
Dynamics Changes of Microorganisms Community and Fermentation Quality in Soybean Meal Prepared with Lactic Acid Bacteria and Artemisia argyi through Fermentation and Aerobic Exposure Processes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Fermentation Preparation
2.2. pH, Chemical Composition, Crude Flavonoids and Microbial Population Analysis of Soybean Meal and Artemisia argyi
2.3. Evaluation of Sensory and Surface Structure of Soybean Meal and Fermented Soybean Meal
2.4. Fermentation Quality, Chemical Composition and Microbial Population Analysis of Fermented Soybean Meal
2.5. Bacterial and Fungal Community Analysis
2.6. Mycotoxin and Crude Flavonoids Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Characteristics of Soybean Meal and Artemisia argyi
3.2. Sensory Evaluation and Surface Structure of Fermented Soybean Meal
3.3. Fermentation Quality, Chemical Composition and Microbial Population of Fermented Soybean Meal
3.3.1. Fermentation Quality
3.3.2. Chemical Composition
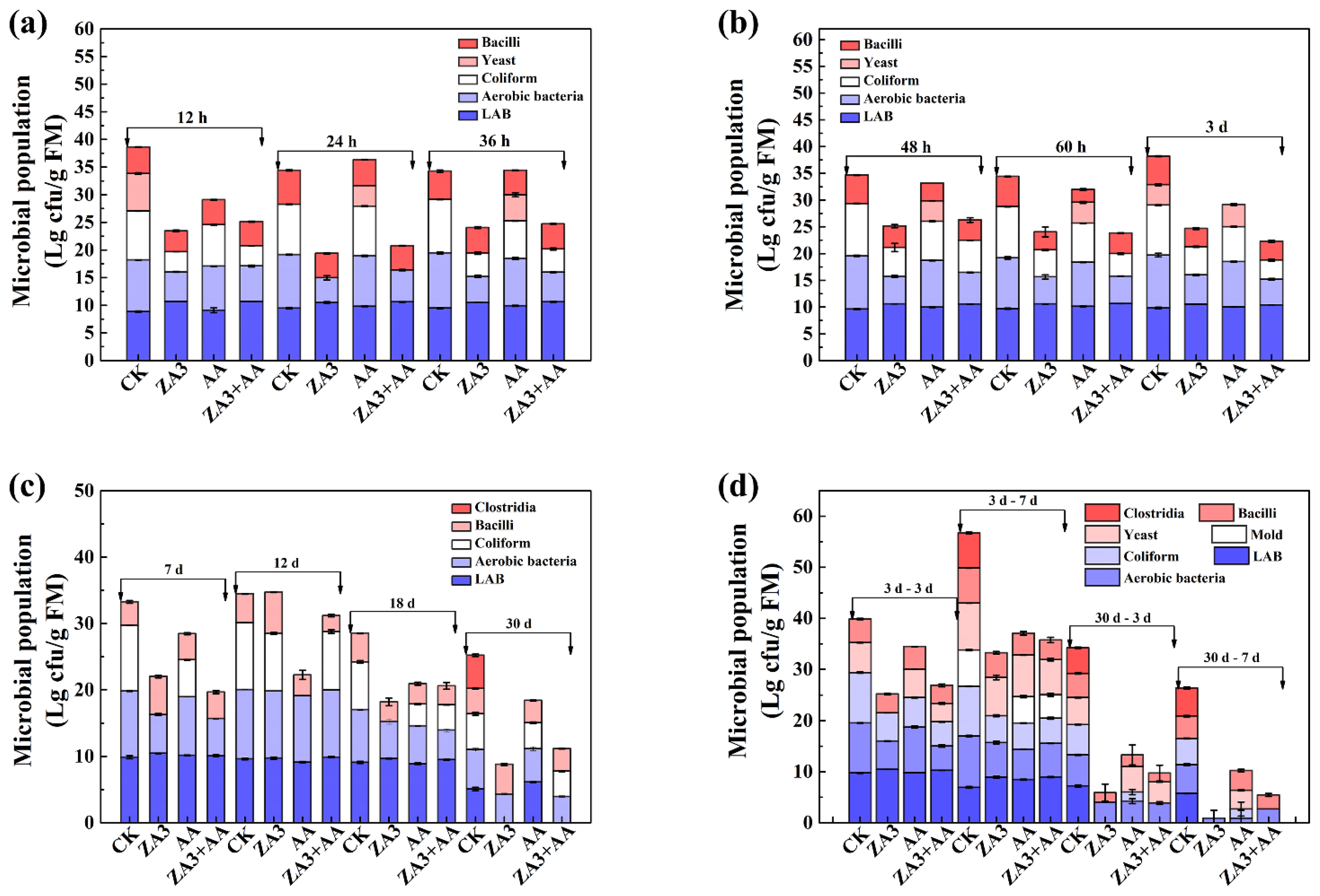
3.3.3. Microbial Population
3.4. Microbial Communities, Diversities and Relative Abundances in Fermented Soybean Meal
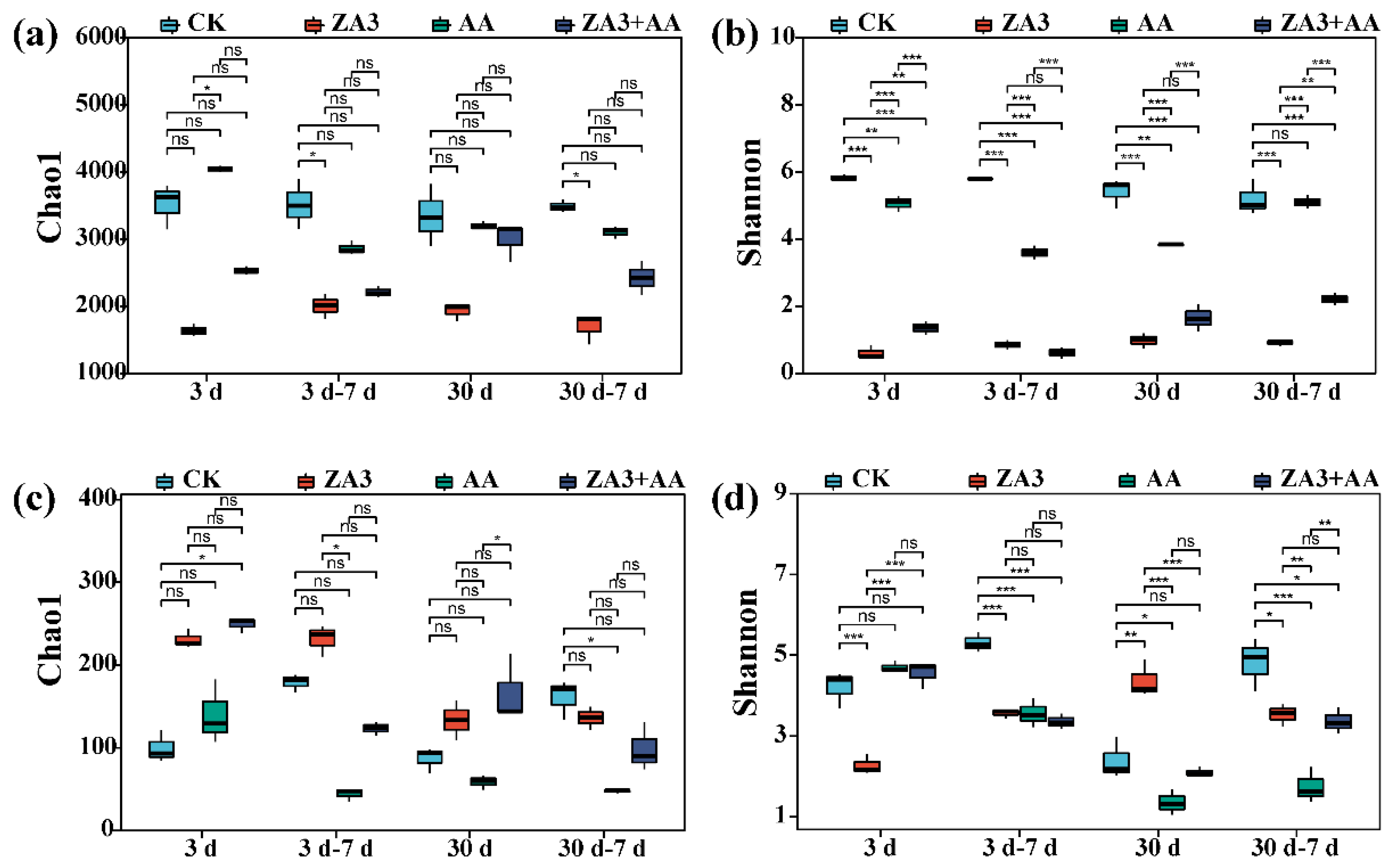
3.4.1. Microbial Communities and Diversities
3.4.2. Relative Abundances at the Phylum Level
3.4.3. Relative Abundances at the Genus Level
3.5. Correlation Analyses of the Bacterial and Fungal Communities with Fermentation Properties of Fermented Soybean Meal
3.6. Toxin Content
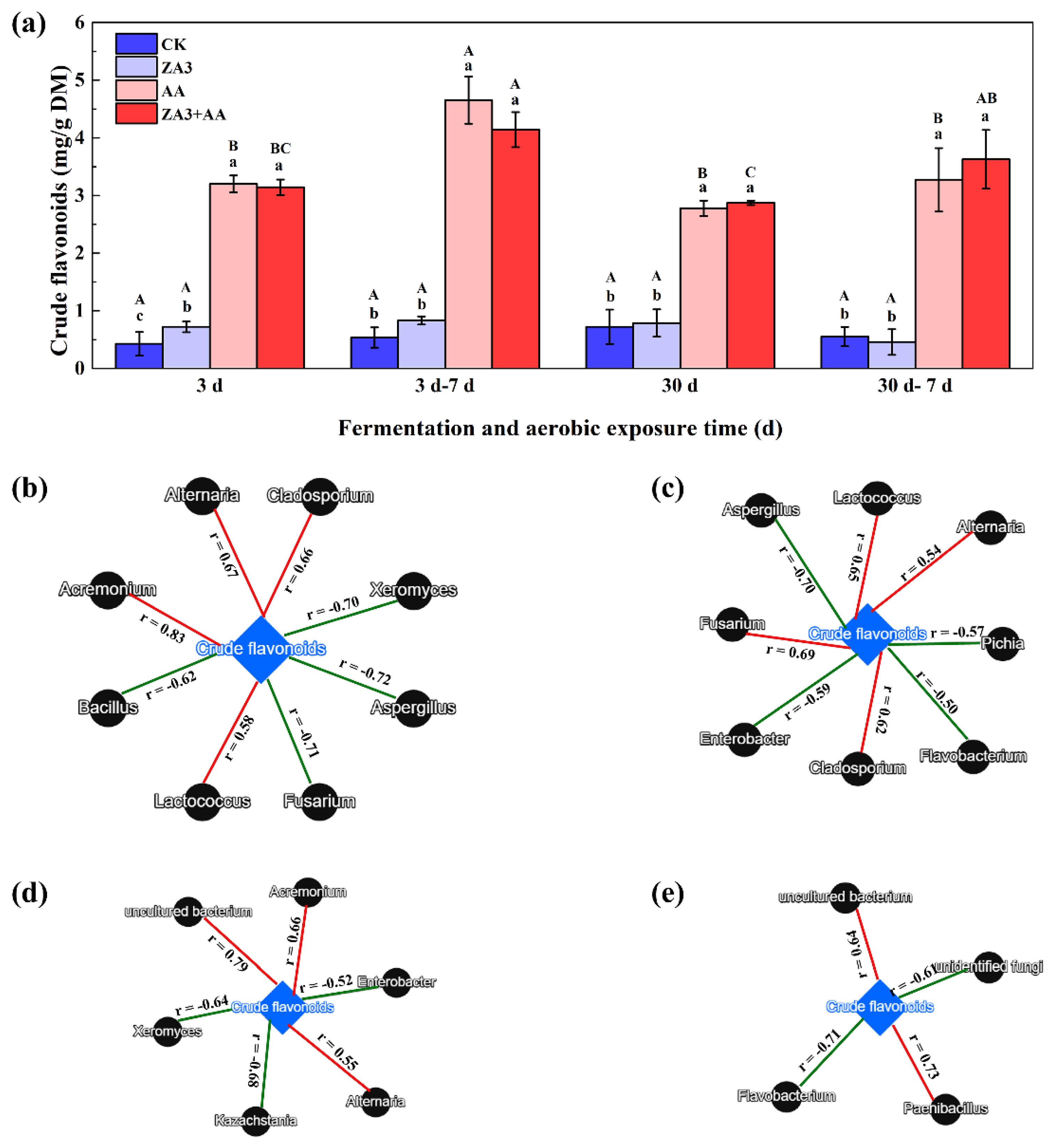
3.7. Determination of Crude Flavonoids and Linkages with Microbial Communities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maruyama, N.; Sato, R.; Wada, Y.; Matsumura, Y.; Goto, H.; Okuda, E.; Nakagawa, S.; Utsumi, S. Structure-physicochemical function relationships of soybean β-conglycinin constituent subunits. J. Agric. Food Chem. 1999, 47, 5278–5284. [Google Scholar] [CrossRef] [PubMed]
- Czech, A.; Grela, E.R.; Nowakowicz-Debek, B.; Wlazlo, L. The effects of a fermented rapeseed meal or/and soybean meal additive on the blood lipid profile and immune parameters of piglets and on minerals in their blood and bone. PLoS ONE 2021, 16, e0253744. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Garg, P.; Kumar, P.; Bhatia, S.K.; Kulshrestha, S. Microbial fermentation and its role in quality improvement of fermented foods. Fermentation 2020, 6, 106. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, Z.; Yu, W.; Zheng, L.; Li, L.; Gu, W.; Xu, H.; Wei, B.; Yan, X. Nutritional quality improvement of soybean meal by Bacillus velezensis and Lactobacillus plantarum during two-stage solid- state fermentation. AMB Express 2021, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shi, C.; Su, W.; Jin, M.; Xu, B.; Hao, L.; Zhang, Y.; Lu, Z.; Wang, F.; Wang, Y.; et al. Dynamics of the physicochemical characteristics, microbiota, and metabolic functions of soybean meal and corn mixed substrates during two-stage solid-state fermentation. mSystems 2020, 5, e00501-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 2019, 275, 280–287. [Google Scholar] [CrossRef]
- Huang, C.; Chen, C.; Chang, S.; Tsai, G. Evaluation of antiobesity activity of soybean meal products fermented by Lactobacillus plantarum FPS 2520 and Bacillus subtilis N1 in rats fed with high-fat diet. J. Med. Food 2020, 23, 667–675. [Google Scholar] [CrossRef]
- Tsai, C.; Lin, L.; Wang, C.; Tsai, C.; Chang, S.; Lee, T. Assessment of intestinal immunity and permeability of broilers on partial replacement diets of two-stage fermented soybean meal by Bacillus velezensis and Lactobacillus brevis ATCC 367. Animals 2021, 11, 2336. [Google Scholar] [CrossRef]
- Shi, C.; Zhang, Y.; Yin, Y.; Wang, C.; Wang, Y. Amino acid and phosphorus digestibility of fermented corn-soybean meal mixed feed with Bacillus subtilis and Enterococcus faecium fed to pigs. J. Anim. Sci. 2017, 95, 3996–4004. [Google Scholar] [CrossRef]
- Yang, H.; Qu, Y.; Li, J.; Liu, X.; Wu, R.; Wu, J. Improvement of the protein quality and degradation of allergens in soybean meal by combination fermentation and enzymatic hydrolysis. LWT Food Sci. Technol. 2020, 128, 109442. [Google Scholar] [CrossRef]
- Nishino, N.; Hattori, H. Resistance to aerobic deterioration of total mixed ration silage inoculated with and without homofermentative or heterofermentative lactic acid bacteria. J. Sci. Food Agric. 2007, 87, 2420–2426. [Google Scholar] [CrossRef]
- Heng, X.; Chen, H.; Lu, C.; Feng, T.; Li, K.; Gao, E. Study on synergistic fermentation of bean dregs and soybean meal by multiple strains and proteases. LWT-Food Sci. Technol. 2022, 154, 112626. [Google Scholar] [CrossRef]
- Guan, X.; Ge, D.; Li, S.; Huang, K.; Liu, J.; Li, F. Chemical composition and antimicrobial activities of Artemisia argyi Levl. et vant essential oils extracted by simultaneous distillation-extraction, subcritical extraction and hydrodistillation. Molecules 2019, 24, 483. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Kim, C.E.; Park, S.Y.; Kim, K.O.; Hiep, N.T.; Lee, D.; Jang, H.J.; Lee, J.W.; Kang, K.S. Protective effect of Artemisia argyi and its flavonoid constituents against contrast-induced cytotoxicity by iodixanol in LLC-PK1 cells. Int. J. Mol. Sci. 2018, 19, 1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menz, J.; Olsson, O.; Kummerer, K. Antibiotic residues in livestock manure: Does the EU risk assessment sufficiently protect against microbial toxicity and selection of resistant bacteria in the environment? J. Hazard. Mater. 2019, 379, 120807. [Google Scholar] [CrossRef]
- Beigh, Y.A.; Ganai, A.M. Potential of wormwood (Artemisia absinthium Linn.) herb for use as additive in livestock feeding: A review. Pharma. Innov. 2017, 6, 176–187. [Google Scholar]
- Wang, W.; Ma, H.; Yu, H.; Qin, G.; Tan, Z.; Wang, Y.; Pang, H. Screening of Lactobacillus plantarum subsp. plantarum with potential probiotic activities for inhibiting ETEC K88 in weaned piglets. Molecules 2020, 25, 4481. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Hudz, N.; Yezerska, O.; Grygorieva, O.; Brindza, J.; Felsöciová, S.; Kačániová, M.; Wieczorek, P. Analytical procedure elaboration of total flavonoid content determination and antimicrobial activity of bee bread extracts. Acta Pol. Pharm. 2019, 76, 439–452. [Google Scholar] [CrossRef]
- Pang, H.; Qin, G.; Tan, Z.; Li, Z.; Wang, Y.; Cai, Y. Natural populations of lactic acid bacteria associated with silage fermentation as determined by phenotype, 16S ribosomal RNA and recA gene analysis. Syst. Appl. Microbiol. 2011, 34, 235–241. [Google Scholar] [CrossRef]
- Meinlschmidt, P.; Ueberham, E.; Lehmann, J.; Schweiggert-Weisz, U.; Eisner, P. Immunoreactivity, sensory and physico-chemical properties of fermented soy protein isolate. Food Chem. 2016, 205, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Matias, I.; Buosi, A.S.; Gomes, F.C. Functions of flavonoids in the central nervous system: Astrocytes as targets for natural compounds. Neurochem. Int. 2016, 95, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhu, H.; Zhang, B.; Chen, J.; Ao, Q.; Wang, X. XRD, SEM, and XPS Analysis of soybean protein powders obtained through extraction involving reverse micelles. J. Am. Oil Chem. Soc. 2015, 92, 975–983. [Google Scholar] [CrossRef]
- Ferrero, F.; Piano, S.; Tabacco, E.; Borreani, G. Effects of conservation period and Lactobacillus hilgardii inoculum on the fermentation profile and aerobic stability of whole corn and sorghum silages. J. Sci. Food Agric. 2019, 99, 2530–2540. [Google Scholar] [CrossRef]
- Bai, J.; Xu, D.; Xie, D.; Wang, M.; Li, Z.; Guo, X. Effects of antibacterial peptide-producing Bacillus subtilis and Lactobacillus buchneri on fermentation, aerobic stability, and microbial community of alfalfa silage. Bioresour. Technol. 2020, 315, 123881. [Google Scholar] [CrossRef]
- Guan, H.; Ran, Q.; Li, H.; Zhang, X. Succession of microbial communities of corn silage inoculated with heterofermentative lactic acid bacteria from ensiling to aerobic exposure. Fermentation 2021, 7, 258. [Google Scholar] [CrossRef]
- Su, W.; Jiang, Z.; Hao, L.; Li, W.; Gong, T.; Zhang, Y.; Du, S.; Wang, C.; Lu, Z.; Jin, M.; et al. Variations of soybean meal and corn mixed substrates in physicochemical characteristics and microbiota during two-stage solid-state fermentation. Front. Microbiol. 2021, 12, 688839. [Google Scholar] [CrossRef]
- Arriola, K.G.; Queiroz, O.C.M.; Romero, J.J.; Casper, D.; Muniz, E.; Hamie, J.; Adesogan, A.T. Effect of microbial inoculants on the quality and aerobic stability of bermudagrass round-bale haylage. J. Dairy Sci. 2015, 98, 478–485. [Google Scholar] [CrossRef]
- Wang, T.; Teng, K.; Cao, Y.; Shi, W.; Xuan, Z.; Zhou, J.; Zhang, J.; Zhong, J. Effects of Lactobacillus hilgardii 60TS-2, with or without homofermentative Lactobacillus plantarum B90, on the aerobic stability, fermentation quality and microbial community dynamics in sugarcane top silage. Bioresour. Technol. 2020, 312, 123600. [Google Scholar] [CrossRef]
- Sonker, N.; Pandey, A.K.; Singh, P. Efficiency of Artemisia nilagirica (Clarke) Pamp. essential oil as a mycotoxicant against postharvest mycobiota of table grapes. J. Sci. Food Agric. 2015, 95, 1932–1939. [Google Scholar] [CrossRef]
- Petretto, G.L.; Chessa, M.; Piana, A. Chemical and biological study on the essential oil of Artemisia caerulescens L. ssp. densiflora (Viv.). Nat. Prod. Res. 2013, 27, 1709–1715. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.J.C. The Biochemistry of Silage (2nd edn). J. Agric. Sci. 1991, 117, 386. [Google Scholar] [CrossRef]
- Namihira, T.; Shinzato, N.; Akamine, H.; Maekawa, H.; Matsui, T. Influence of nitrogen fertilization on tropical-grass silage assessed by ensiling process monitoring using chemical and microbial community analyses. J. Appl. Microbiol. 2010, 108, 1954–1965. [Google Scholar] [CrossRef] [PubMed]
- Preston, G.M.; Studholme, D.J.; Caldelari, I. Profiling the secretomes of plant pathogenic Proteobacteria. FEMS Microbiol. Rev. 2005, 29, 331–360. [Google Scholar] [CrossRef] [PubMed]
- Keshri, J.; Chen, Y.R.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biot. 2018, 102, 4025–4037. [Google Scholar] [CrossRef]
- Ogunade, I.M.; Jiang, Y.; Pech Cervantes, A.A.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157: H7 and silage additives. J. Dairy Sci. 2018, 101, 2048–2059. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Ke, W.C.; Vyas, D.; Adesogan, A.T.; Franco, M.; Li, F.H.; Bai, J.; Guo, X.S. Antioxidant status, chemical composition and fermentation profile of alfalfa silage ensiled at two dry matter contents with a novel Lactobacillus plantarum strain with high-antioxidant activity. Anim. Feed Sci. Tech. 2021, 272, 114751. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, H.T.; Jian, K.L. Outline of Ascomycota: 2017. Fungal Divers. 2017, 88, 167–263. [Google Scholar] [CrossRef]
- Wang, X.; Liu, X.; Groenewald, J.Z. Phylogeny of anaerobic fungi (phylum Neocallimastigomycota), with contributions from yak in China. Anton. Leeuw. Int. J. Gen. 2017, 110, 87–103. [Google Scholar] [CrossRef] [Green Version]
- Muck, R. Silage microbiology and its control through additives. Rev. Bras. Zootec. 2010, 39, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Hashemi, M. Aflatoxin B1 levels in feedstuffs from dairy cow farms in south of Iran. Food Agric. Immunol. 2015, 27, 251–258. [Google Scholar] [CrossRef]
- Roncero, I.G.M.; Concepción, H.; Manuel, R.R.; Maceira, G.; Marta, M. Fusarium as a model for studying virulence in soilborne plant pathogens. Physiol. Mol. Plant 2003, 62, 87–98. [Google Scholar] [CrossRef]
- Moore, G.G.; Fapohunda, S.O. Molecular investigations of food-borne Cladosporium and Fusarium species from nigeria. J. Bacteriol. Mycol. 2016, 3, 1024. [Google Scholar]
- Liu, L.; Long, L.K.; An, Y.; Yang, J.; Xu, X.; Hu, C.H.; Liu, G. The thioredoxin reductase-encoding gene ActrxR1 is involved in the cephalosporin C production of Acremonium chrysogenum in methionine-supplemented medium. Appl. Microbiol. Biotechnol. 2013, 97, 2551–2562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, X.; Gu, Q.; Liang, M.; Mu, S.; Zhou, B.; Huang, F.; Lin, B.; Zou, C. Analysis of the correlation between bacteria and fungi in sugarcane tops silage prior to and after aerobic exposure. Bioresour. Technol. 2019, 291, 121835. [Google Scholar] [CrossRef]
- Xu, S.; Yang, J.; Qi, M.; Smiley, B.; Rutherford, W.; Wang, Y.; McAllister, T.A. Impact of Saccharomyces cerevisiae and Lactobacillus buchneri on microbial communities during ensiling and aerobic spoilage of corn silage. J. Anim. Sci. 2019, 97, 1273–1285. [Google Scholar] [CrossRef]
- Fabiszewska, A.U.; Zielinska, K.J.; Wrobel, B. Trends in designing microbial silage quality by biotechnological methods using lactic acid bacteria inoculants: A minireview. World J. Microbiol. Biotechnol. 2019, 35, 76. [Google Scholar] [CrossRef] [Green Version]
- Pahlow, G.; Muck, R.E.; Driehuis, F.; Elferink, O.; Spoelstra, S.F. Microbiology of ensiling. Agronomy 2003, 42, 31–94. [Google Scholar] [CrossRef]
- Duan, J.; Yin, J.; Wu, M.; Liao, P.; Deng, D.; Liu, G.; Wen, Q.; Wang, Y.; Qiu, W.; Liu, Y. Dietary glutamate supplementation ameliorates mycotoxin-induced abnormalities in the intestinal structure and expression of amino acid transporters in young pigs. PLoS ONE 2014, 9, e112357. [Google Scholar] [CrossRef] [Green Version]
- Zychowski, K.E.; Pohlenz, C.; Mays, T.; Romoser, A.; Hume, M.; Buentello, A.; Gatlin, D.M.; Phillips, T.D. The effect of NovaSil dietary supplementation on the growth and health performance of Nile tilapia (Oreochromis niloticus) fed aflatoxin-B1 contaminated feed. Aquaculture 2013, 376-379, 117–123. [Google Scholar] [CrossRef]
- Turbic, A.; Ahokas, J.T.; Haskard, C.A. Selective in vitro binding of dietary mutagens, individually or in combination, by lactic acid bacteria. Food Addit. Contam. A 2002, 19, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Bueno, D.J.; Casale, C.H.; Pizzolitto, R.P.; Salvano, M.A.; Oliver, G. Physical adsorption of aflatoxin B1 by lactic acid bacteria and Saccharomyces cerevisiae: A theoretical model. J. Food Prot. 2007, 70, 2148–2154. [Google Scholar] [CrossRef] [PubMed]
- Hoensch, H.P.; Oertel, R. The value of flavonoids for the human nutrition: Short review and perspectives. Clin. Nutr. Exp. 2015, 3, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Li, H.; Qin, X.; Wang, T.; Hao, J.; Zhao, J.; Wang, J.; Wang, R.; Wang, D.; Wei, S.; et al. Antibacterial evaluation of plants extracts against ampicillin-resistant Escherichia coli (E. coli) by microcalorimetry and principal component analysis. AMB Express 2019, 9, 101. [Google Scholar] [CrossRef] [PubMed]
- Simard, F.; Gauthier, C.; Legault, J.; Lavoie, S.; Mshvildadze, V.; Pichette, A. Structure elucidation of anti-methicillin resistant Staphylococcus aureus (MRSA) flavonoids from balsam poplar buds. Bioorg. Med. Chem. 2016, 24, 4188–4198. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | SBM | AA |
|---|---|---|
| pH | 6.7 ± 0.02 | 5.6 ± 0.01 |
| DM (g/kg) | 882.1 ± 2.81 | 371.6 ± 3.77 |
| CP (%DM) | 43.8 ± 0.02 | 10.9 ± 0.02 |
| CF (%DM) | 6.8 ± 0.07 | 53.8 ± 0.84 |
| EE (%DM) | 1.6 ± 0.07 | 3.1 ± 0.08 |
| Crude flavonoids (mg/g DM) | 0.6 ± 0.06 | 23.7 ± 1.84 |
| LAB (lg cfu/g FM) | 7.0 ± 0.02 | 5.6 ± 0.08 |
| Aerobic bacteria (lg cfu/g FM) | 8.4 ± 0.23 | 5.8 ± 0.05 |
| Coliform bacteria (lg cfu/g FM) | 5.0 ± 0.13 | 5.6 ± 0.46 |
| Bacilli (lg cfu/g FM) | 4.5 ± 0.19 | 6.6 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Tan, Z.; Gu, L.; Ma, H.; Wang, Z.; Wang, L.; Wu, G.; Qin, G.; Wang, Y.; Pang, H. Dynamics Changes of Microorganisms Community and Fermentation Quality in Soybean Meal Prepared with Lactic Acid Bacteria and Artemisia argyi through Fermentation and Aerobic Exposure Processes. Foods 2022, 11, 795. https://doi.org/10.3390/foods11060795
Wang W, Tan Z, Gu L, Ma H, Wang Z, Wang L, Wu G, Qin G, Wang Y, Pang H. Dynamics Changes of Microorganisms Community and Fermentation Quality in Soybean Meal Prepared with Lactic Acid Bacteria and Artemisia argyi through Fermentation and Aerobic Exposure Processes. Foods. 2022; 11(6):795. https://doi.org/10.3390/foods11060795
Chicago/Turabian StyleWang, Weiwei, Zhongfang Tan, Lingbiao Gu, Hao Ma, Zhenyu Wang, Lei Wang, Guofang Wu, Guangyong Qin, Yanping Wang, and Huili Pang. 2022. "Dynamics Changes of Microorganisms Community and Fermentation Quality in Soybean Meal Prepared with Lactic Acid Bacteria and Artemisia argyi through Fermentation and Aerobic Exposure Processes" Foods 11, no. 6: 795. https://doi.org/10.3390/foods11060795