
Inhibition of Listeria monocytogenes by Phage Lytic Enzymes Displayed on Tailored Bionanoparticles


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids and Culture Conditions
2.2. Bioinformatic Analysis of Phage vB_LmoS_293
2.3. Plasmid Construction for PHA BNP Generation
2.4. Generation of PHA-BNPs
2.5. Lysis and Growth Inhibition Assays
2.5.1. Preparation of Bacterial Culture and Protein
2.5.2. Application of PHA_BNPs for Lysis of L. monocytogenes
2.5.3. Application of PHA_BNPs for Growth Inhibition of L. monocytogenes
2.6. Statistical Analysis
3. Results
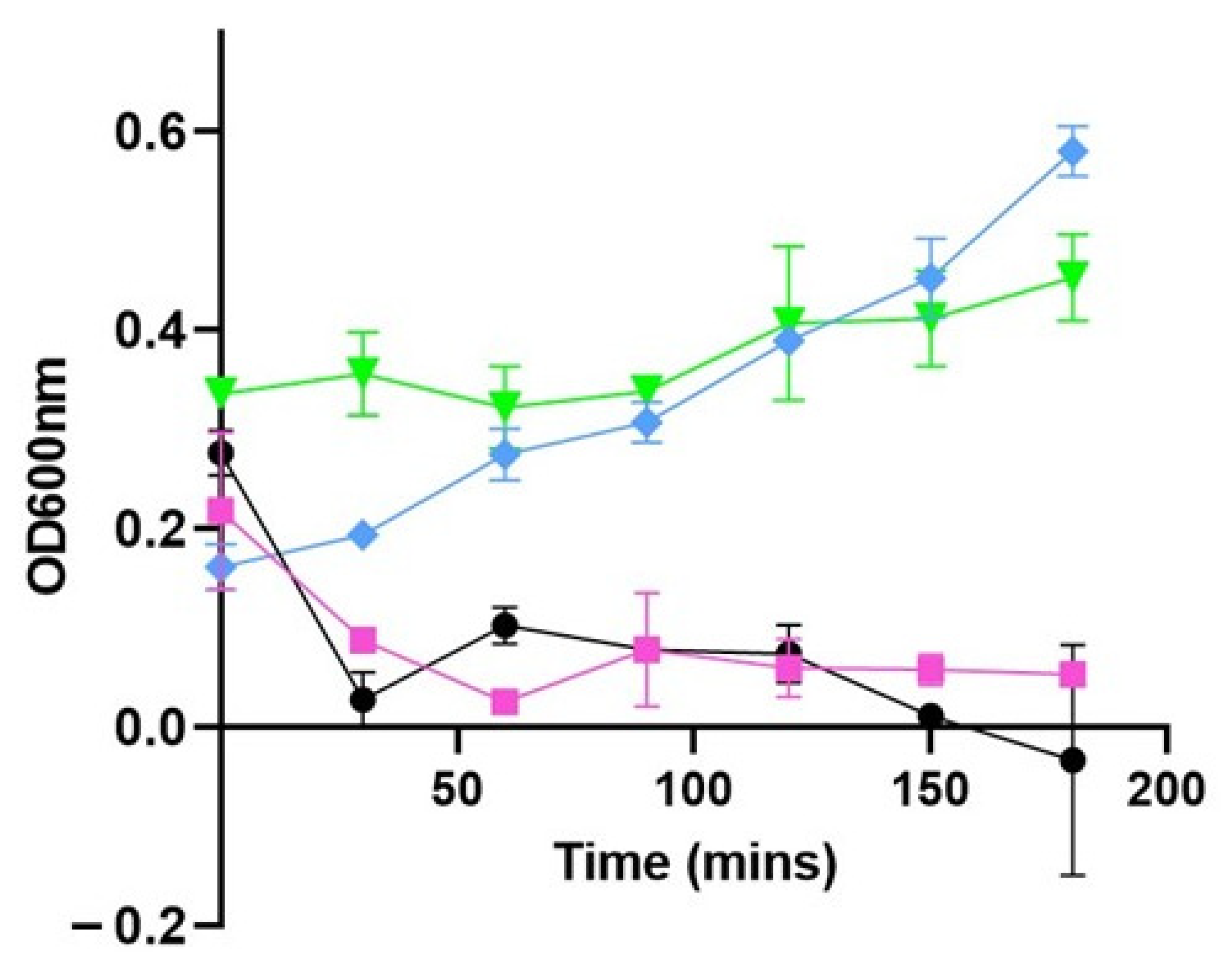
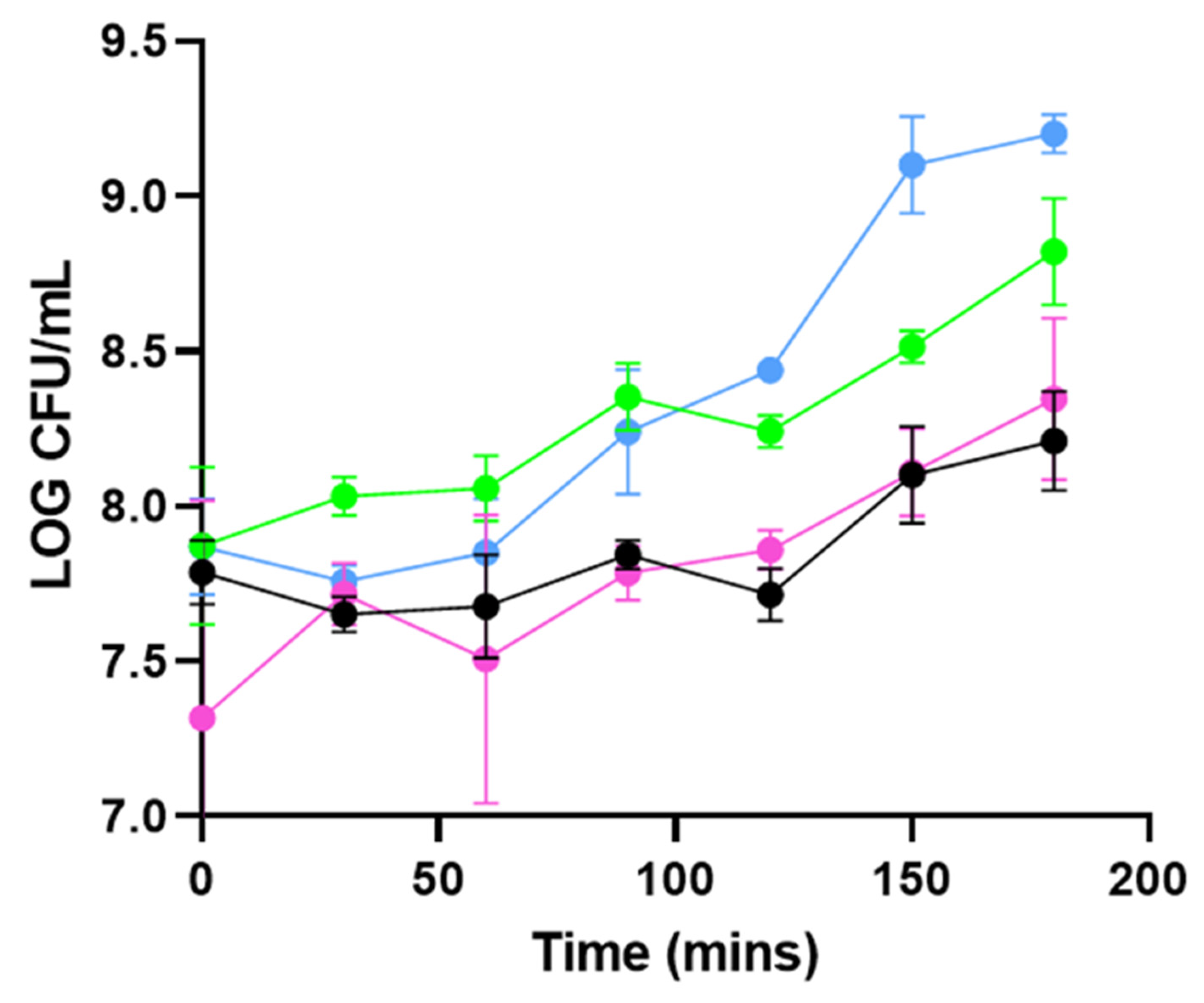
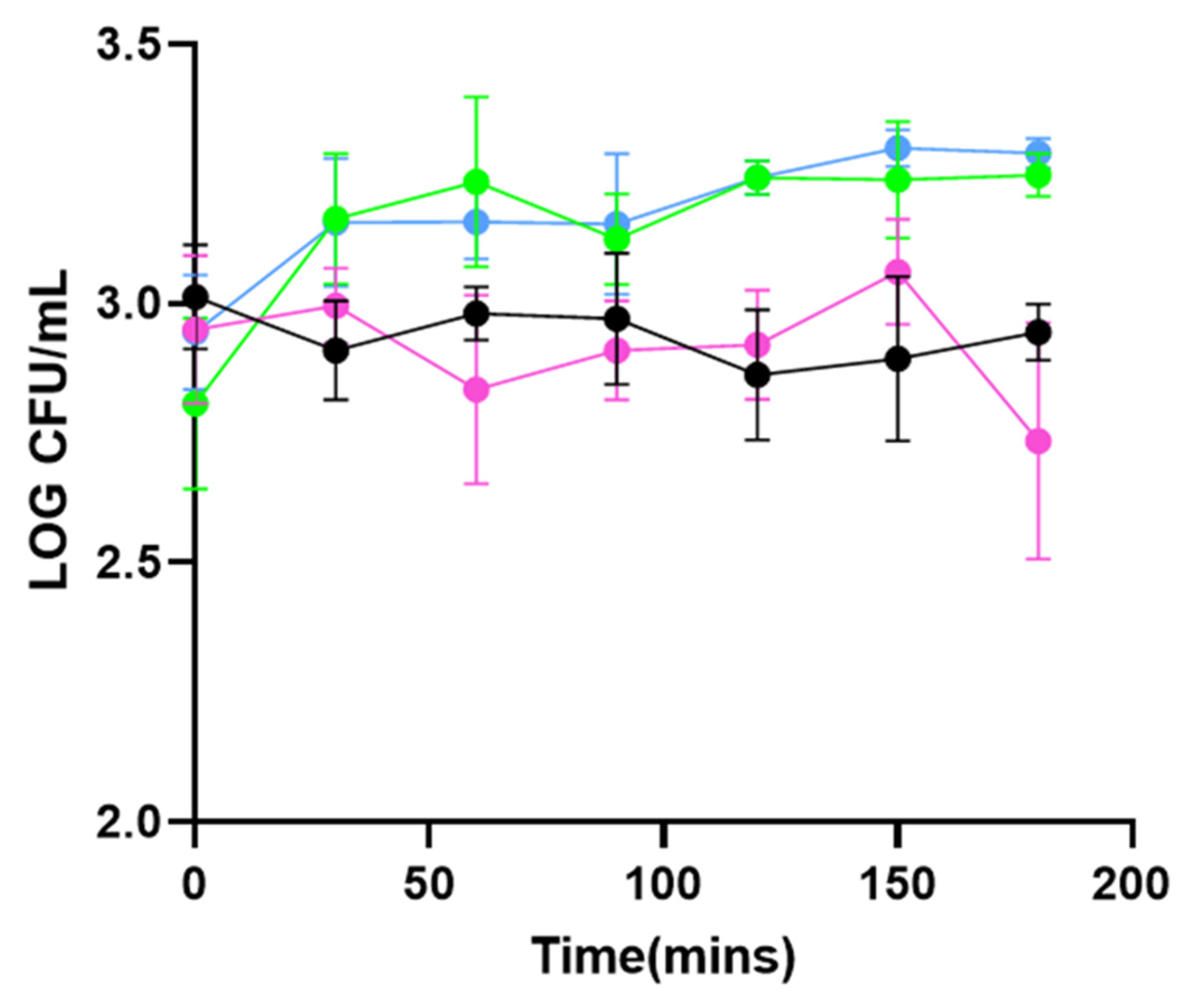
3.1. PHA_BNPs Displaying Lysin293 and Amidase293 Cause Lysis of L. monocytogenes
3.2. PHA_BNPs Displaying Lysin293 and Amidase293 Cause Growth Inhibition of L. monocytogenes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, Q.; Gooneratne, R.; Hussain, M.A. Listeria monocytogenes in Fresh Produce: Outbreaks, Prevalence and Contamination Levels. Foods 2017, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Desai, A.N.; Anyoha, A.; Madoff, L.C.; Lassmann, B. Changing epidemiology of Listeria monocytogenes outbreaks, sporadic cases, and recalls globally: A review of ProMED reports from 1996 to 2018. Int. J. Infect. Dis. 2019, 84, 48–53. [Google Scholar] [CrossRef] [Green Version]
- de Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef] [Green Version]
- Martins, E.A.; Germano, P.M.L. Listeria monocytogenes in ready-to-eat, sliced, cooked ham and salami products, marketed in the city of São Paulo, Brazil: Occurrence, quantification, and serotyping. Food Control 2011, 22, 297–302. [Google Scholar] [CrossRef]
- Oliveira, M.; Viñas, I.; Colàs, P.; Anguera, M.; Usall, J.; Abadias, M. Effectiveness of a bacteriophage in reducing Listeria monocytogenes on fresh-cut fruits and fruit juices. Food Microbiol. 2014, 38, 137–142. [Google Scholar] [CrossRef]
- Stone, E.; Lhomet, A.; Neve, H.; Grant, I.; Campbell, K.; McAuliffe, O. Isolation and Characterization of Listeria monocytogenes phage vB_LmoH_P61, a phage with biocontrol potential on different food matrices. Front. Sustain. Food Syst. 2020, 4, 521645. [Google Scholar] [CrossRef]
- Oechslin, F.; Menzi, C.; Moreillon, P.; Resch, G. The multi-domain architecture of a bacteriophage endolysin enables intramolecular synergism and regulation of bacterial lysis. J. Biol. Chem. 2021, 296, 100639. [Google Scholar] [CrossRef]
- Borysowski, J.; Weber-Dąbrowska, B.; Górski, A. Bacteriophage endolysins as a novel class of antibacterial agents. Exp. Biol. Med. 2006, 231, 366–377. [Google Scholar] [CrossRef]
- Haddad Kashani, H.; Schmelcher, M.; Sabzalipoor, H.; Seyed Hosseini, E.; Moniri, R. Recombinant endolysins as potential therapeutics against antibiotic-resistant Staphylococcus aureus: Current status of research and novel delivery strategies. Clin. Microbiol. Rev. 2018, 31, e00071-17. [Google Scholar] [CrossRef] [Green Version]
- Radford, D.; Guild, B.; Strange, P.; Ahmed, R.; Lim, L.-T.; Balamurugan, S. Characterization of antimicrobial properties of Salmonella phage Felix O1 and Listeria phage A511 embedded in xanthan coatings on poly(lactic acid) films. Food Microbiol. 2017, 66, 117–128. [Google Scholar] [CrossRef]
- Schmelcher, M.; Loessner, M.J. Bacteriophage endolysins: Applications for food safety. Curr. Opin. Biotechnol. 2016, 37, 76–87. [Google Scholar] [CrossRef]
- Shannon, R.; Radford, D.R.; Balamurugan, S. Impacts of food matrix on bacteriophage and endolysin antimicrobial efficacy and performance. Crit. Rev. Food Sci. Nutr. 2020, 60, 1631–1640. [Google Scholar] [CrossRef]
- Pennone, V.; Sanz-Gaitero, M.; O’Connor, P.; Coffey, A.; Jordan, K.; van Raaij, M.J.; McAuliffe, O. Inhibition of L. monocytogenes Biofilm Formation by the Amidase Domain of the Phage vB_LmoS_293 Endolysin. Viruses 2019, 11, 722. [Google Scholar] [CrossRef] [Green Version]
- Altermann, E.; Schofield, L.R.; Ronimus, R.S.; Beatty, A.K.; Reilly, K. Inhibition of Rumen Methanogens by a Novel Archaeal Lytic Enzyme Displayed on Tailored Bionanoparticles. Front. Microbiol. 2018, 9, 2378. [Google Scholar] [CrossRef] [Green Version]
- Pakalapati, H.; Chang, C.K.; Show, P.L.; Arumugasamy, S.K.; Lan, J.C. Development of polyhydroxyalkanoates production from waste feedstocks and applications. J. Biosci. Bioeng. 2018, 126, 282–292. [Google Scholar] [CrossRef]
- Raza, Z.A.; Abid, S.; Banat, I.M. Polyhydroxyalkanoates: Characteristics, production, recent developments and applications. Int. Biodeterior. Biodegrad. 2018, 126, 45–56. [Google Scholar] [CrossRef]
- Davies, C.G.; Reilly, K.; Altermann, E.; Hendrickson, H.L. PLAN-M; Mycobacteriophage Endolysins Fused to Biodegradable Nanobeads Mitigate Mycobacterial Growth in Liquid and on Surfaces. Front. Microbiol. 2021, 12, 562748. [Google Scholar] [CrossRef]
- Amara, A.A.; Rehm, B.H. Replacement of the catalytic nucleophile cysteine-296 by serine in class II polyhydroxyalkanoate synthase from Pseudomonas aeruginosa-mediated synthesis of a new polyester: Identification of catalytic residues. Biochem. J. 2003, 374, 413–421. [Google Scholar] [CrossRef]
- Casey, A.; Jordan, K.; Coffey, A.; McAuliffe, O. Complete Genome Sequences of vB_LmoS_188 and vB_LmoS_293, Two Bacteriophages with Specificity for Listeria monocytogenes Strains of Serotypes 4b and 4e. Genome Announc. 2015, 3, e00040-15. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Panchenko, A.R.; Shoemaker, B.A.; Thiessen, P.A.; Geer, L.Y.; Bryant, S.H. CDD: A database of conserved domain alignments with links to domain three-dimensional structure. Nucleic Acids Res. 2002, 30, 281–283. [Google Scholar] [CrossRef]
- González-Miró, M.; Rodríguez-Noda, L.M.; Fariñas-Medina, M.; Cedré-Marrero, B.; Madariaga-Zarza, S.; Zayas-Vignier, C.; Hernández-Cedeño, M.; Kleffmann, T.; García-Rivera, D.; Vérez-Bencomo, V.; et al. Bioengineered polyester beads co-displaying protein and carbohydrate-based antigens induce protective immunity against bacterial infection. Sci Rep. 2018, 8, 1888. [Google Scholar] [CrossRef] [Green Version]
- Fenton, M.; Keary, R.; McAuliffe, O.; Ross, R.P.; O’Mahony, J.; Coffey, A. Bacteriophage-Derived Peptidase CHAP(K) Eliminates and Prevents Staphylococcal Biofilms. Int. J. Microbiol. 2013, 2013, 625341. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.J.; Garefalaki, V.; Spoerl, R.; Narbad, A.; Meijers, R. Structure-based modification of a Clostridium difficile-targeting endolysin affects activity and host range. J. Bacteriol. 2011, 193, 5477–5486. [Google Scholar] [CrossRef] [Green Version]
- Casey Jordan, K.; Neve, H.; Coffey, A.; McAuliffe, O. A tail of two phages: Genomic and functional analysis of Listeria monocytogenes phages vB_LmoS_188 and vB_LmoS_293 reveal the receptor-binding proteins involved in host specificity. Front. Microbiol. 2015, 6, 1107. [Google Scholar] [CrossRef] [Green Version]
- Horgan, M.; O’Flynn, G.; Garry, J.; Cooney, J.; Coffey, A.; Fitzgerald, G.F.; Ross, R.P.; McAuliffe, O. Phage lysin LysK can be truncated to its CHAP domain and retain lytic activity against live antibiotic-resistant staphylococci. Appl. Environ. Microbiol. 2009, 75, 872–874. [Google Scholar] [CrossRef] [Green Version]
- Pennone, V. The Occurrence of Listeria Monocytogenes in the Mushroom Production Chain and the Use of Bacteriophage for Its Control. Ph.D. Thesis, Munster Technological University, Cork, Ireland, March 2019. [Google Scholar]
- Solanki, K.; Grover, N.; Downs, P.; Paskaleva, E.E.; Mehta, K.K.; Lee, L.; Schadler, L.S.; Kane, R.S.; Dordick, J.S. Enzyme-based listericidal nanocomposites. Sci. Rep. 2013, 3, 1584. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Plasmid Name | Insert | Resistance | Host Bacterium | Product | Reference |
|---|---|---|---|---|---|
| pET14b-PHA_lysin293_BNPs | Gene fusion of lysin293 and PhaC | AmpR | E. coli BL21 (DE3) | PHA_lysin293_BNPs | This study |
| pET14b-PHA_amidase293_BNPs | Gene fusion of amidase293 and PhaC | AmpR | E. coli BL21 (DE3) | PHA_amidase293_BNPs | This study |
| pET14b-PHA_BNPs | PhaC sequence | AmpR | E. coli BL21 (DE3) | PHA_BNPs | This study |
| pMCS69 (helper plasmid) | N/A | CmR | E. coli BL21 (DE3) | N/A | [18] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stone, E.; Pennone, V.; Reilly, K.; Grant, I.R.; Campbell, K.; Altermann, E.; McAuliffe, O. Inhibition of Listeria monocytogenes by Phage Lytic Enzymes Displayed on Tailored Bionanoparticles. Foods 2022, 11, 854. https://doi.org/10.3390/foods11060854
Stone E, Pennone V, Reilly K, Grant IR, Campbell K, Altermann E, McAuliffe O. Inhibition of Listeria monocytogenes by Phage Lytic Enzymes Displayed on Tailored Bionanoparticles. Foods. 2022; 11(6):854. https://doi.org/10.3390/foods11060854
Chicago/Turabian StyleStone, Edel, Vincenzo Pennone, Kerri Reilly, Irene R. Grant, Katrina Campbell, Eric Altermann, and Olivia McAuliffe. 2022. "Inhibition of Listeria monocytogenes by Phage Lytic Enzymes Displayed on Tailored Bionanoparticles" Foods 11, no. 6: 854. https://doi.org/10.3390/foods11060854