
Dietary Postbiotics Reduce Cytotoxicity and Inflammation Induced by Crystalline Silica in an In Vitro RAW 264.7 Macrophage Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
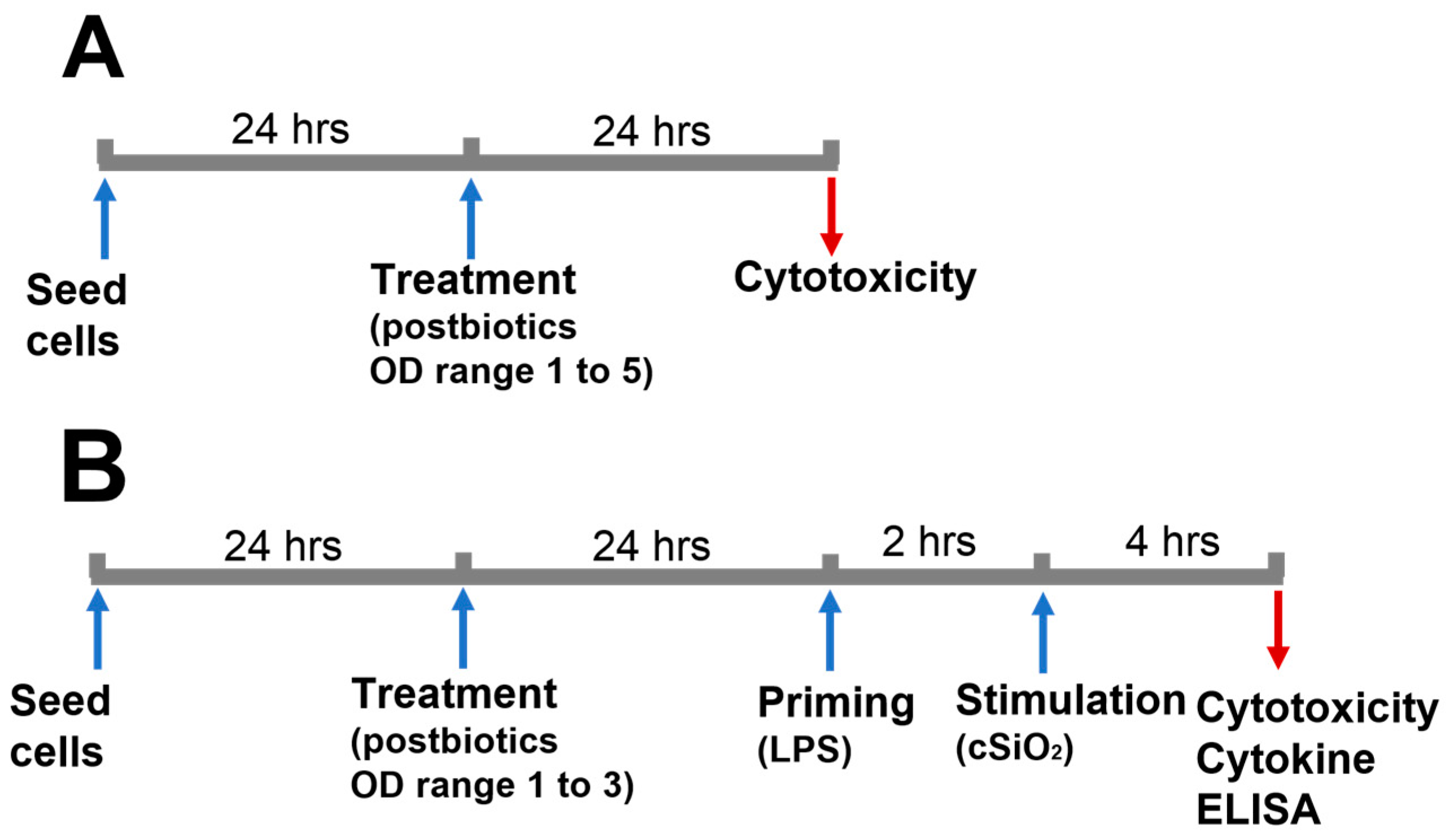
2.1. Experimental Design
2.2. Bacterial Strains and Preparation of Postbiotics
2.3. Cell Lines and Culture Conditions
2.4. cSiO2 Preparation
2.5. Cytotoxicity
2.6. IL-1 Cytokine Analyses
2.7. Statistical Analyses
3. Results
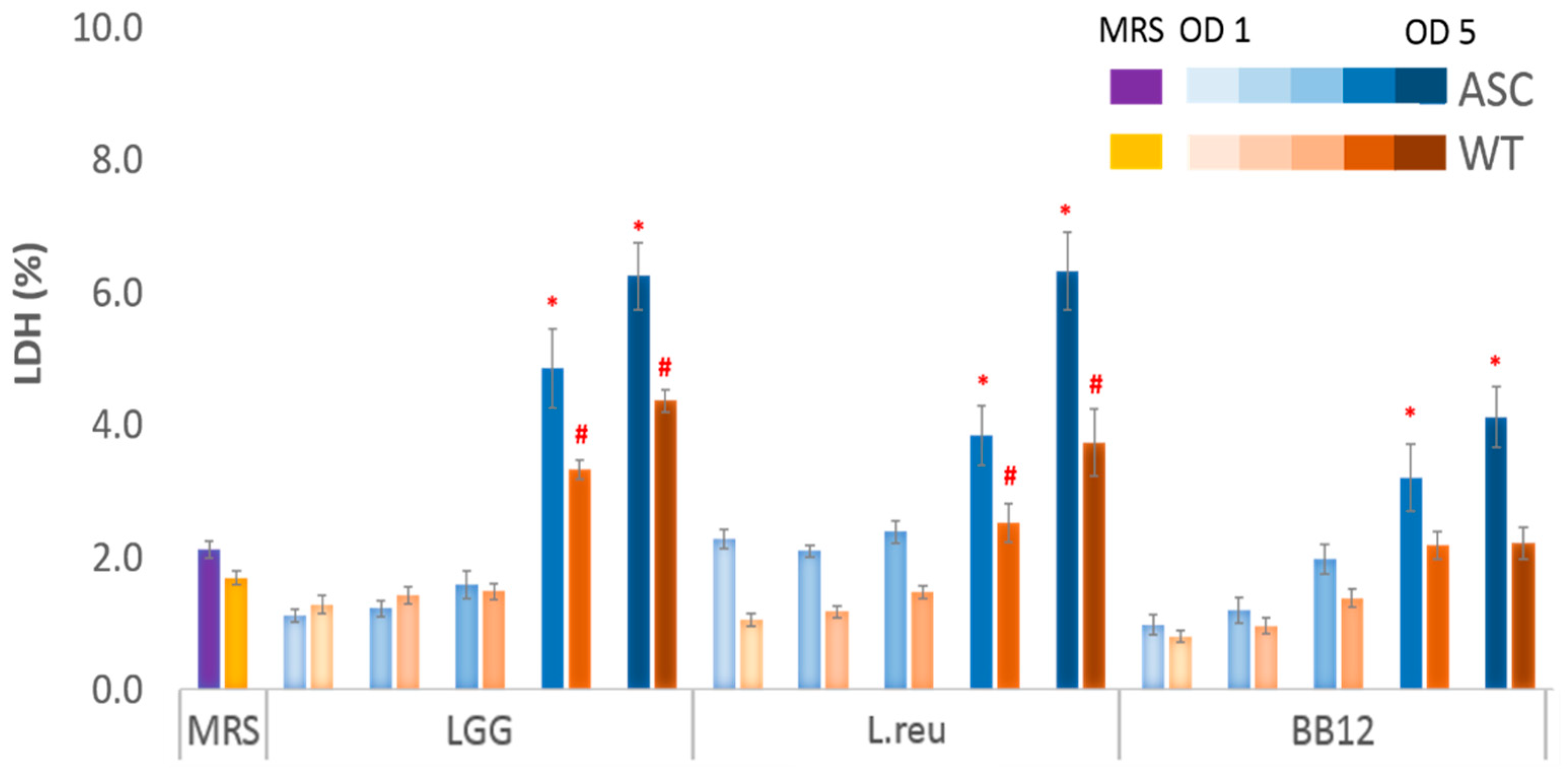
3.1. Effect of Postbiotics on Macrophage Survivability
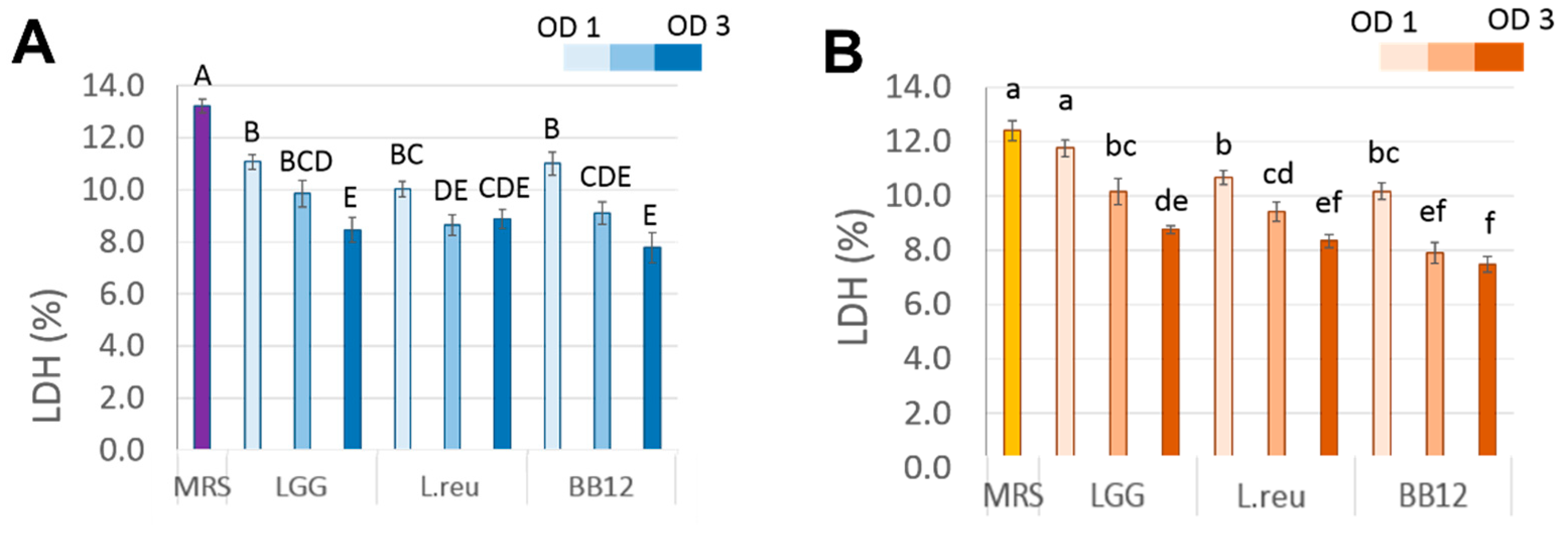
3.2. Effect of Postbiotics on cSiO2-Induced Cytotoxicity in Macrophages
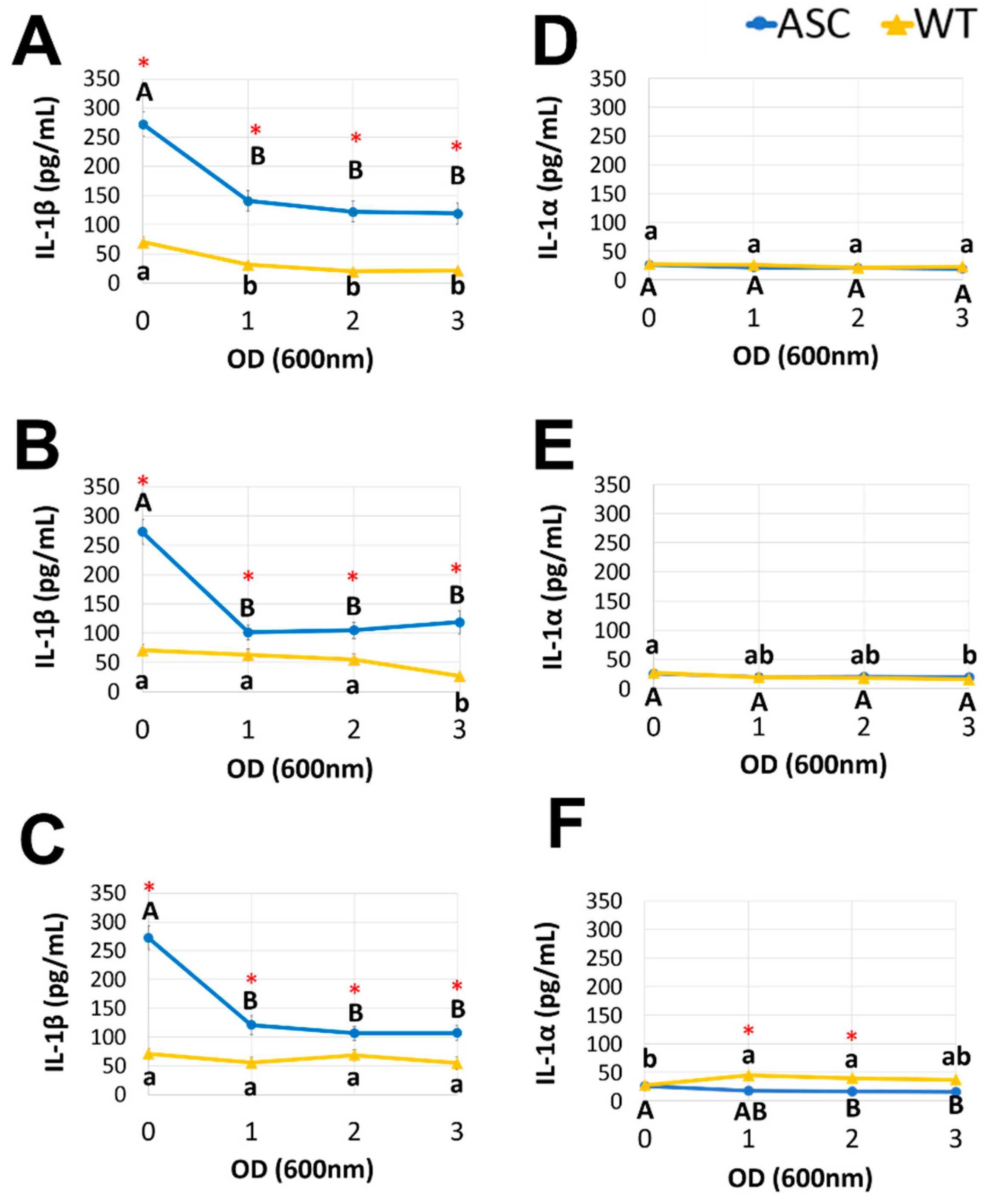
3.3. Postbiotics Suppressed Proinflammatory Cytokine Release
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Tsilingiri, K.; Rescigno, M. Postbiotics: What else? Benef. Microbes 2013, 4, 101–107. [Google Scholar] [CrossRef]
- Aguilar-Toala, J.E.; Arioli, S.; Behare, P.; Belzer, C.; Berni Canani, R.; Chatel, J.M.; D’Auria, E.; de Freitas, M.Q.; Elinav, E.; Esmerino, E.A.; et al. Postbiotics—When simplification fails to clarify. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 825–826. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Mayorgas, A.; Dotti, I.; Salas, A. Microbial Metabolites, Postbiotics, and Intestinal Epithelial Function. Mol. Nutr. Food Res. 2021, 65, e2000188. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Shi, B. Gut microbiota as a potential target of metabolic syndrome: The role of probiotics and prebiotics. Cell Biosci. 2017, 7, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.J.; Guo, J.; Jia, Q.; Huang, Y.S.; Huang, W.J.; Zhang, W.; Zhang, F.; Liu, W.J.; Wang, Y. The effect of probiotic and synbiotic supplementation on biomarkers of inflammation and oxidative stress in diabetic patients: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2019, 142, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Kesika, P.; Suganthy, N.; Sivamaruthi, B.S.; Chaiyasut, C. Role of gut-brain axis, gut microbial composition, and probiotic intervention in Alzheimer’s disease. Life Sci. 2021, 264, 118627. [Google Scholar] [CrossRef]
- Manirarora, J.N.; Kosiewicz, M.M.; Alard, P. Feeding lactobacilli impacts lupus progression in (NZBxNZW)F1 lupus-prone mice by enhancing immunoregulation. Autoimmunity 2020, 53, 323–332. [Google Scholar] [CrossRef]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermudez-Humaran, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [Green Version]
- Morniroli, D.; Vizzari, G.; Consales, A.; Mosca, F.; Gianni, M.L. Postbiotic Supplementation for Children and Newborn’s Health. Nutrients 2021, 13, 781. [Google Scholar] [CrossRef] [PubMed]
- Mosca, F.; Gianni, M.L.; Rescigno, M. Can Postbiotics Represent a New Strategy for NEC? In Probiotics and Child Gastrointestinal Health: Advances in Microbiology, Infectious Diseases and Public Health, Volume 10; Guandalini, S., Indrio, F., Eds.; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2019; Volume 1125, pp. 37–45. [Google Scholar]
- Tsilingiri, K.; Barbosa, T.; Penna, G.; Caprioli, F.; Sonzogni, A.; Viale, G.; Rescigno, M. Probiotic and postbiotic activity in health and disease: Comparison on a novel polarised ex-vivo organ culture model. Gut 2012, 61, 1007–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolkiewicz, J.; Marzec, A.; Ruszczynski, M.; Feleszko, W. Postbiotics-A Step Beyond Pre- and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef] [PubMed]
- Shenderov, B.A. Metabiotics: Novel idea or natural development of probiotic conception. Microb. Ecol. Health Dis. 2013, 24, 20399. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lu, B.; Fu, J.; Zhu, X.; Song, E.; Song, Y. Amorphous silica nanoparticles induce inflammation via activation of NLRP3 inflammasome and HMGB1/TLR4/MYD88/NF-kb signaling pathway in HUVEC cells. J. Hazard. Mater. 2021, 404, 124050. [Google Scholar] [CrossRef] [PubMed]
- National Institude for Occupational Safety and Health. Health Effects of Occupational Exposure to Respirable Crystalline Silica; National Institude for Occupational Safety and Health: Cincinnati, OH, USA, 2002.
- Greenberg, M.I.; Waksman, J.; Curtis, J. Silicosis: A review. Dis Mon. 2007, 53, 394–416. [Google Scholar] [CrossRef] [PubMed]
- Bates, M.A.; Brandenberger, C.; Langohr, I.I.; Kumagai, K.; Lock, A.L.; Harkema, J.R.; Holian, A.; Pestka, J.J. Silica-Triggered Autoimmunity in Lupus-Prone Mice Blocked by Docosahexaenoic Acid Consumption. PLoS ONE 2016, 11, e0160622. [Google Scholar] [CrossRef]
- Cooper, G.S.; Miller, F.W.; Germolec, D.R. Occupational exposures and autoimmune diseases. Int. Immunopharmacol. 2002, 2, 303–313. [Google Scholar] [CrossRef]
- Finckh, A.; Cooper, G.S.; Chibnik, L.B.; Costenbader, K.H.; Watts, J.; Pankey, H.; Fraser, P.A.; Karlson, E.W. Occupational silica and solvent exposures and risk of systemic lupus erythematosus in urban women. Arthritis Rheum. 2006, 54, 3648–3654. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Brito, M.; Munoz-Quezada, S.; Gomez-Llorente, C.; Matencio, E.; Bernal, M.J.; Romero, F.; Gil, A. Human intestinal dendritic cells decrease cytokine release against Salmonella infection in the presence of Lactobacillus paracasei upon TLR activation. PLoS ONE 2012, 7, e43197. [Google Scholar] [CrossRef] [Green Version]
- Frick, J.S.; Schenk, K.; Quitadamo, M.; Kahl, F.; Koberle, M.; Bohn, E.; Aepfelbacher, M.; Autenrieth, I.B. Lactobacillus fermentum attenuates the proinflammatory effect of Yersinia enterocolitica on human epithelial cells. Inflamm. Bowel. Dis. 2007, 13, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Wierenga, K.A.; Wee, J.; Gilley, K.N.; Rajasinghe, L.D.; Bates, M.A.; Gavrilin, M.A.; Holian, A.; Pestka, J.J. Docosahexaenoic Acid Suppresses Silica-Induced Inflammasome Activation and IL-1 Cytokine Release by Interfering With Priming Signal. Front. Immunol. 2019, 10, 2130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.S.; Liao, X.F.; Sparks, J.B.; Luo, X.M. Dynamics of Gut Microbiota in Autoimmune Lupus. Appl. Environ. Microbiol. 2014, 80, 7551–7560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antypas, H.; Libberton, B.; Melican, K. Reducing background cytokine expression in epithelial cells without serum starvation. MethodsX 2014, 1, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Gutting, B.W.; Gaske, K.S.; Schilling, A.S.; Slaterbeck, A.F.; Sobota, L.; Mackie, R.S.; Buhr, T.L. Differential susceptibility of macrophage cell lines to Bacillus anthracis-Vollum 1B. Toxicol. Vitr. 2005, 19, 221–229. [Google Scholar] [CrossRef]
- Di Nunzio, M.; Valli, V.; Tomas-Cobos, L.; Tomas-Chisbert, T.; Murgui-Bosch, L.; Danesi, F.; Bordoni, A. Is cytotoxicity a determinant of the different in vitro and in vivo effects of bioactives? BMC Complement Altern. Med. 2017, 17, 453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurazawa, T.; Ohkusa, T. Cytotoxicity of organic acids produced by anaerobic intestinal bacteria on cultured epithelial cells. J. Gastroenterol. 2005, 40, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Martinet, W.; Coornaert, I.; Puylaert, P.; De Meyer, G.R.Y. Macrophage Death as a Pharmacological Target in Atherosclerosis. Front. Pharmacol. 2019, 10, 306. [Google Scholar] [CrossRef] [PubMed]
- Cicenia, A.; Santangelo, F.; Gambardella, L.; Pallotta, L.; Iebba, V.; Scirocco, A.; Marignani, M.; Tellan, G.; Carabotti, M.; Corazziari, E.S.; et al. Protective Role of Postbiotic Mediators Secreted by Lactobacillus rhamnosus GG Versus Lipopolysaccharide-induced Damage in Human Colonic Smooth Muscle Cells. J. Clin. Gastroenterol. 2016, 50 (Suppl. 2), S140–S144. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Hara, H.; Nunez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef] [Green Version]
- Gross, O.; Yazdi, A.S.; Thomas, C.J.; Masin, M.; Heinz, L.X.; Guarda, G.; Quadroni, M.; Drexler, S.K.; Tschopp, J. Inflammasome activators induce interleukin-1alpha secretion via distinct pathways with differential requirement for the protease function of caspase-1. Immunity 2012, 36, 388–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Li, J.; Liu, Y.; Xiao, N.; Suo, H.; Xie, K.; Yang, C.; Wu, C. Short-chain fatty acids suppress lipopolysaccharide-induced production of nitric oxide and proinflammatory cytokines through inhibition of NF-kappaB pathway in RAW264.7 cells. Inflammation 2012, 35, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, J.; Weng, T.; Shen, K.; Chen, Z.; Yu, Y.; Huang, Q.; Wang, G.; Liu, Z.; Jin, S. The Inflammation Induced by Lipopolysaccharide can be Mitigated by Short-chain Fatty Acid, Butyrate, through Upregulation of IL-10 in Septic Shock. Scand. J. Immunol. 2017, 85, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Toala, J.E.; Hall, F.G.; Urbizo-Reyes, U.C.; Garcia, H.S.; Vallejo-Cordoba, B.; Gonzalez-Cordova, A.F.; Hernandez-Mendoza, A.; Liceaga, A.M. In Silico Prediction and In Vitro Assessment of Multifunctional Properties of Postbiotics Obtained From Two Probiotic Bacteria. Probiotics Antimicrob. Proteins 2020, 12, 608–622. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, X.; Rodriguez, J.; Wee, J. Dietary Postbiotics Reduce Cytotoxicity and Inflammation Induced by Crystalline Silica in an In Vitro RAW 264.7 Macrophage Model. Foods 2022, 11, 877. https://doi.org/10.3390/foods11060877
Du X, Rodriguez J, Wee J. Dietary Postbiotics Reduce Cytotoxicity and Inflammation Induced by Crystalline Silica in an In Vitro RAW 264.7 Macrophage Model. Foods. 2022; 11(6):877. https://doi.org/10.3390/foods11060877
Chicago/Turabian StyleDu, Xue, Jessica Rodriguez, and Josephine Wee. 2022. "Dietary Postbiotics Reduce Cytotoxicity and Inflammation Induced by Crystalline Silica in an In Vitro RAW 264.7 Macrophage Model" Foods 11, no. 6: 877. https://doi.org/10.3390/foods11060877