
Investigation on Cell Disruption Techniques and Supercritical Carbon Dioxide Extraction of Mortierella alpina Lipid


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Microorganism Cultivation
2.2. Cell Disruption—Assisted Extraction Techniques
2.2.1. High-Speed Homogenizer
2.2.2. Acid Treatment
2.2.3. Ultrasonication
2.2.4. High-Pressure Processing (HPP)
2.3. Supercritical Carbon Dioxide (SC-CO2) Extraction
2.4. Lipid Recovery Calculation
2.5. Fatty Acid Composition Analysis
2.6. Scanning Electron Microscopy
2.7. Statistical Analysis
3. Results and Discussion
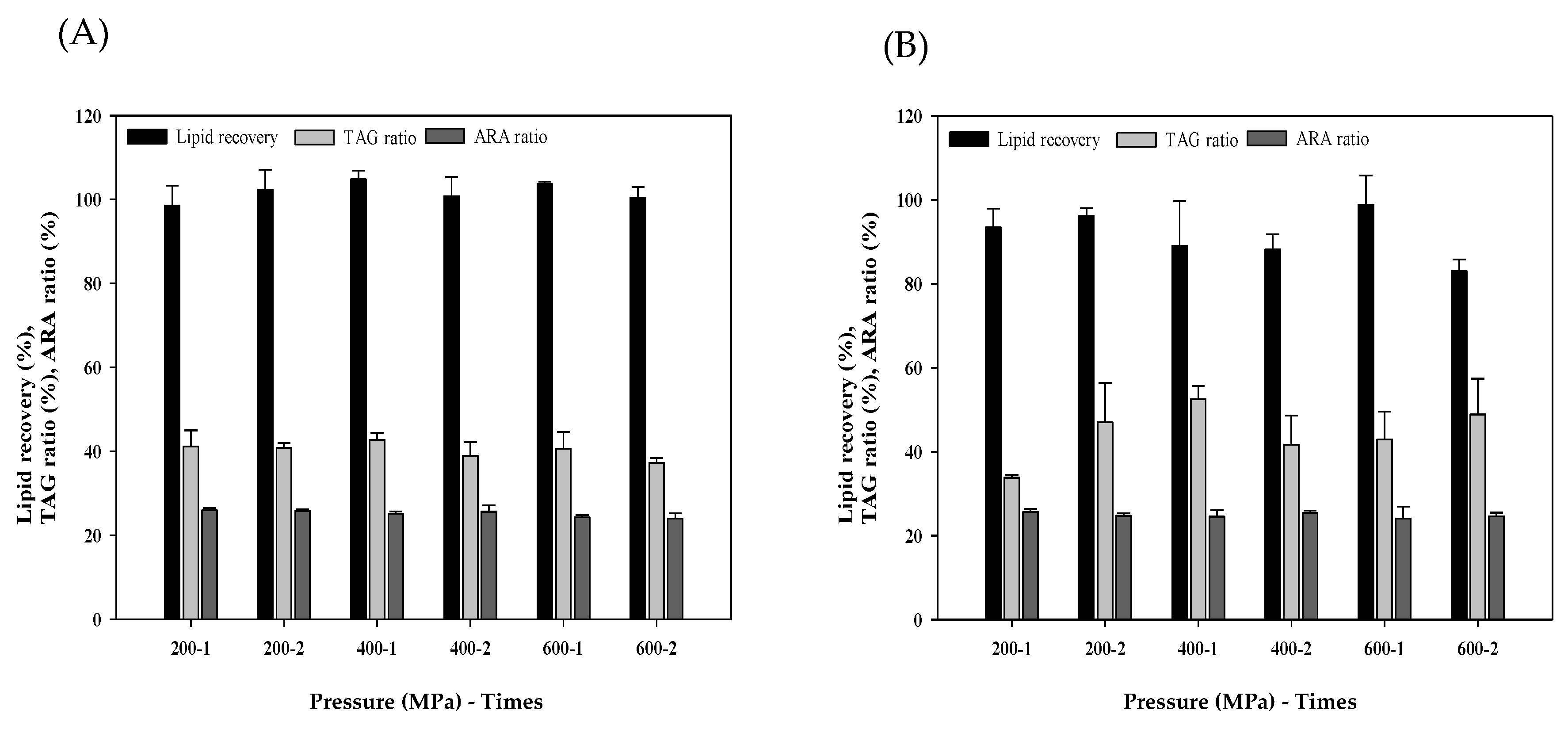
3.1. Lipid Recovery of Different Cell Disruption
3.2. SEM Image of M. alpina after Different Cell Disruption
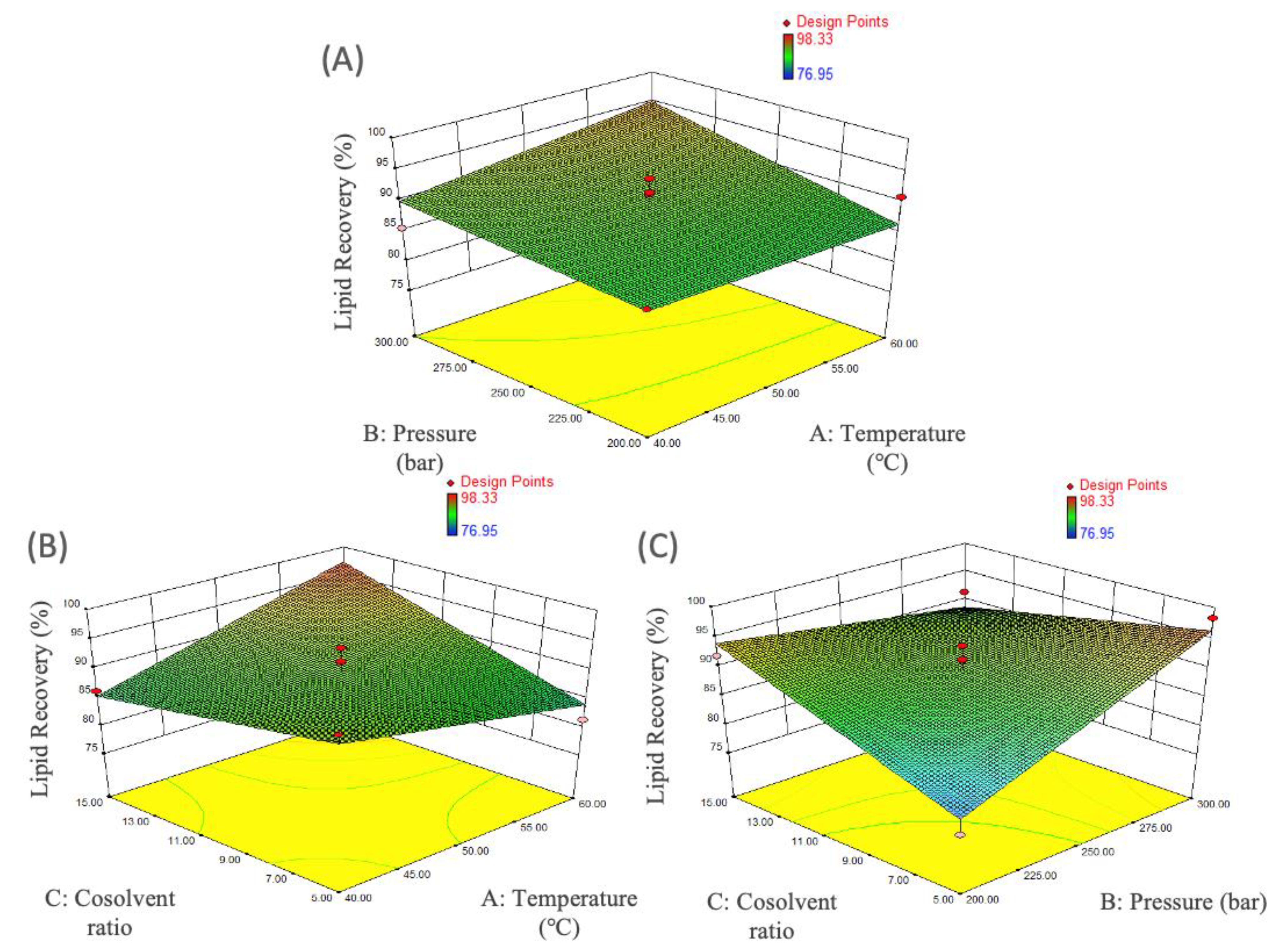
3.3. Supercritical Carbon Dioxides (SC-CO2) Extraction of M. alpina Lipid
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ward, O.P.; Singh, A. Omega-3/6 fatty acids: Alternative sources of production. Proc. Biochem. 2005, 40, 3627–3652. [Google Scholar] [CrossRef]
- Zhang, H.; Cui, Q.; Song, X. Research advances on arachidonic acid production by fermentation and genetic modification of Mortierella alpina. World J. Microbiol. Biotechnol. 2021, 37, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lu, D.; Li, X.; Feng, Y.; Cui, Q.; Song, X. Heavy ion mutagenesis combined with triclosan screening provides a new strategy for improving the arachidonic acid yield in Mortierella alpina. BMC Biotechnol. 2018, 18, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asadi, S.Z.; Khosravi-Darani, K.; Nikoopour, H.; Bakhoda, H. Production of arachidonic acid and eicosapentaenoic acid by Mortierella alpina CBS 528.72 on date waste. Food Technol. Biotechnol. 2018, 56, 197–207. [Google Scholar] [CrossRef]
- Needleman, P.; Jakschik, B.; Morrison, A.; Lefkowith, J. Arachidonic acid metabolism. Ann. Rev. Biochem. 1986, 55, 69–102. [Google Scholar] [CrossRef]
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for single cell oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef]
- Halim, R.; Danquah, M.K.; Webley, P.A. Extraction of oil from microalgae for biodiesel production: A review. Biotechnol. Adv. 2012, 30, 709–732. [Google Scholar] [CrossRef]
- Monks, L.M.; Rigo, A.; Mazutti, M.A.; Vladimir Oliveira, J.; Valduga, E. Use of chemical, enzymatic and ultrasound-assisted methods for cell disruption to obtain carotenoids. Biocatal. Agric. Biotechnol. 2013, 2, 165–169. [Google Scholar] [CrossRef]
- Machado, F.R.S.; Trevisol, T.C.; Boschetto, D.L.; Burkert, J.F.M.; Ferreira, S.R.S.; Oliveira, J.V.; Burkert, C.A.V. Technological process for cell disruption, extraction and encapsulation of astaxanthin from Haematococcus pluvialis. J. Biotechnol. 2016, 218 (Suppl. C), 108–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza Silva, A.P.F.; Costa, M.C.; Lopes, A.C.; Neto, E.F.A.; Leitão, R.C.; Mota, C.R.; dos Santos, A.B. Comparison of pretreatment methods for total lipids extraction from mixed microalgae. Renew. Energy 2014, 63, 762–766. [Google Scholar] [CrossRef]
- Yu, X.; Dong, T.; Zheng, Y.; Miao, C.; Chen, S. Investigations on cell disruption of oleaginous microorganisms: Hydrochloric acid digestion is an effective method for lipid extraction. Eur. J. Lipid Sci. Technol. 2015, 117, 730–737. [Google Scholar] [CrossRef]
- Duarte, S.H.; dos Santos, P.; Michelon, M.; de Pinho Oliveira, S.M.; Martínez, J.; Maugeri, F. Recovery of yeast lipids using different cell disruption techniques and supercritical CO2 extraction. Biochem. Eng. J. 2017, 125, 230–237. [Google Scholar] [CrossRef]
- Nisha, A.; Sankar, K.U.; Venkateswaran, G. Supercritical CO2 extraction of Mortierella alpina single cell oil: Comparison with organic solvent extraction. Food Chem. 2012, 133, 220–226. [Google Scholar] [CrossRef]
- You, J.-Y.; Peng, C.; Liu, X.; Ji, X.-J.; Lu, J.; Tong, Q.; Wei, P.; Cong, L.; Li, Z.; Huang, H. Enzymatic hydrolysis and extraction of arachidonic acid rich lipids from Mortierella alpina. Bioresour. Technol. 2011, 102, 6088–6094. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zhou, P.P.; Yu, L.J. Extraction of lipids from Mortierella alpina and enrichment of arachidonic acid from the fungal lipids. Bioresour. Technol. 2002, 84, 93–95. [Google Scholar] [CrossRef]
- Reboleira, J.; Félix, R.; Vicente, T.F.L.; Januário, A.P.; Félix, C.; de Melo, M.M.R.; Silva, C.M.; Ribeiro, A.C.; Saraiva, J.A.; Bandarra, N.M.; et al. Uncovering the bioactivity of Aurantiochytrium sp.: A comparison of extraction methodologies. J. Mar. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Gänzle, M.; Liu, Y. Mechanisms of pressure-mediated cell death and injury in Escherichia coli: From fundamentals to food applications. Front. Microbiol. 2015, 6, 599. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Calvo, M.; Prieto, M.; Lopez, M.; Alvarez-Ordonez, A. Effects of high hydrostatic pressure on Escherichia coli ultrastructure, membrane integrity and molecular composition as assessed by FTIR spectroscopy and microscopic imaging techniques. Molecules 2014, 19, 21310–21323. [Google Scholar] [CrossRef]
- Muntean, M.-V.; Marian, O.; Barbieru, V.; Cătunescu, G.M.; Ranta, O.; Drocas, I.; Terhes, S. High pressure processing in food industry—Characteristics and applications. Agric. Agric. Sci. Procedia 2016, 10, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Dong, M.; Walker, T.H. Characterization of high-pressure carbon dioxide explosion to enhance oil extraction from canola. J. Supercrit. Fluids 2008, 44, 193–200. [Google Scholar] [CrossRef]
- Machmudah, S.; Sulaswatty, A.; Sasaki, M.; Goto, M.; Hirose, T. Supercritical CO2 extraction of nutmeg oil: Experiments and modeling. J. Supercrit. Fluids 2006, 39, 30–39. [Google Scholar] [CrossRef]
- Sahena, F.; Zaidul, I.S.M.; Jinap, S.; Karim, A.A.; Abbas, K.A.; Norulaini, N.A.N.; Omar, A.K.M. Application of supercritical CO2 in lipid extraction—A review. J. Food Eng. 2009, 95, 240–253. [Google Scholar] [CrossRef]
- Dunford, N.T.; Zhang, M. Pressurized solvent extraction of wheat germ oil. Food Res. Int. 2003, 36, 905–909. [Google Scholar] [CrossRef]
- Dong, M.; Walker, T.H. Addition of polyunsaturated fatty acids to canola oil by fungal conversion. Enzym. Microb. Technol. 2008, 42, 514–520. [Google Scholar] [CrossRef]
- Wang, D.; Li, Y.; Hu, X.; Su, W.; Zhong, M. Combined enzymatic and mechanical cell disruption and lipid extraction of green alga Neochloris oleoabundans. Int. J. Mol. Sci. 2015, 16, 7707–7722. [Google Scholar] [CrossRef] [PubMed]
- Messens, W.; Van Camp, J.; Huyghebaert, A. The use of high pressure to modify the functionality of food proteins. Trends Food Sci. Technol. 1997, 8, 107–112. [Google Scholar] [CrossRef]
- Mat Yusoff, M.; Gordon, M.H.; Ezeh, O.; Niranjan, K. High pressure pre-treatment of Moringa oleifera seed kernels prior to aqueous enzymatic oil extraction. Innov. Food Sci. Emerg. Technol. 2017, 39, 129–136. [Google Scholar] [CrossRef]
- Liu, S.; Yang, F.; Zhang, C.; Ji, H.; Hong, P.; Deng, C. Optimization of process parameters for supercritical carbon dioxide extraction of Passiflora seed oil by response surface methodology. J. Supercrit. Fluids 2009, 48, 9–14. [Google Scholar] [CrossRef]
- Couto, R.M.; Simões, P.C.; Reis, A.; Silva, T.L.D.; Martins, V.H.; Sánchez-Vicente, Y. Supercritical fluid extraction of lipids from the heterotrophic microalga Crypthecodinium cohnii. Eng. Life Sci. 2010, 10, 158–164. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Lipid Recovery (%) | ARA Ratio (%) |
|---|---|---|
| High-speed homogenizer | 115.40 ± 3.47 a | 31.72 ± 0.65 a |
| Acid disruption (HCl) | ||
| 1 M | 115.55 ± 3.88 a | 29.03 ± 0.19 b |
| 2 M | 112.95 ± 0.31 a | 29.21 ± 0.18 b |
| 3 M | 107.36 ± 1.47 b | 28.23 ± 0.19 c |
| Ultrasonication | 101.73 ± 4.44 b | 28.91 ± 0.55 bc |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value Prob > F |
|---|---|---|---|---|---|
| Model | 322.36 | 6 | 53.73 | 4.89 | 0.0138 |
| A: Temperature | 9.10 | 1 | 9.10 | 0.83 | 0.3842 |
| B: Pressure | 64.24 | 1 | 64.24 | 5.85 | 0.0361 |
| C: Cosolvent ratio | 22.85 | 1 | 22.85 | 2.08 | 0.1797 |
| AB | 8.12 | 1 | 8.12 | 0.74 | 0.4099 |
| AC | 96.73 | 1 | 96.73 | 8.81 | 0.0141 |
| BC | 121.33 | 1 | 121.33 | 11.05 | 0.0077 |
| Residual | 109.80 | 10 | 10.98 | ||
| Lack of fit | 84.36 | 6 | 14.06 | 2.21 | 0.2313 |
| Pure error | 25.45 | 4 | 6.36 | ||
| Cor total | 432.17 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ong, C.C.; Chen, Y.-H. Investigation on Cell Disruption Techniques and Supercritical Carbon Dioxide Extraction of Mortierella alpina Lipid. Foods 2022, 11, 582. https://doi.org/10.3390/foods11040582
Ong CC, Chen Y-H. Investigation on Cell Disruption Techniques and Supercritical Carbon Dioxide Extraction of Mortierella alpina Lipid. Foods. 2022; 11(4):582. https://doi.org/10.3390/foods11040582
Chicago/Turabian StyleOng, Chang Chng, and Yen-Hui Chen. 2022. "Investigation on Cell Disruption Techniques and Supercritical Carbon Dioxide Extraction of Mortierella alpina Lipid" Foods 11, no. 4: 582. https://doi.org/10.3390/foods11040582