
Soft-Shelled Turtle Peptide Supplementation Modifies Energy Metabolism and Oxidative Stress, Enhances Exercise Endurance, and Decreases Physical Fatigue in Mice

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Experiment Protocol
2.3. Body Weight and Organ Index
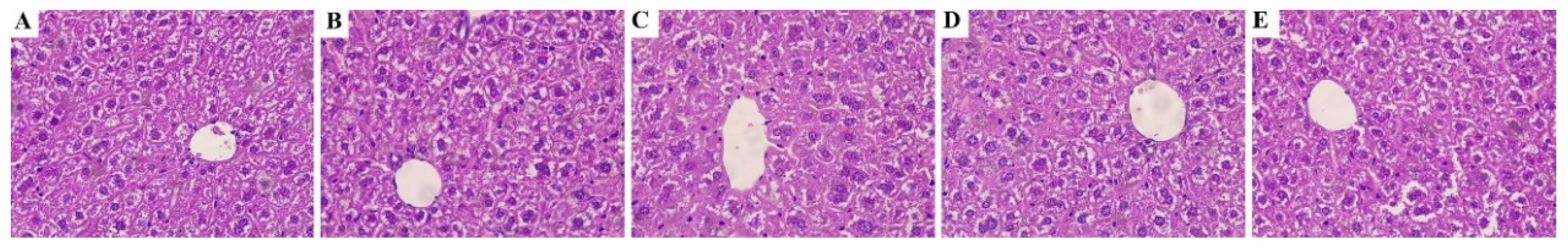
2.4. Histological Observation
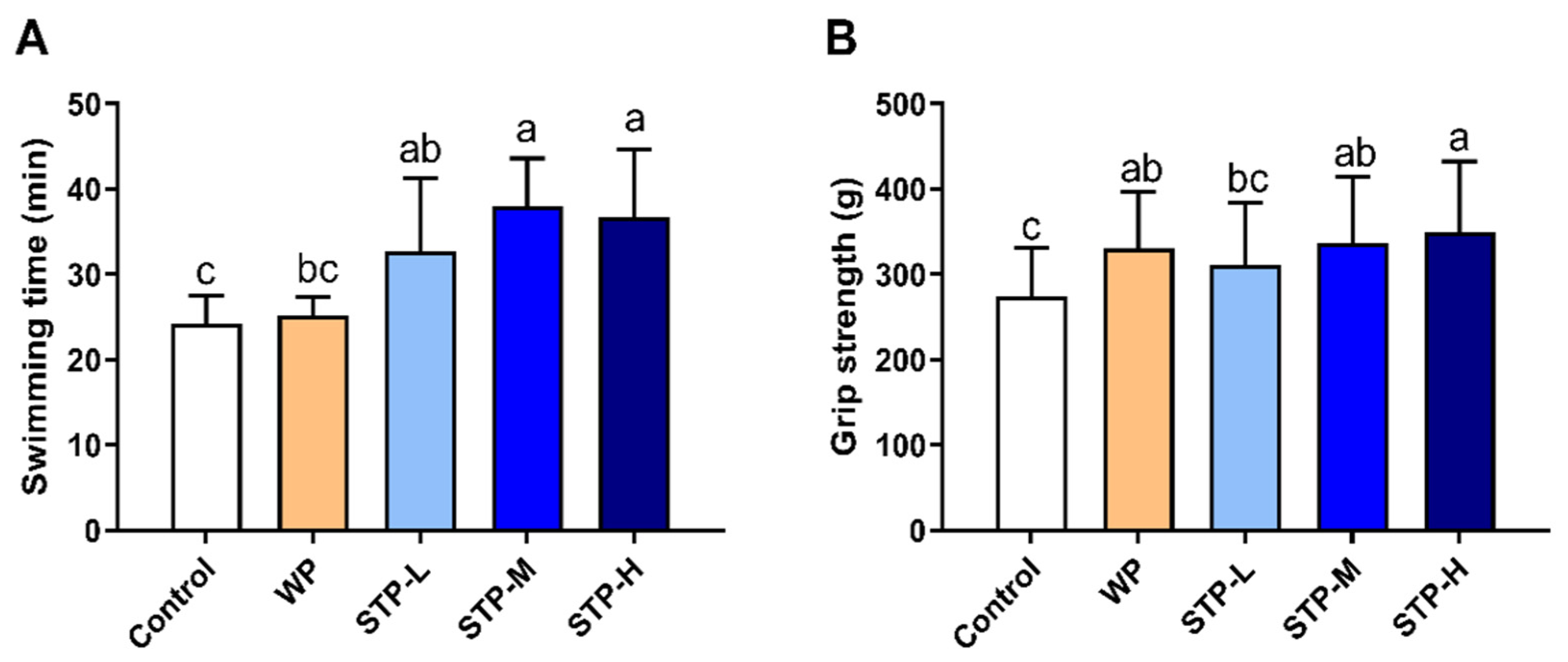
2.5. Weight-Loaded Forced Swimming Test
2.6. Forelimb Grip Strength Test
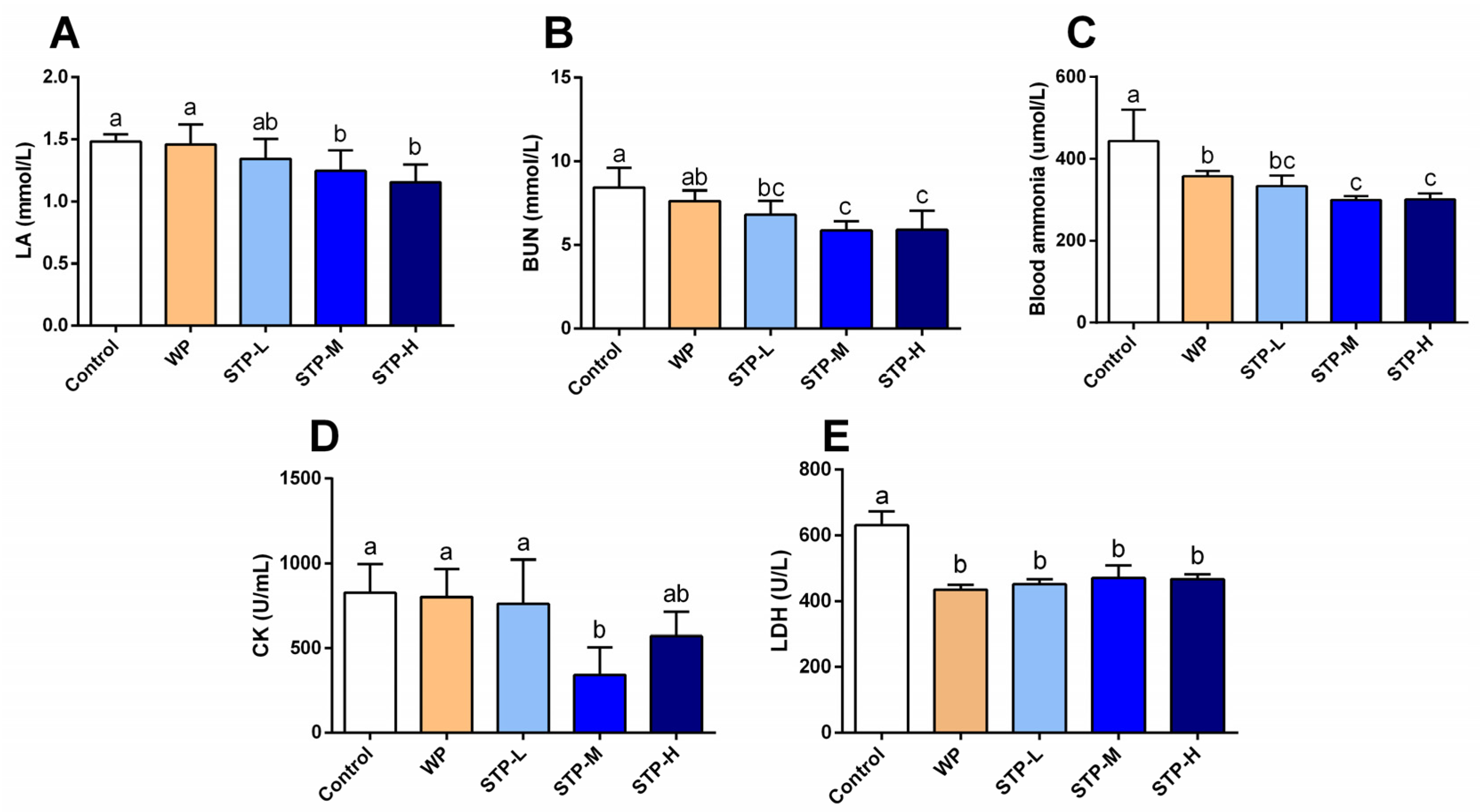
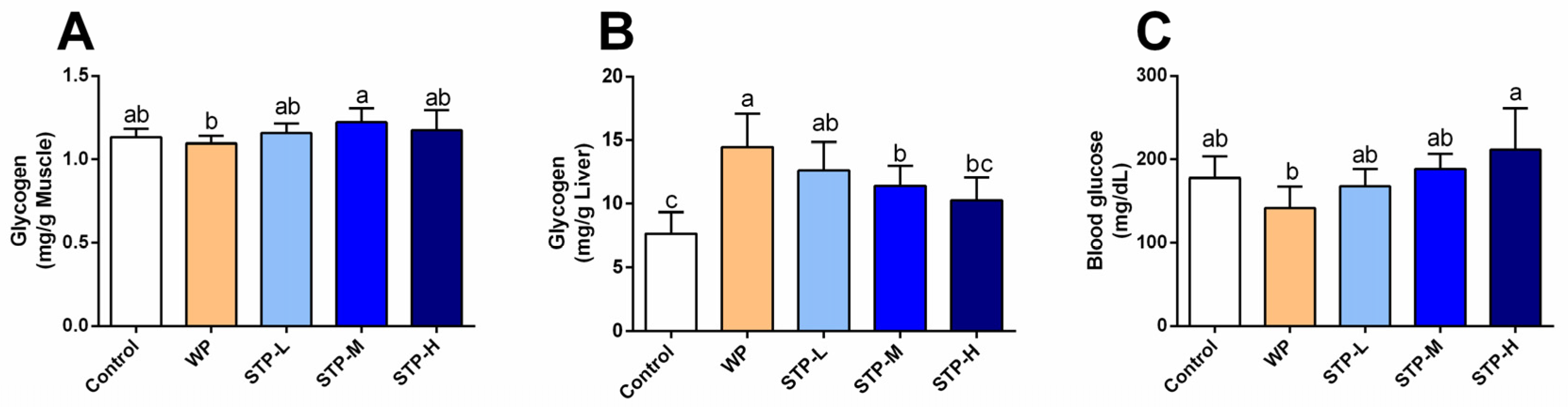
2.7. Determination of Biochemical Indicators
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. STP Augment Physiological Indexes in Mice
3.2. STP Would Not Affect Morphological and Pathological Changes
3.3. STP Could Improve Exercise Tolerance
3.4. STP Modified Metabolism Accumulation
3.5. STP Increased Energy Storage in Mice
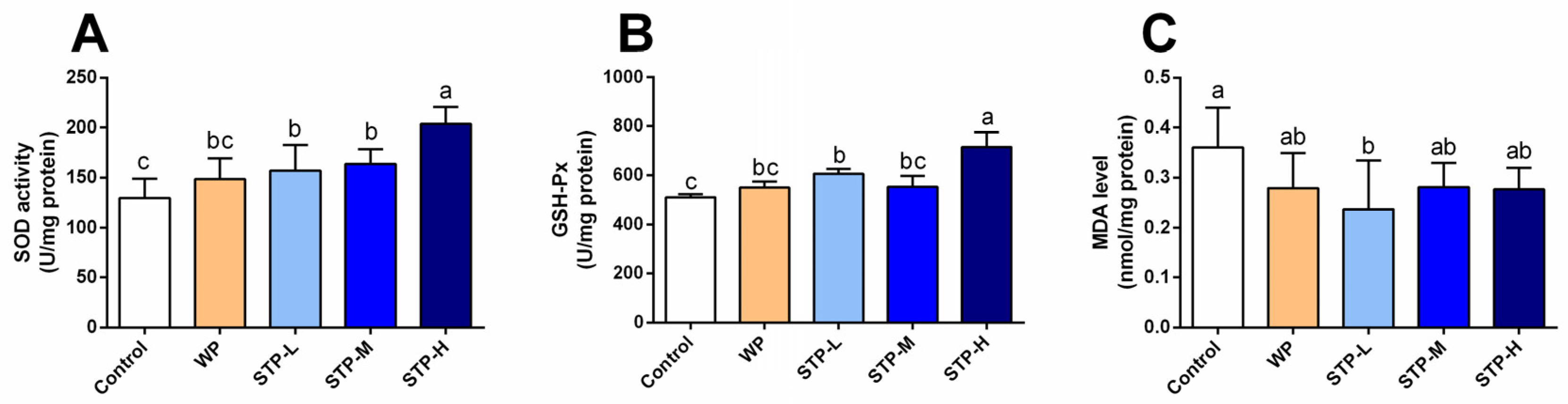
3.6. STP Exerted Antioxidant Activity
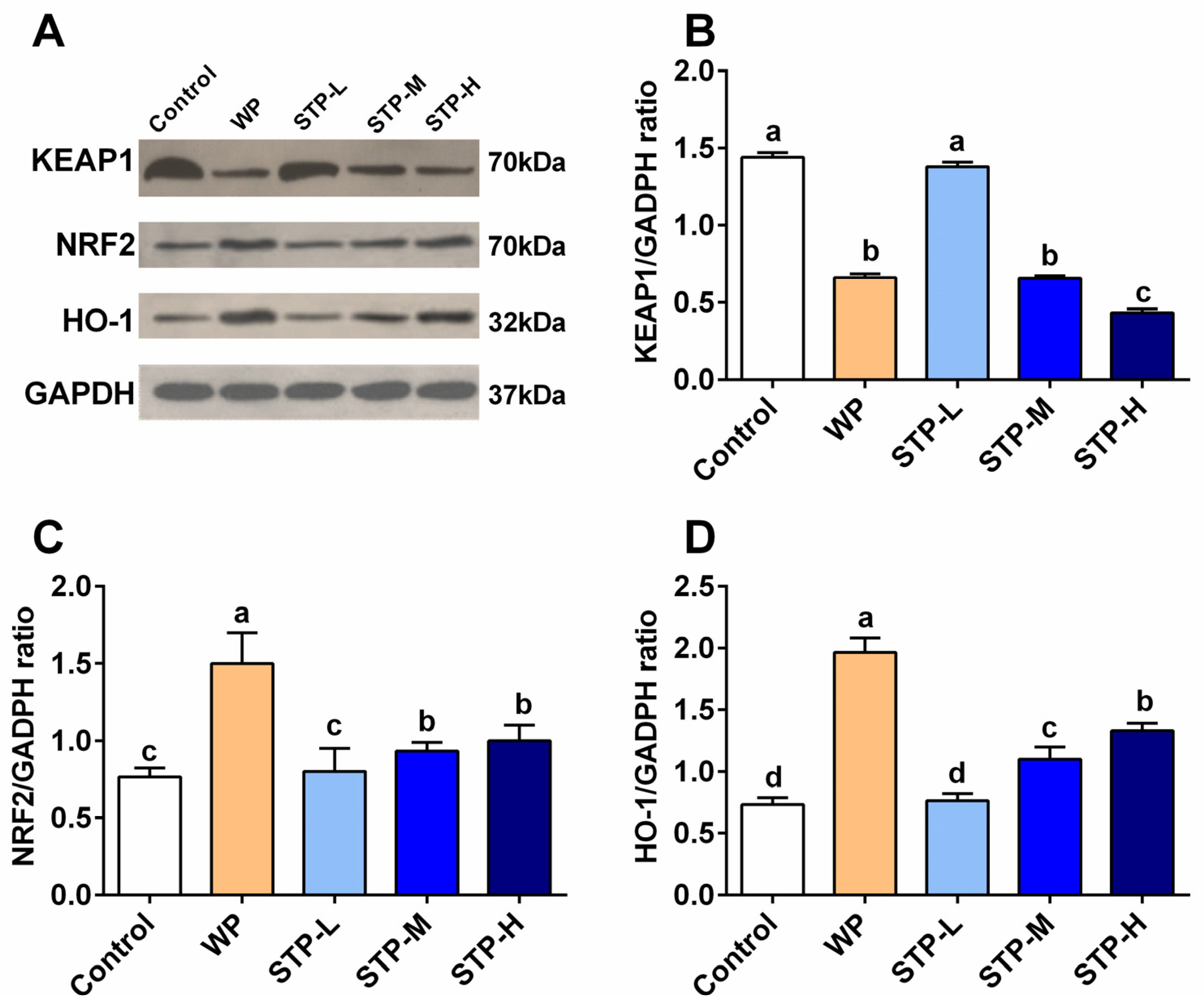
3.7. STP Modulated the NRF2/KEAP1 Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanaka, M.; Baba, Y.; Kataoka, Y.; Kinbara, N.; Sagesaka, Y.M.; Kakuda, T.; Watanabe, Y. Effects of (−)-epigallocatechin gallate in liver of an animal model of combined (physical and mental) fatigue. Nutrition 2008, 24, 599–603. [Google Scholar] [CrossRef]
- Yuan, T.; Wu, D.; Sun, K.; Tan, X.; Wang, J.; Zhao, T.; Ren, B.; Zhao, B.; Liu, Z.; Liu, X. Anti-Fatigue activity of aqueous extracts of Sonchus arvensis L. in exercise trained mice. Molecules 2019, 24, 1168. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, H.-L.; Lu, R.; Zhou, Y.-J.; Ma, R.; Lv, J.-Q.; Li, X.-L.; Chen, L.-J.; Yao, Z. The decapeptide CMS001 enhances swimming endurance in mice. Peptides 2008, 29, 1176–1182. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. In vitro antioxidant activity and in vivo anti-fatigue effect of loach (Misgurnus anguillicaudatus) peptides prepared by papain digestion. Food Chem. 2011, 124, 188–194. [Google Scholar] [CrossRef]
- Willcox, J.K.; Ash, S.L.; Catignani, G.L. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef]
- Liu, G.; Yang, X.; Zhang, J.; Liang, L.; Miao, F.; Ji, T.; Ye, Z.; Chu, M.; Ren, J.; Xu, X. Synthesis, stability and anti-fatigue activity of selenium nanoparticles stabilized by Lycium barbarum polysaccharides. Int. J. Biol. Macromol. 2021, 179, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Wessely, S.; Chalder, T.; Hirsch, S.; Wallace, P.; Wright, D. The prevalence and morbidity of chronic fatigue and chronic fatigue syndrome: A prospective primary care study. Am. J. Public Health 1997, 87, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.S. Investigating unexplained fatigue in general practice with a particular focus on CFS/ME. BMC Fam. Pract. 2016, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- van’t Leven, M.; Zielhuis, G.A.; van der Meer, J.W.; Verbeek, A.L.; Bleijenberg, G. Fatigue and chronic fatigue syndrome-like complaints in the general population. Eur. J. Public Health 2010, 20, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Lv, J.; Lo, Y.M.; Cui, S.W.; Hu, X.; Fan, M. Effects of oat β-glucan on endurance exercise and its anti-fatigue properties in trained rats. Carbohyd. Polym. 2013, 92, 1159–1165. [Google Scholar] [CrossRef]
- Chen, L.; Song, L.; Li, T.; Zhu, J.; Xu, J.; Zheng, Q.; Yu, R. A New Antiproliferative and Antioxidant Peptide Isolated from Arca subcrenata. Mar. Drugs 2013, 11, 1800–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Ruiz, F.; Mancera-Andrade, E.I.; Iqbal, H.M. Marine-derived bioactive peptides for biomedical cectors: A review. Protein Pept. Lett. 2017, 24, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Khotimchenko, Y. Pharmacological Potential of Sea Cucumbers. Int. J. Mol. Sci. 2018, 19, 1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broekman, D.C.; Frei, D.M.; Gylfason, G.A.; Steinarsson, A.; Jörnvall, H.; Agerberth, B.; Gudmundsson, G.H.; Maier, V.H. Cod cathelicidin: Isolation of the mature peptide, cleavage site characterisation and developmental expression. Dev. Comp. Immunol. 2011, 35, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.R.; Zhang, L.; Ding, D.G.; Chi, C.F.; Wang, B.; Huo, J.C. Preparation, identification, and activity evaluation of eight antioxidant peptides from protein hydrolysate of Hairtail (Trichiurus japonicas) muscle. Mar. Drugs 2019, 17, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Mao, X.; Zhang, T.; Guo, X.; Ge, Y.; Ma, C.; Zhang, X. Separation and nanoencapsulation of antitumor peptides from Chinese three-striped box turtle (Cuora trifasciata). J. Microencapsul. 2016, 33, 344–354. [Google Scholar] [CrossRef]
- Xu, H. Studies on the antioxidant activity of soft-shelled turtle enzymatic hydrolysates. J. Chin. Inst. Food Sci. Technol. 2007, 4, 25–31. [Google Scholar]
- Wu, Y.C.; Liu, X.; Wang, J.L.; Chen, X.L.; Lei, L.; Han, J.; Jiang, Y.S.; Ling, Z.Q. Soft-shelled turtle peptide modulates microRNA profile in human gastric cancer AGS cells. Oncol. Lett. 2018, 15, 3109–3120. [Google Scholar] [CrossRef]
- Wang, P.; Zeng, H.; Lin, S.; Zhang, Z.; Zhang, Y.; Hu, J. Anti-fatigue activities of hairtail (Trichiurus lepturus) hydrolysate in an endurance swimming mice model. J. Funct. Foods 2020, 74, 104207. [Google Scholar] [CrossRef]
- Wang, X.; Xing, R.; Chen, Z.; Yu, H.; Li, R.; Li, P. Effect and mechanism of mackerel (Pneumatophorus japonicus) peptides for anti-fatigue. Food Funct. 2014, 5, 2113–2119. [Google Scholar] [CrossRef]
- Jin, H.M.; Wei, P. Anti-fatigue properties of tartary buckwheat extracts in mice. Int. J. Mol. Sci. 2011, 12, 4770–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Ren, J.W.; Zhang, T.; Liu, R.; Wu, L.; Du, Q.; Li, Y. Anti-fatigue effects of small-molecule oligopeptides isolated from Panax quinquefolium L. in mice. Food Funct. 2018, 9, 4266–4273. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.B.; Zhao, Y.Q.; Wang, Y.M.; Chi, C.F.; Wang, B. Eight collagen peptides from hydrolysate fraction of Spanish mackerel skins: Isolation, identification, and in vitro antioxidant activity evaluation. Mar. Drugs 2019, 17, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Wu, G.; Jiang, Y.; Li, B.; Feng, C.; Ge, Y.; Le, H.; Jiang, L.; Liu, H.; Shi, Y.; et al. Sea cucumber peptides improved the mitochondrial capacity of mice: A potential mechanism to enhance gluconeogenesis and fat catabolism during exercise for improved antifatigue property. Oxidative Med. Cell. Longev. 2020, 2020, 4604387. [Google Scholar] [CrossRef]
- Wang, Q.; Shi, J.; Zhong, H.; Zhuang, J.; Zhang, J.; Wang, J.; Zhang, X.; Feng, F. High-degree hydrolysis sea cucumber peptides improve exercise performance and exert antifatigue effect via activating the NRF2 and AMPK signaling pathways in mice. J. Funct. Foods 2021, 86, 104677. [Google Scholar] [CrossRef]
- Hu, M.; Du, J.; Du, L.; Luo, Q.; Xiong, J. Anti-fatigue activity of purified anthocyanins prepared from purple passion fruit (P. edulis Sim) epicarp in mice. J. Funct. Foods 2020, 65, 103725. [Google Scholar] [CrossRef]
- Zhang, X.; Jing, S.; Lin, H.; Sun, W.; Jiang, W.; Yu, C.; Sun, J.; Wang, C.; Chen, J.; Li, H. Anti-fatigue effect of anwulignan via the NRF2 and PGC-1α signaling pathway in mice. Food Funct. 2019, 10, 7755–7766. [Google Scholar] [CrossRef]
- Zhong, H.; Deng, L.; Zhao, M.; Tang, J.; Liu, T.; Zhang, H.; Feng, F. Probiotic-fermented blueberry juice prevents obesity and hyperglycemia in high fat diet-fed mice in association with modulating the gut microbiota. Food Funct. 2020, 11, 9192–9207. [Google Scholar] [CrossRef]
- Ørtenblad, N.; Westerblad, H.; Nielsen, J. Muscle glycogen stores and fatigue. J. Physiol. 2013, 591, 4405–4413. [Google Scholar] [CrossRef]
- Wang, J.; Yang, J.; Cao, M.; Zhao, Z.; Cao, B.; Yu, S. The potential roles of Nrf2/Keap1 signaling in anticancer drug interactions. Curr. Res. Pharmacol. Drug Discov. 2021, 2, 100028. [Google Scholar] [CrossRef]
- Cuadrado, A.; Rojo, A.I.; Wells, G.; Hayes, J.D.; Cousin, S.P.; Rumsey, W.L.; Attucks, O.C.; Franklin, S.; Levonen, A.-L.; Kensler, T.W. Therapeutic targeting of the NRF2 and KEAP1 partnership in chronic diseases. Nat. Rev. Drug Discov. 2019, 18, 295–317. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Wang, W.; Sadiq, F.A.; Wang, S.; Caiqin, L.; Jianchang, J. Involvement of Nrf2 and Keap1 in the activation of antioxidant responsive element (ARE) by chemopreventive agent peptides from soft-shelled turtle. Process Biochem. 2020, 92, 174–181. [Google Scholar] [CrossRef]
- Rabinowitz, J.D.; Enerbäck, S. Lactate: The ugly duckling of energy metabolism. Nat. Metab. 2020, 2, 566–571. [Google Scholar] [CrossRef]
- Villareal, M.O.; Matsukawa, T.; Isoda, H. L-Citrulline supplementation-increased skeletal muscle PGC-1α expression is associated with exercise performance and increased skeletal muscle weight. Mol. Nutr. Food Res. 2018, 62, 1701043. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Lin, D.; Guo, J.; Zhang, Y.; Zheng, B. In vitro antioxidant activity and in vivo anti-fatigue effect of sea horse (Hippocampus) peptides. Molecules 2017, 22, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef]
- Beyfuss, K.; Hood, D.A. A systematic review of p53 regulation of oxidative stress in skeletal muscle. Redox Rep. 2018, 23, 100–117. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Yang, Q.; Chen, X.; Tian, Y.; Liu, Z.; Wang, S. Bioactive peptides derived from crimson snapper and in vivo anti-aging effects on fat diet-induced high fat Drosophila melanogaster. Food Funct. 2020, 11, 524–533. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Liver (%) | Muscle (%) | Kidney (%) | Spleen (%) | Thymus (%) |
|---|---|---|---|---|---|
| Control | 5.57± 0.31 | 5.08 ± 0.93 | 1.51 ± 0.16 | 0.29 ±0.05 | 0.11 ± 0.03 |
| WP | 5.86 ± 0.50 | 5.32 ± 1.00 | 1.52 ± 0.14 | 0.31 ± 0.05 | 0.14 ± 0.04 |
| STP-L | 5.57 ± 0.57 | 5.30 ± 1.00 | 1.44 ± 0.29 | 0.32 ± 0.04 | 0.12 ± 0.05 |
| STP-M | 5.43 ± 0.25 | 5.14 ± 0.98 | 1.51 ± 0.07 | 0.27 ± 0.04 | 0.15 ± 0.06 |
| STP-H | 5.20 ± 0.81 | 5.65 ± 1.06 | 1.36 ± 0.16 | 0.30 ± 0.07 | 0.14 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, H.; Shi, J.; Zhang, J.; Wang, Q.; Zhang, Y.; Yu, P.; Guan, R.; Feng, F. Soft-Shelled Turtle Peptide Supplementation Modifies Energy Metabolism and Oxidative Stress, Enhances Exercise Endurance, and Decreases Physical Fatigue in Mice. Foods 2022, 11, 600. https://doi.org/10.3390/foods11040600
Zhong H, Shi J, Zhang J, Wang Q, Zhang Y, Yu P, Guan R, Feng F. Soft-Shelled Turtle Peptide Supplementation Modifies Energy Metabolism and Oxidative Stress, Enhances Exercise Endurance, and Decreases Physical Fatigue in Mice. Foods. 2022; 11(4):600. https://doi.org/10.3390/foods11040600
Chicago/Turabian StyleZhong, Hao, Jinyuan Shi, Junhui Zhang, Qianqian Wang, Yipeng Zhang, Peng Yu, Rongfa Guan, and Fengqin Feng. 2022. "Soft-Shelled Turtle Peptide Supplementation Modifies Energy Metabolism and Oxidative Stress, Enhances Exercise Endurance, and Decreases Physical Fatigue in Mice" Foods 11, no. 4: 600. https://doi.org/10.3390/foods11040600