyldodecanoate | Iso-methyl-C12:0 | - | 0.72 ± 0.07 | 0.44 ± 0.00 | - | - |
| 7 | 22.00 | Methyl 13-Methyltetradecanoate | Iso-methyl-C14:0 | - | 3.64 ± 0.21 | 1.98 ± 0.01 | 1.42 ± 0.17 | 0.89 ± 0.17 |
| 8 | 22.00 | Methyl 12-Methyltetradecanoate | Iso-methyl-C14:0 | - | 0.85 ± 0.04 | 0.54 ± 0.00 | 1.84 ± 0.10 | 0.29 ± 0.19 |
| 9 | 22.29 | Methyl Pentadecanoate | C15:0 | 0.38 ± 0.04 | 3.79 ± 0.28 | 1.61 ± 0.00 | 3.20 ± 0.48 | 2.93 ± 0.56 |
| 10 | 22.74 | Methyl 5,9,13-Trimethyltetradecanoate | Iso-trimethyl-C14:0 | - | 0.45 ± 0.05 | 0.00 ± 0.00 | - | - |
| 11 | 22.94 | Methyl 14-Methylpentadecanoate | Iso-methyl-C15:0 | - | 1.22 ± 0.10 | 0.85 ± 0.01 | 1.24 ± 0.07 | 0.94 ± 0.06 |
| 12 | 23.37 | Methyl Hexadecanoate | C16:0 | 32.22 ± 1.37 | 200.94 ± 6.55 | 255.32 ± 7.40 | 559.63 ± 50.55 | 319.22 ± 28.46 |
| 13 | 23.93 | Methyl 15-Methylhexadecanoate | Iso-methyl-C16:0 | - | 3.89 ± 0.38 | 2.90 ± 0.06 | 3.09 ± 0.42 | 3.15 ± 0.39 |
| 14 | 24.02 | Methyl 14-Methylhexadecanoate | Iso-methyl-C16:0 | 1.30 ± 0.13 | 11.82 ± 0.94 | 5.85 ± 0.12 | 12.80 ± 1.11 | 6.23 ± 0.88 |
| 15 | 24.29 | Methyl Heptadecanoate | C17:0 | 1.11 ± 0.22 | 9.92 ± 0.71 | 3.08 ± 0.05 | 7.40 ± 0.68 | 4.58 ± 0.91 |
| 16 | 24.69 | Methyl 14-Methylheptadecanoate | Iso-methyl-C17:0 | - | 1.40 ± 0.18 | 1.43 ± 0.02 | 2.99 ± 0.24 | 1.66 ± 0.20 |
| 17 | 25.25 | Methyl Octadecanoate | C18:0 | 13.02 ± 0.51 | 170.54 ± 2.98 | 78.14 ± 1.41 | 36.71 ± 2.68 | 114.00 ± 23.09 |
| 18 | 26.12 | Methyl Nonadecanoate | C19:0 | - | 2.08 ± 0.14 | 1.46 ± 0.05 | 1.92 ± 0.06 | 1.44 ± 0.30 |
| 19 | 26.98 | Methyl Eicosanoate | C20:0 | 3.05 ± 0.06 | 13.07 ± 0.15 | 11.07 ± 0.00 | 26.99 ± 1.49 | 16.48 ± 2.30 |
| 20 | 27.58 | Methyl 18-Methyleicosanoate | Iso-methyl-C20:0 | - | 4.90 ± 0.78 | 3.45 ± 0.00 | 5.11 ± 0.76 | 2.80 ± 0.17 |
| 21 | 27.80 | Methyl Heneicosanoate | C21:0 | - | 2.30 ± 0.23 | 3.34 ± 0.30 | 3.75 ± 0.66 | 1.79 ± 0.10 |
| 22 | 28.59 | Methyl Docosanoate | C22:0 | 2.47 ± 0.02 | 5.35 ± 0.13 | 6.30 ± 0.13 | 9.01 ± 0.45 | 6.92 ± 0.85 |
| 23 | 29.37 | Methyl Tricosanoate | C23:0 | - | 3.95 ± 0.09 | 3.93 ± 0.01 | 4.63 ± 0.39 | 2.89 ± 0.21 |
| 24 | 30.13 | Methyl Tetracosanoate | C24:0 | - | 9.45 ± 0.54 | 6.07 ± 0.04 | 10.98 ± 0.48 | 4.43 ± 1.23 |
| ∑ SFA | ||||||||
| 25 | 20.95 | Methyl 11Z-Tetradecenoate | C14:1 (n-3) | - | 0.52 ± 0.01 | - | 0.93 ± 0.09 | - |
| 26 | 21.08 | Methyl 9Z-Tetradecenoate | C14:1 (n-3) | - | 0.89 ± 0.09 | - | 0.67 ± 0.07 | - |
| 27 | 23.12 | Methyl 9Z-Hexadecenoate | C16:1 (n-7) | 2.24 ± 0.26 | 55.80 ± 1.46 | 18.50 ± 0.16 | 67.70 ± 1.84 | 31.80 ± 2.43 |
| 28 | 24.09 | Methyl 10Z-Heptadecenoate | C17:1 (n-7) | - | 7.42 ± 1.10 | 1.37 ± 0.03 | 1.75 ± 0.16 | 0.81 ± 0.08 |
| 29 | 25.00 | Methyl 11-Octadecenoate | C18:1 (n-9) | - | 3.69 ± 0.18 | 3.48 ± 0.11 | 2.83 ± 0.19 | 3.72 ± 0.12 |
| 30 | 25.07 | Methyl 9E-Octadecenoate | C18:1 (n-9) | 111.68 ± 6.88 | 729.29 ± 41.54 | 630.70 ± 12.89 | 1149.35 ± 93.82 | 332.62 ± 40.54 |
| 31 | 25.88 | Methyl 10-Nonadecenoate | C19:1 (n-9) | - | 2.69 ± 0.14 | 3.81 ± 0.17 | 8.99 ± 0.45 | 4.55 ± 0.46 |
| 32 | 26.77 | Methyl 11Z-Eicosenoate | C20:1 (n-9) | 2.94 ± 0.14 | 8.63 ± 0.55 | 8.52 ± 0.16 | 19.96 ± 0.57 | 11.31 ± 2.20 |
| 33 | 28.41 | Methyl 11-Docosenoate | C22:1 (n-11) | - | 4.23 ± 0.20 | - | 3.27 ± 0.30 | - |
| 34 | 29.95 | Methyl 15Z-Tetracosenoate | C24:1 (n-9) | - | 3.31 ± 0.16 | - | - | - |
| ∑ MUFA | ||||||||
| 35 | 24.74 | Methyl 9Z,12Z-Octadecadienoate | C18:2 (n-6) | - | 74.70 ± 4.02 | 38.38 ± 2.09 | 16.69 ± 1.10 | 22.62 ± 2.07 |
| 36 | 24.76 | methyl 6Z,9Z,12Z-Octadecatrienoate | C18:3 (n-3) | - | 4.60 ± 0.43 | 3.48 ± 0.40 | 1.67 ± 0.15 | 2.80 ± 0.03 |
| 37 | 25.41 | Methyl 7,12-Octadecadienoate | C18:2 (n-7) | - | 8.87 ± 0.66 | 11.07 ± 0.94 | 6.55 ± 1.96 | 5.22 ± 0.27 |
| 38 | 25.79 | Methyl 9Z,11E-Octadecadienoate | C18:2 (n-7) | - | 2.39 ± 0.09 | 2.60 ± 0.42 | 1.62 ± 0.21 | 1.35 ± 0.01 |
| 39 | 26.24 | Methyl 9Z,11E,13E-Octadecatrienoate (α-ESA) | C18:3 (n-3) | - | 1.03 ± 0.04 | 0.92 ± 0.10 | 0.57 ± 0.00 | 0.74 ± 0.03 |
| 40 | 26.26 | Methyl 9Z,12Z,15Z-Octadecatrienoate (ALA) | C18:3 (n-3) | - | 7.01 ± 0.23 | 7.10 ± 0.61 | 3.79 ± 0.01 | 5.07 ± 0.02 |
| 41 | 26.44 | Methyl 5Z,8Z,11Z,14Z-Eicosatetraenoate (AA) | C20:4 (n-6) | - | 1.55 ± 0.09 | 2.76 ± 0.06 | 1.89 ± 0.04 | 1.95 ± 0.19 |
| 42 | 26.50 | Methyl 5Z,8Z,11Z,14Z,17Z-Eicosapentaenoate (EPA) | C20:5 (n-3) | - | 13.07 ± 0.77 | 4.44 ± 0.02 | - | - |
| 43 | 26.64 | Methyl 8,11,14,17-Eicosatetraenoate (AA) | C20:4 (n-6) | - | 2.040 ± 0.10 | - | - | - |
| 44 | 28.07 | Methyl 4Z,7Z,10Z,13Z,16Z,19Z-Docosahexaenoate (DHA) | C22:6 (n-3) | - | 2.40 ± 0.34 | - | - | - |
| ∑ PUFA | - | 117.66 ± 3.17 | 70.76 ± 1.30 | 32.68 ± 1.72 | 39.73 ± 2.16 | |||
| ∑ n-6 PUFA | - | 89.55 ± 4.42 | 54.82 ± 0.92 | 26.65 ± 1.68 | 31.13 ± 2.15 | |||
| ∑ n-3 PUFA | - | 28.11 ± 0.75 | 15.94 ± 0.99 | 6.03 ± 1.15 | 8.60 ± 0.01 | |||
| ∑ n-6/n-3 | - | 3.2 | 3.4 | 4.4 | 3.6 | |||
| ∑ ALA + EPA + DHA | - | 27.08 ± 0.70 | 15.02 ± 1.00 | 5.47 ± 1.15 | 7.87 ± 0.04 |
| Vitamins | Porridge Products | ||||
|---|---|---|---|---|---|
| CPF | FFM–AC | GFM–AC | RFM–AC | UFM–AC | |
| Vitamin C | 149.6 ± 2.1 c | 146.5 ± 2.8 c | 72.0 ± 6.5 b | 55.2 ± 2.9 a | 58.0 ± 5.1 a |
| Thiamine (B1) | – | 39.5 ± 3.0 c | – | 4.3 ± 0.2 b | 5.9 ± 0.4 b |
| Nicotinic acid (B3) | 27.7 ± 2.9 e | – | 19.5 ± 1.2 d | 6.1 ± 0.5 b | 10.6 ± 0.7 c |
| Pyridoxine (B6) | 0.5 ± 0.1 a | 10.8 ± 1.1 c | 6.0 ± 0.3 b | – | – |
| Nicotinamide | 3.0 ± 0.3 a | 47.9 ± 2.1 d | 33.9 ± 1.1 c | 7.1 ± 0.2 b | 8.8 ± 0.4 b |
| Pantothenic acid (B5) | 26.4 ± 2.5 a | 423.3 ± 3.4 d | 453.8 ± 44.5 d | 314.4 ± 18.4 c | 209.6 ± 3.8 b |
| Folate (B9) | 28.6 ± 2.6 a | 38.8 ± 1.5 b | 42.4 ± 3.7 b | 41.8 ± 0.3 b | 29.3 ± 1.8 a |
| Cyanocobalamin (B12) | 3.2 ± 0.3 a | 21.9 ± 1.4 c | 13.7 ± 4.0 b | 12.4 ± 0.9 b | 37.7 ± 4.0 d |
| Riboflavin (B2) | 74.2 ± 8.2 b | 41.6 ± 1.0 a | 34.8 ± 2.9 a | 45.5 ± 2.4 a | 41.6 ± 5.0 b |
| Retinol | 0.55 ± 0.03 c | 0.54 ± 0.08 c | 0.07 ± 0.01 a | 0.29 ± 0.01 ab | 0.38 ± 0.19 bc |
| γ-Tocopherols | 0.88 ± 0.04 c | 0.17 ± 0.01 a | 0.19 ± 0.01 a | 0.54 ±0.00 b | 0.52 ± 0.02 b |
| α-Tocopherols | 0.46 ± 0.06 a | 0.35 ± 0.09 a | 1.48 ± 0.03 c | 0.76 ± 0.08 b | 0.83 ± 0.03 b |
| Minerals | Porridge Products | ||||
|---|---|---|---|---|---|
| CPF | FFM–AC | GFM–AC | RFM–AC | UFM–AC | |
| Mg (mg/100 g) | 145.08 ± 0.25 a | 169.57 ± 8.03 b | 207.82 ± 11.93 c | 210.84 ± 1.45 c | 203.09 ± 5.14 c |
| Fe (mg/100 g) | 9.86 ± 2.08 a | 8.56 ± 1.45 a | 19.48 ± 6.69 b | 9.18 ± 1.18 a | 9.55 ± 2.42 a |
| Ca (mg/100 g) | 312.69 ± 0.57 c | 234.87 ± 17.60 a | 278.61 ± 17.90 b | 257.69 ± 1.37 ab | 244.69 ± 4.94 a |
| Zn (mg/100 g) | 1.86 ± 0.04 a | 3.23 ± 0.28 b | 3.71 ± 0.18 b | 3.39 ± 0.31 b | 3.08 ± 0.16 b |
| P (mg/100 g) | 221.63 ± 5.57 a | 372.71 ± 19.14 b | 469.28 ± 9.55 c | 458.70 ± 3.76 c | 476.72 ± 17.46 c |
| Mn (mg/100 g) | 22.44 ± 0.20 c | 10.92 ± 0.63 b | 9.32 ± 0.67 a | 9.45 ± 0.15 a | 8.64 ± 0.04 a |
| Cu (µg/100 g) | 477.42 ± 1.88 a | 728.78 ± 15.37 b | 787.20 ± 42.28 b | 724.94 ± 22.80 b | 736.28 ± 14.47 b |
| Molar ratios (Bioavailability) | |||||
| Phy:Fe | 1.46 ± 0.05 a | 5.69 ± 1.38 b | 1.31 ± 0.42 a | 9.63 ± 1.41 c | 7.76 ± 1.90 bc |
| Phy:Zn | 9.03 ± 1.97 a | 17.27 ± 1.40 b | 7.43 ± 0.64 a | 30.48 ± 5.39 c | 26.90 ± 2.04 c |
| Phy:Ca | 0.03 ± 0.01 a | 0.14 ± 0.00 c | 0.06 ± 0.01 a | 0.24 ± 0.01 e | 0.21 ± 0.00 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maiyo, N.C.; Khamis, F.M.; Okoth, M.W.; Abong, G.O.; Subramanian, S.; Egonyu, J.P.; Xavier, C.; Ekesi, S.; Omuse, E.R.; Nakimbugwe, D.; et al. Nutritional Quality of Four Novel Porridge Products Blended with Edible Cricket (Scapsipedus icipe) Meal for Food. Foods 2022, 11, 1047. https://doi.org/10.3390/foods11071047
Maiyo NC, Khamis FM, Okoth MW, Abong GO, Subramanian S, Egonyu JP, Xavier C, Ekesi S, Omuse ER, Nakimbugwe D, et al. Nutritional Quality of Four Novel Porridge Products Blended with Edible Cricket (Scapsipedus icipe) Meal for Food. Foods. 2022; 11(7):1047. https://doi.org/10.3390/foods11071047
Chicago/Turabian StyleMaiyo, Nelly C., Fathiya M. Khamis, Michael W. Okoth, George O. Abong, Sevgan Subramanian, James P. Egonyu, Cheseto Xavier, Sunday Ekesi, Evanson R. Omuse, Dorothy Nakimbugwe, and et al. 2022. "Nutritional Quality of Four Novel Porridge Products Blended with Edible Cricket (Scapsipedus icipe) Meal for Food" Foods 11, no. 7: 1047. https://doi.org/10.3390/foods11071047