
Curcumin-Mediated Sono-Photodynamic Treatment Inactivates Listeria monocytogenes via ROS-Induced Physical Disruption and Oxidative Damage

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Preparation of Bacterial Suspension and Curcumin Solution
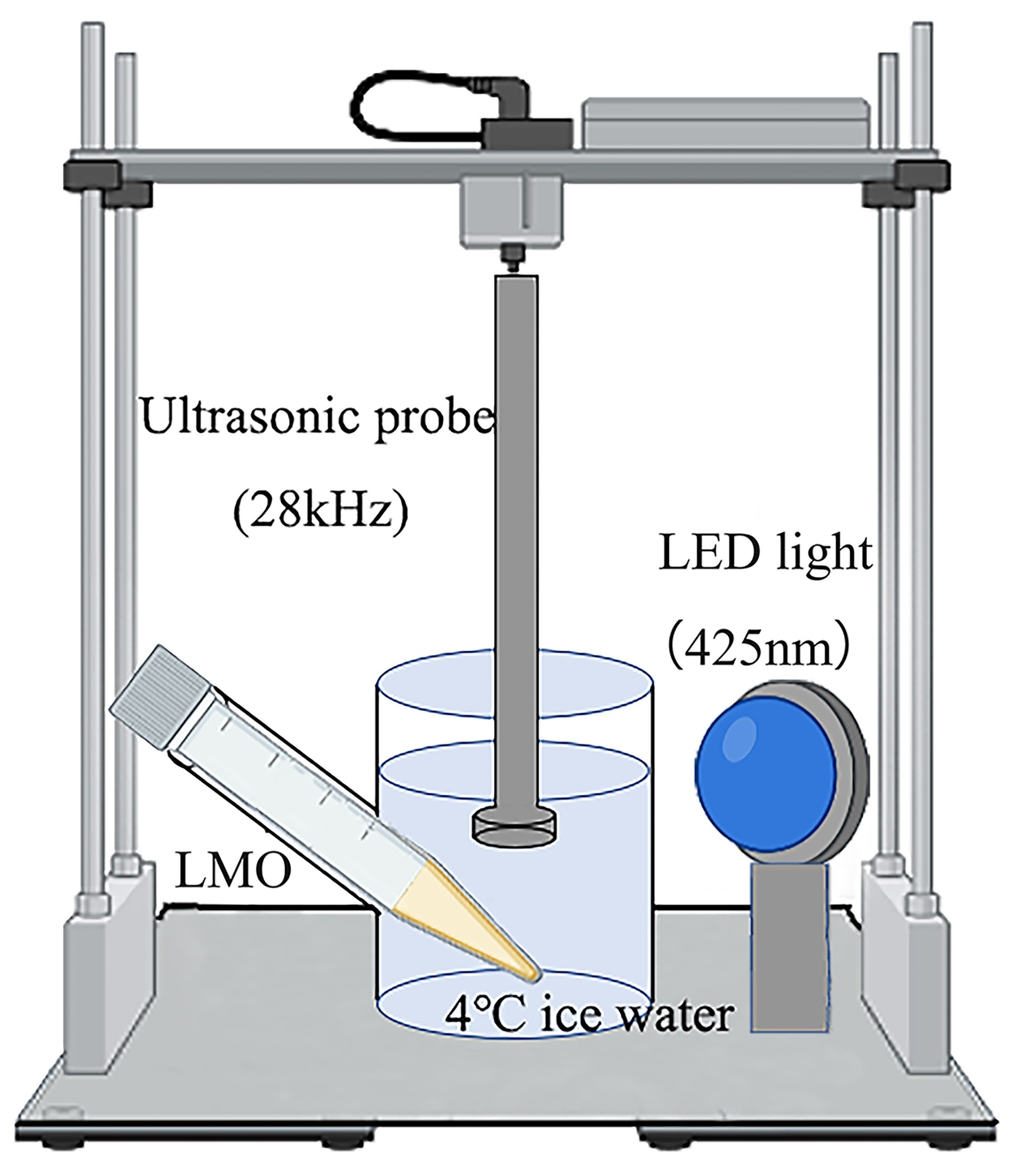
2.2. Sono-Photodynamic Sterilization Treatment
2.3. Detection of Reactive Oxygen Species (ROS)
2.4. Scanning Electron Microscopy (SEM)
2.5. Determination of Cytoplasmic Material Leakage
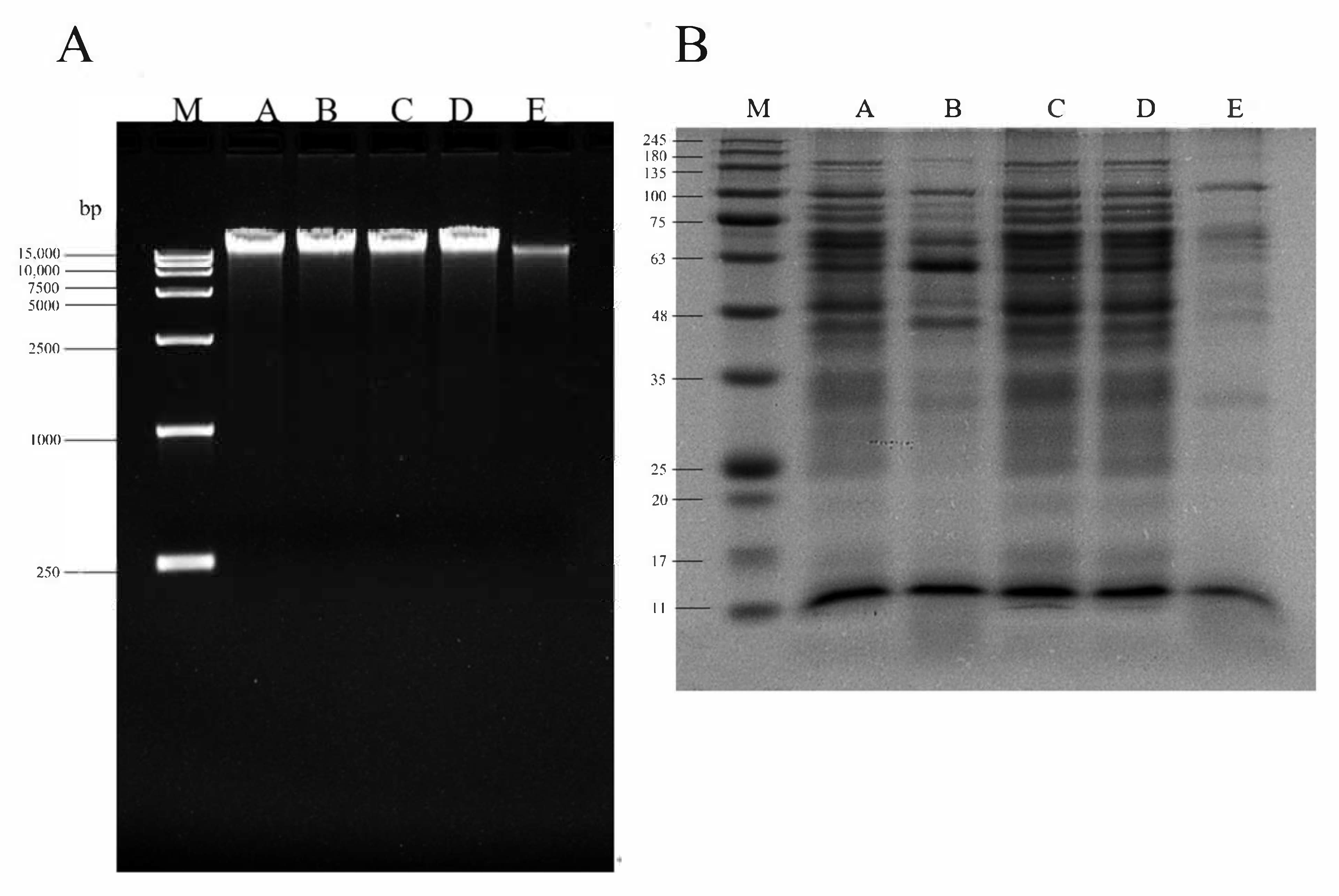
2.6. DNA Agarose Gel Electrophoresis
2.7. Polyacrylamide Gel Electrophoresis (SDS-PAGE)
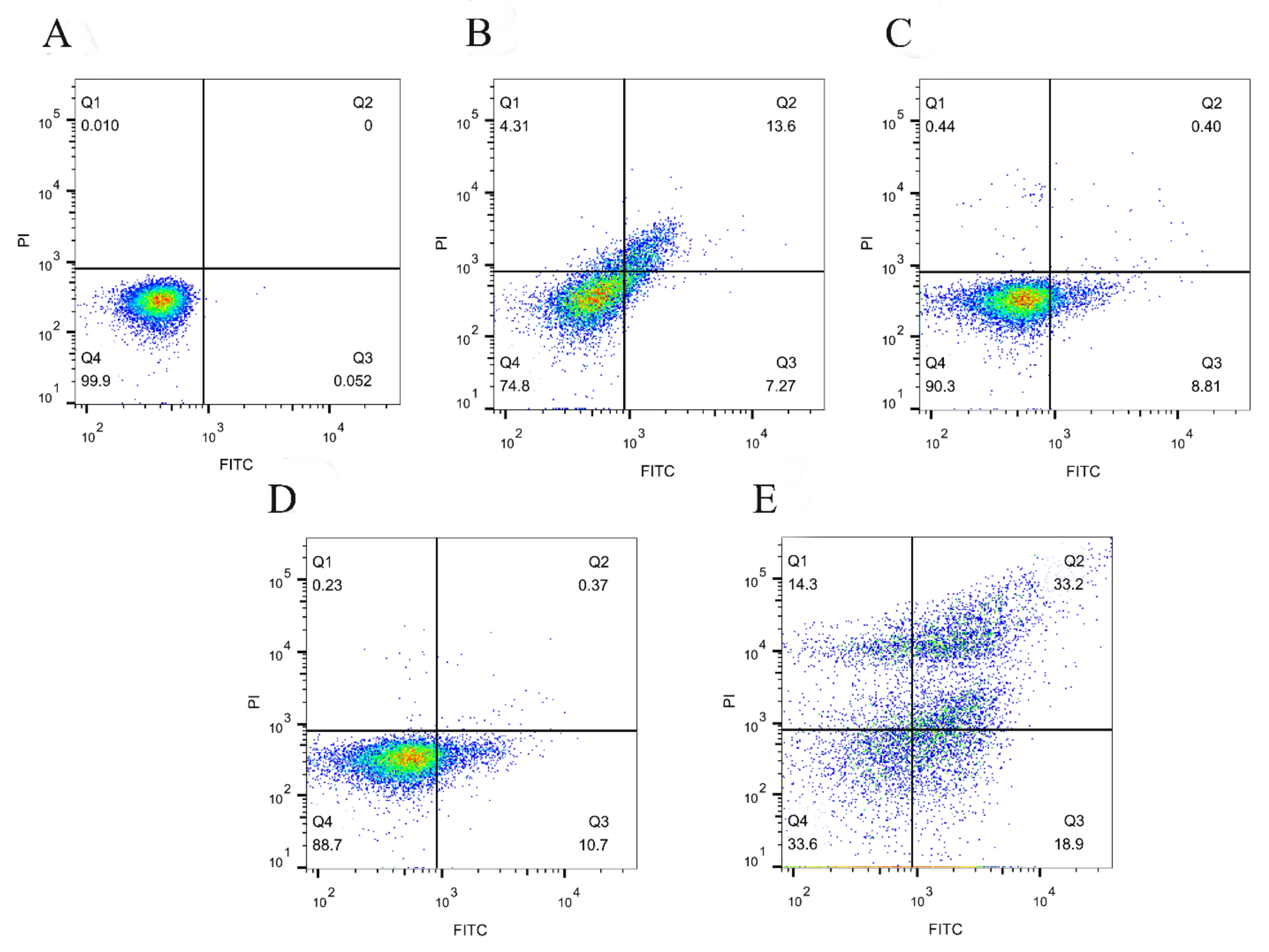
2.8. Annexin V-FITC/PI Staining Assay
2.9. Data Statistics and Analysis
3. Results
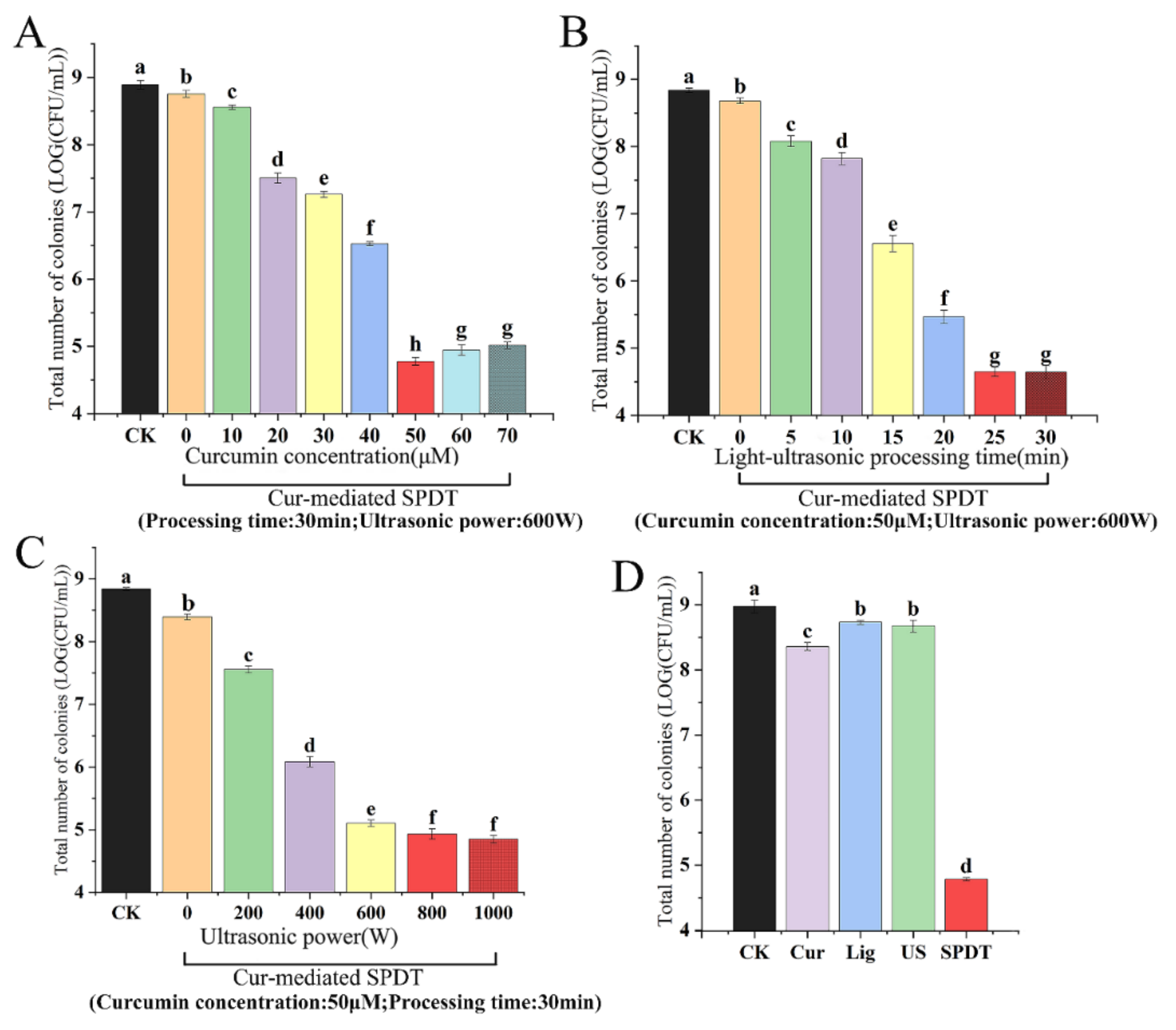
3.1. SPDT Showed Effective Bactericidal Activity against LMO
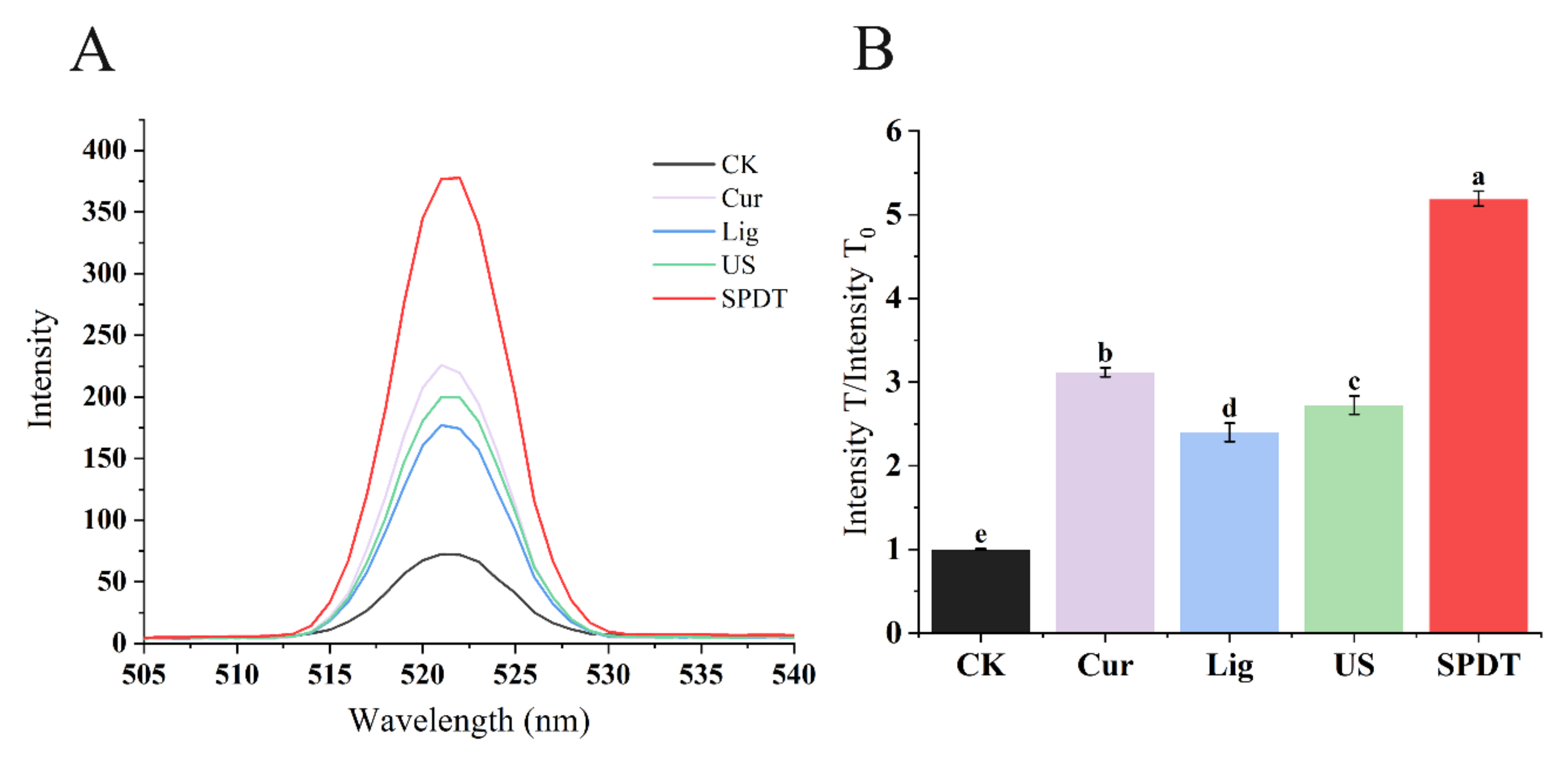
3.2. SPDT Generated Significant Intracellular ROS in LMO
3.3. SPDT Altered Morphology of LMO
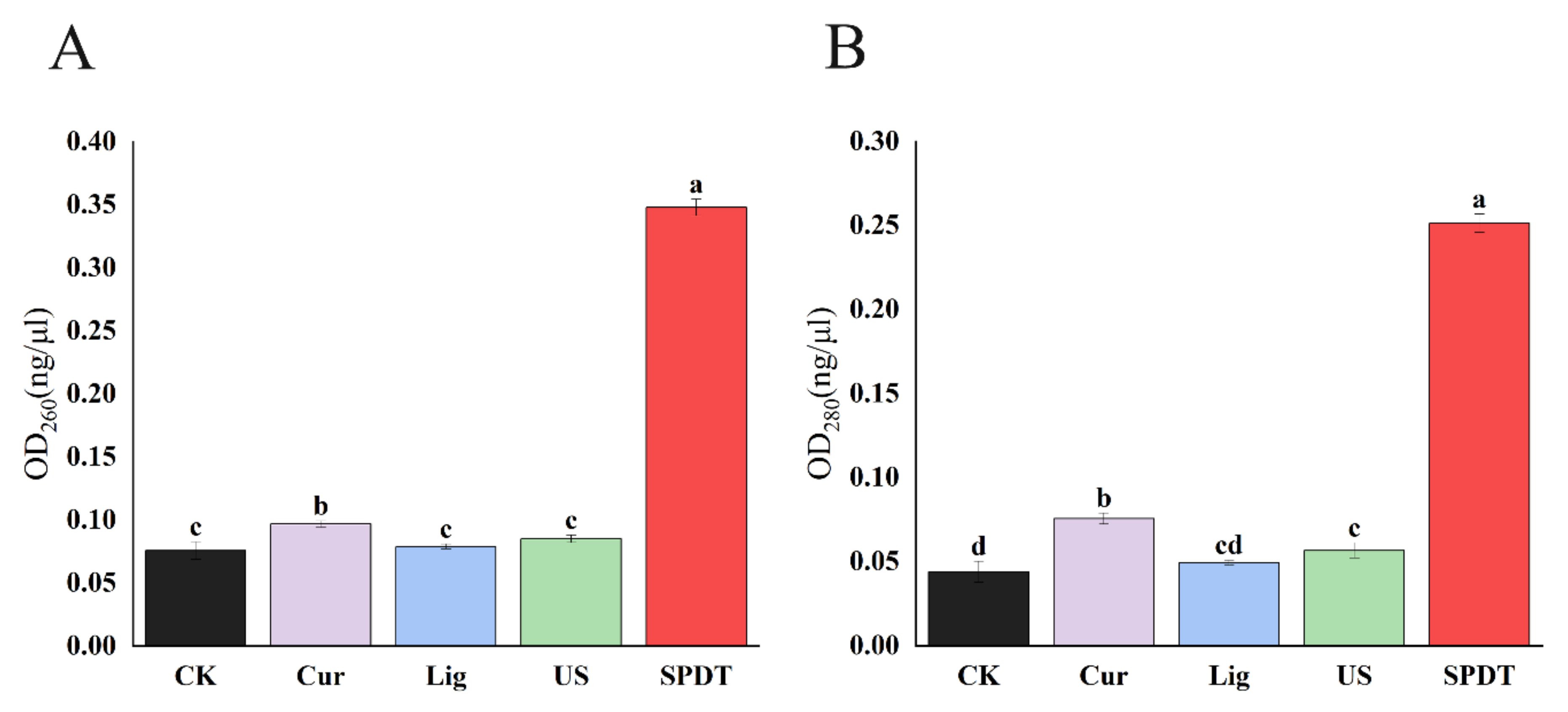
3.4. SPDT Induced Cytoplasmic Material Leakage in LMO
3.5. SPDT Induced DNA Fragmentation and Protein Degradation in LMO
3.6. SPDT Induced Membrane Depolarization in LMO
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pires, S.M.; Desta, B.N.; Mughini-Gras, L.; Mmbaga, B.T.; Fayemi, O.E.; Salvador, E.M.; Gobena, T.; Majowicz, S.E.; Hald, T.; Hoejskov, P.S.; et al. Burden of foodborne diseases: Think global, act local. Curr. Opin. Food Sci. 2021, 39, 152–159. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Desvaux, M.; Hébraud, M.; Møretrø, T.; Langsrud, S.; Doulgeraki, A.; Nychas, G.J.; Kačániová, M.; Czaczyk, K.; et al. Intra- and inter-species interactions within biofilms of important foodborne bacterial pathogens. Front. Microbiol. 2015, 6, 841. [Google Scholar] [CrossRef]
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on Major Food-Borne Zoonotic Bacterial Pathogens. J. Trop. Med. 2020, 2020, 4674235. [Google Scholar] [CrossRef]
- Su, X.; Cao, G.; Zhang, J.; Pan, H.; Zhang, D.; Kuang, D.; Yang, X.; Xu, X.; Shi, X.; Meng, J. Characterization of internalin genes in Listeria monocytogenes from food and humans, and their association with the invasion of Caco-2 cells. Gut Pathog. 2019, 11, 30. [Google Scholar] [CrossRef]
- Manso, B.; Melero, B.; Stessl, B.; Fernandez-Natal, I.; Jaime, I.; Hernandez, M.; Wagner, M.; Rovira, J.; Rodriguez-Lazaro, D. Characterization of Virulence and Persistence Abilities of Listeria monocytogenes Strains Isolated from Food Processing Premises. J. Food Prot. 2019, 82, 1922–1930. [Google Scholar] [CrossRef]
- Ghebrehiwet, B.; Jesty, J.; Peerschke, E.I.B. gC1q-R/p33: Structure-Function Predictions from the Crystal Structure. Immunobiology 2002, 205, 421–432. [Google Scholar] [CrossRef]
- Zemser, R.B.; Martin, S.E. Heat stability of virulence-associated enzymes from Listeria monocytogenes SLCC 5764. J. Food Prot. 1998, 61, 899–902. [Google Scholar] [CrossRef]
- Schnupf, P.; Portnoy, D.A. Listeriolysin O: A phagosome-specific lysin. Microbes Infect. 2007, 9, 1176–1187. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Akgul, A.; Karsi, A.; Abdelhamed, H.; Wills, R.W.; Lawrence, M.L. The role of Listeria monocytogenes cell wall surface anchor protein LapB in virulence, adherence, and intracellular replication. Microb. Pathog. 2016, 92, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinner, S.; Kaltschmidt, J.; Stump-Guthier, C.; Hetjens, S.; Ishikawa, H.; Tenenbaum, T.; Schroten, H.; Schwerk, C. Mitogen-activated protein kinases are required for effective infection of human choroid plexus epithelial cells by Listeria monocytogenes. Microbes Infect. 2017, 19, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Drolia, R.; Tenguria, S.; Durkes, A.C.; Turner, J.R.; Bhunia, A.K. Listeria Adhesion Protein Induces Intestinal Epithelial Barrier Dysfunction for Bacterial Translocation. Cell Host Microbe 2018, 23, 470–484.e7. [Google Scholar] [CrossRef] [Green Version]
- Drolia, R.; Bhunia, A.K. Crossing the Intestinal Barrier via Listeria Adhesion Protein and Internalin A. Trends Microbiol. 2019, 27, 408–425. [Google Scholar] [CrossRef]
- Awuah, G.B.; Ramaswamy, H.S.; Economides, A. Thermal processing and quality: Principles and overview. Chem. Eng. Process. Process Intensif. 2007, 46, 584–602. [Google Scholar] [CrossRef]
- Misra, N.N.; Koubaa, M.; Roohinejad, S.; Juliano, P.; Alpas, H.; Inacio, R.S.; Saraiva, J.A.; Barba, F.J. Landmarks in the historical development of twenty first century food processing technologies. Food Res. Int. 2017, 97, 318–339. [Google Scholar] [CrossRef]
- Mtaoua, H.; Sánchez-Vega, R.; Ferchichi, A.; Martín-Belloso, O. Impact of High-Intensity Pulsed Electric Fields or Thermal Treatment on the Quality Attributes of Date Juice through Storage. J. Food Process. Preserv. 2017, 41, e13052. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Huang, Z.; Tong, J.; Wu, Q.; Pan, Y.; Malakar, P.K.; Zhao, Y. An outlook for food sterilization technology: Targeting the outer membrane of foodborne gram-negative pathogenic bacteria. Curr. Opin. Food Sci. 2021, 42, 15–22. [Google Scholar] [CrossRef]
- Pang, X.; Liu, X.; Cheng, Y.; Zhang, C.; Ren, E.; Liu, C.; Zhang, Y.; Zhu, J.; Chen, X.; Liu, G. Sono-Immunotherapeutic Nanocapturer to Combat Multidrug-Resistant Bacterial Infections. Adv. Mater. 2019, 31, e1902530. [Google Scholar] [CrossRef]
- McHale, A.P.; Callan, J.F.; Nomikou, N.; Fowley, C.; Callan, B. Sonodynamic Therapy: Concept, Mechanism and Application to Cancer Treatment. Adv. Exp. Med. Biol. 2016, 880, 429–450. [Google Scholar] [CrossRef]
- Akhtar, F.; Khan, A.U.; Misba, L.; Akhtar, K.; Ali, A. Antimicrobial and antibiofilm photodynamic therapy against vancomycin resistant Staphylococcus aureus (VRSA) induced infection in vitro and in vivo. Eur. J. Pharm. Biopharm. 2021, 160, 65–76. [Google Scholar] [CrossRef]
- Sadanala, K.C.; Chaturvedi, P.K.; Seo, Y.M.; Kim, J.M.; Jo, Y.S.; Lee, Y.K.; Ahn, W.S. Sono-Photodynamic Combination Therapy: A Review on Sensitizers. Anticancer Res. 2014, 34, 4657–4664. [Google Scholar]
- Niavarzi, S.; Pourhajibagher, M.; Khedmat, S.; Ghabraei, S.; Chiniforush, N.; Bahador, A. Effect of ultrasonic activation on the efficacy of antimicrobial photodynamic therapy: Evaluation of penetration depth of photosensitizer and elimination of Enterococcus faecalis biofilms. Photodiagnosis Photodyn. Ther. 2019, 27, 362–366. [Google Scholar] [CrossRef]
- Drantantiyas, N.; Astuti, S.D.; Nasution, A. Comparison microbial killing efficacy between sonodynamic therapy and photodynamic therapy. In Proceedings of the International Seminar on Photonics, Optics, & Its Applications, Bali, Indonesia, 24–25 August 2016. [Google Scholar]
- Lai, D.; Zhou, A.; Tan, B.K.; Tang, Y.; Sarah Hamzah, S.; Zhang, Z.; Lin, S.; Hu, J. Preparation and photodynamic bactericidal effects of curcumin-beta-cyclodextrin complex. Food Chem. 2021, 361, 130117. [Google Scholar] [CrossRef]
- Turrini, E.; Ferruzzi, L.; Fimognari, C. Natural compounds to overcome cancer chemoresistance: Toxicological and clinical issues. Expert Opin. Drug Metab. Toxicol. 2014, 10, 1677–1690. [Google Scholar] [CrossRef]
- Fan, L.; Idris Muhammad, A.; Bilyaminu Ismail, B.; Liu, D. Sonodynamic antimicrobial chemotherapy: An emerging alternative strategy for microbial inactivation. Ultrason. Sonochem. 2021, 75, 105591. [Google Scholar] [CrossRef]
- Alves, F.; Gomes Guimaraes, G.; Mayumi Inada, N.; Pratavieira, S.; Salvador Bagnato, V.; Kurachi, C. Strategies to Improve the Antimicrobial Efficacy of Photodynamic, Sonodynamic, and Sonophotodynamic Therapies. Lasers Surg. Med. 2021, 53, 1113–1121. [Google Scholar] [CrossRef]
- Pourhajibagher, M.; Bahador, A. Attenuation of Aggregatibacter actinomycetemcomitans virulence using curcumin-decorated nanophytosomes-mediated photo-sonoantimicrobial chemotherapy. Sci. Rep. 2021, 11, 6012. [Google Scholar] [CrossRef]
- Bhavya, M.L.; Hebbar, H.U. Sono-photodynamic inactivation of Escherichia coli and Staphylococcus aureus in orange juice. Ultrason. Sonochem. 2019, 57, 108–115. [Google Scholar] [CrossRef]
- Wang, D.; Zhou, F.; Lai, D.; Zhang, Y.; Hu, J.; Lin, S. Curcumin-mediated sono/photodynamic treatment preserved the quality of shrimp surimi and influenced its microbial community changes during refrigerated storage. Ultrason. Sonochem. 2021, 78, 105715. [Google Scholar] [CrossRef]
- Hu, J.; Lin, S.; Tan, B.K.; Hamzah, S.S.; Lin, Y.; Kong, Z.; Zhang, Y.; Zheng, B.; Zeng, S. Photodynamic inactivation of Burkholderia cepacia by curcumin in combination with EDTA. Food Res. Int. 2018, 111, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Su, H.-L.; Chou, C.-C.; Hung, D.-J.; Lin, S.-H.; Pao, I.C.; Lin, J.-H.; Huang, F.-L.; Dong, R.-X.; Lin, J.-J. The disruption of bacterial membrane integrity through ROS generation induced by nanohybrids of silver and clay. Biomaterials 2009, 30, 5979–5987. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Li, X.; Bi, W.; Jiang, C. Relationship between membrane damage, leakage of intracellular compounds, and inactivation of Escherichia coli treated by pressurized CO2. J. Basic Microbiol. 2014, 54, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Morales-de la Peña, M.; Welti-Chanes, J.; Martín-Belloso, O. Novel technologies to improve food safety and quality. Curr. Opin. Food Sci. 2019, 30, 1–7. [Google Scholar] [CrossRef]
- Wang, X.; Jia, Y.; Wang, P.; Liu, Q.; Zheng, H. Current status and future perspectives of sonodynamic therapy in glioma treatment. Ultrason. Sonochem. 2017, 37, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Hirschberg, H.; Madsen, S.J. Synergistic efficacy of ultrasound, sonosensitizers and chemotherapy: A review. Ther. Deliv. 2017, 8, 331–342. [Google Scholar] [CrossRef]
- Nakonechny, F.; Nisnevitch, M.; Nitzan, Y.; Nisnevitch, M. Sonodynamic excitation of Rose Bengal for eradication of gram-positive and gram-negative bacteria. Biomed. Res. Int. 2013, 2013, 684930. [Google Scholar] [CrossRef]
- Lichtenberg, D.; Pinchuk, I. Oxidative stress, the term and the concept. Biochem. Biophys. Res. Commun. 2015, 461, 441–444. [Google Scholar] [CrossRef]
- Wu, M.; Zhang, Z.; Liu, Z.; Zhang, J.; Zhang, Y.; Ding, Y.; Huang, T.; Xiang, D.; Wang, Z.; Dai, Y.; et al. Piezoelectric nanocomposites for sonodynamic bacterial elimination and wound healing. Nano Today 2021, 37, 101104. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Storey, K.B. Oxidative stress concept updated: Definitions, classifications, and regulatory pathways implicated. EXCLI J. 2021, 20, 956–967. [Google Scholar] [CrossRef]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef]
- Xu, F.; Hu, M.; Liu, C.; Choi, S.K. Yolk-structured multifunctional up-conversion nanoparticles for synergistic photodynamic-sonodynamic antibacterial resistance therapy. Biomater. Sci. 2017, 5, 678–685. [Google Scholar] [CrossRef]
- Li, X.-F.; Feng, X.-Q.; Yang, S.; Fu, G.-Q.; Wang, T.-P.; Su, Z.-X. Chitosan kills Escherichia coli through damage to be of cell membrane mechanism. Carbohydr. Polym. 2010, 79, 493–499. [Google Scholar] [CrossRef]
- Costley, D.; Nesbitt, H.; Ternan, N.; Dooley, J.; Huang, Y.Y.; Hamblin, M.R.; McHale, A.P.; Callan, J.F. Sonodynamic inactivation of Gram-positive and Gram-negative bacteria using a Rose Bengal-antimicrobial peptide conjugate. Int. J. Antimicrob. Agents 2017, 49, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Alves, F.; Ayala, E.T.P.; Pratavieira, S. Sonophotodynamic Inactivation: The power of light and ultrasound in the battle against microorganisms. J. Photochem. Photobiol. 2021, 7, 100039. [Google Scholar] [CrossRef]
- Ray, S.K.; Dhakal, D.; Hur, J.; Lee, S.W. Visible light driven MoS2/alpha-NiMoO4 ultra-thin nanoneedle composite for efficient Staphylococcus aureus inactivation. J. Hazard. Mater. 2020, 385, 121553. [Google Scholar] [CrossRef]
- Wu, J.; Mou, H.; Xue, C.; Leung, A.W.; Xu, C.; Tang, Q.J. Photodynamic effect of curcumin on Vibrio parahaemolyticus. Photodiagnosis Photodyn. Ther. 2016, 15, 34–39. [Google Scholar] [CrossRef]
- Pourhajibagher, M.; Rahimi Esboei, B.; Hodjat, M.; Bahador, A. Sonodynamic excitation of nanomicelle curcumin for eradication of Streptococcus mutans under sonodynamic antimicrobial chemotherapy: Enhanced anti-caries activity of nanomicelle curcumin. Photodiagn. Photodyn. Ther. 2020, 30, 101780. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zheng, P.; Li, J.; Yang, Y.; Zeng, S.; Qiu, J.; Lin, S. Curcumin-Mediated Sono-Photodynamic Treatment Inactivates Listeria monocytogenes via ROS-Induced Physical Disruption and Oxidative Damage. Foods 2022, 11, 808. https://doi.org/10.3390/foods11060808
Zhang J, Zheng P, Li J, Yang Y, Zeng S, Qiu J, Lin S. Curcumin-Mediated Sono-Photodynamic Treatment Inactivates Listeria monocytogenes via ROS-Induced Physical Disruption and Oxidative Damage. Foods. 2022; 11(6):808. https://doi.org/10.3390/foods11060808
Chicago/Turabian StyleZhang, Jiawen, Peiying Zheng, Jing Li, Yijing Yang, Shaoxiao Zeng, Jianqing Qiu, and Shaoling Lin. 2022. "Curcumin-Mediated Sono-Photodynamic Treatment Inactivates Listeria monocytogenes via ROS-Induced Physical Disruption and Oxidative Damage" Foods 11, no. 6: 808. https://doi.org/10.3390/foods11060808