
Impact of Enzymatic Hydrolysis and Heat Inactivation on the Physicochemical Properties of Milk Protein Hydrolysates


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
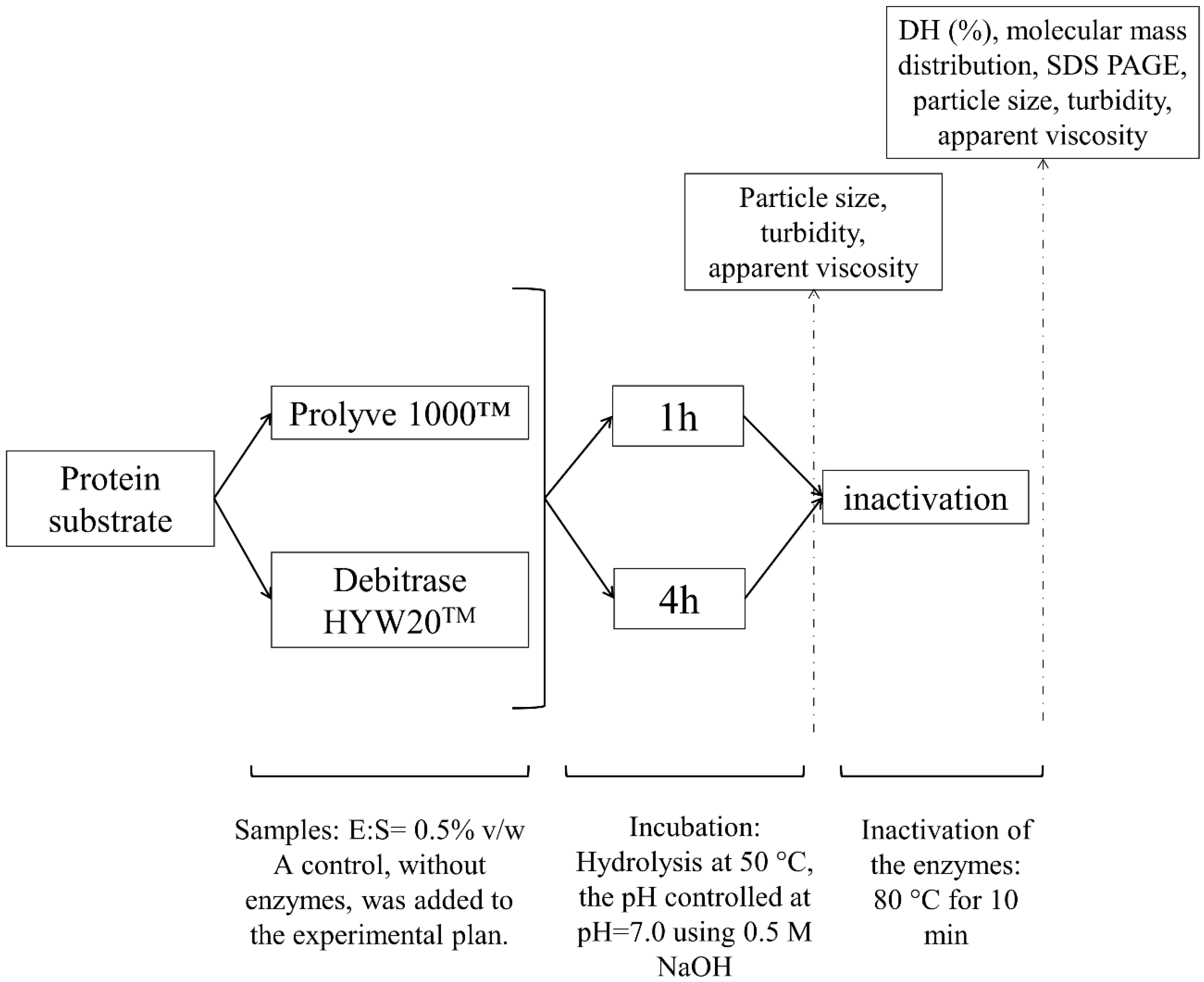
2.2.1. Generation of Enzymatic Hydrolysates
2.2.2. Determination of DH
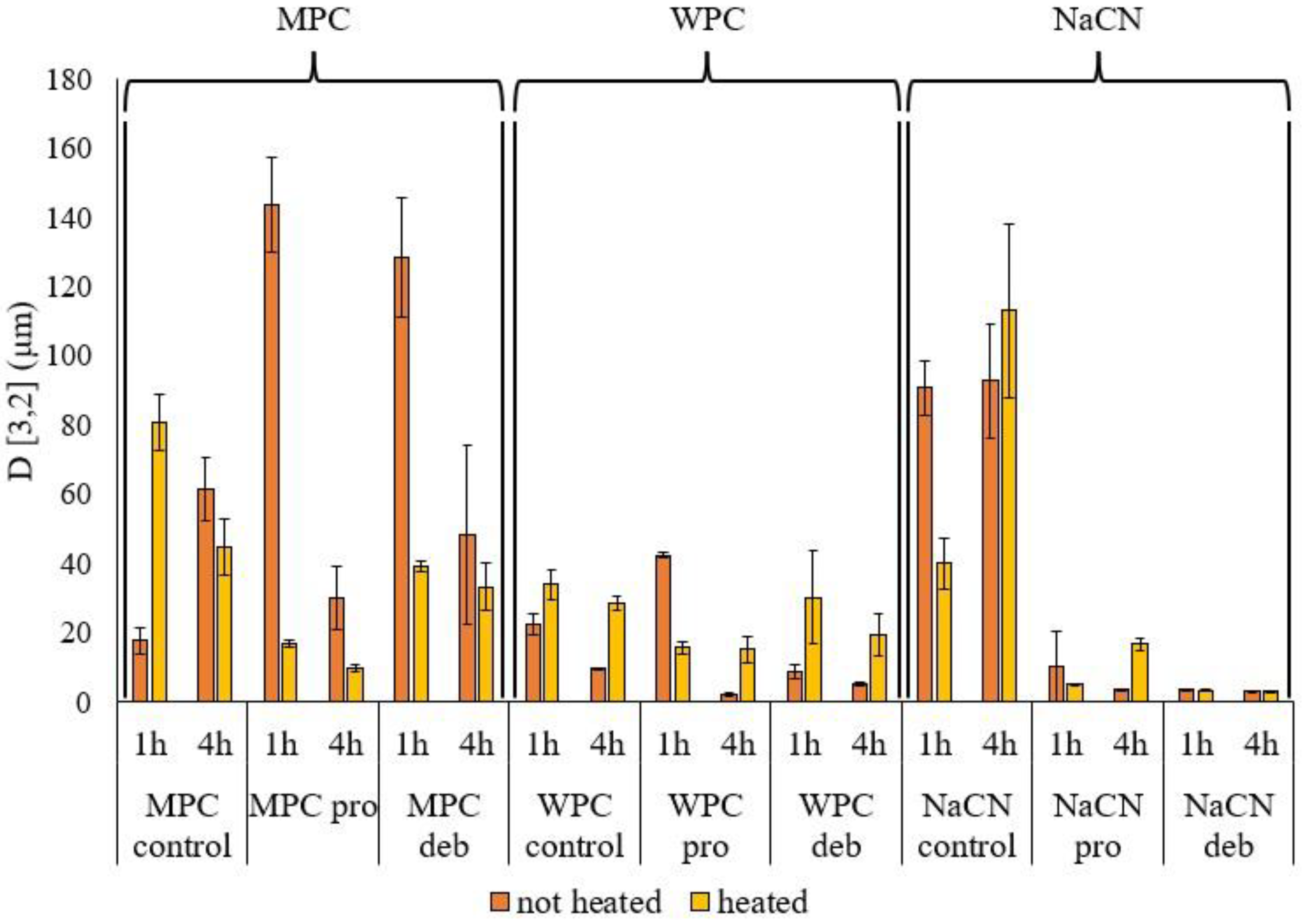
2.2.3. Particle Size Distribution
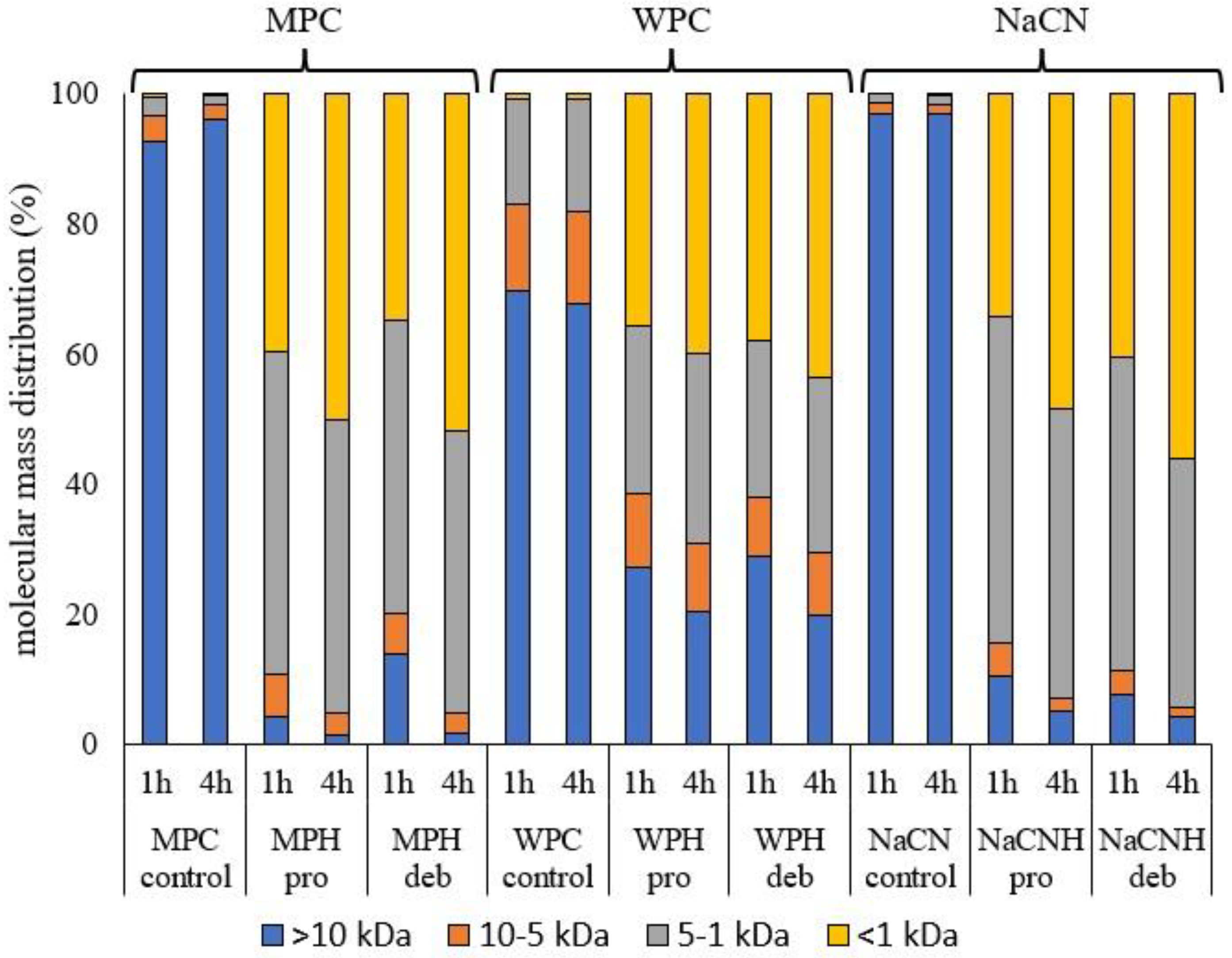
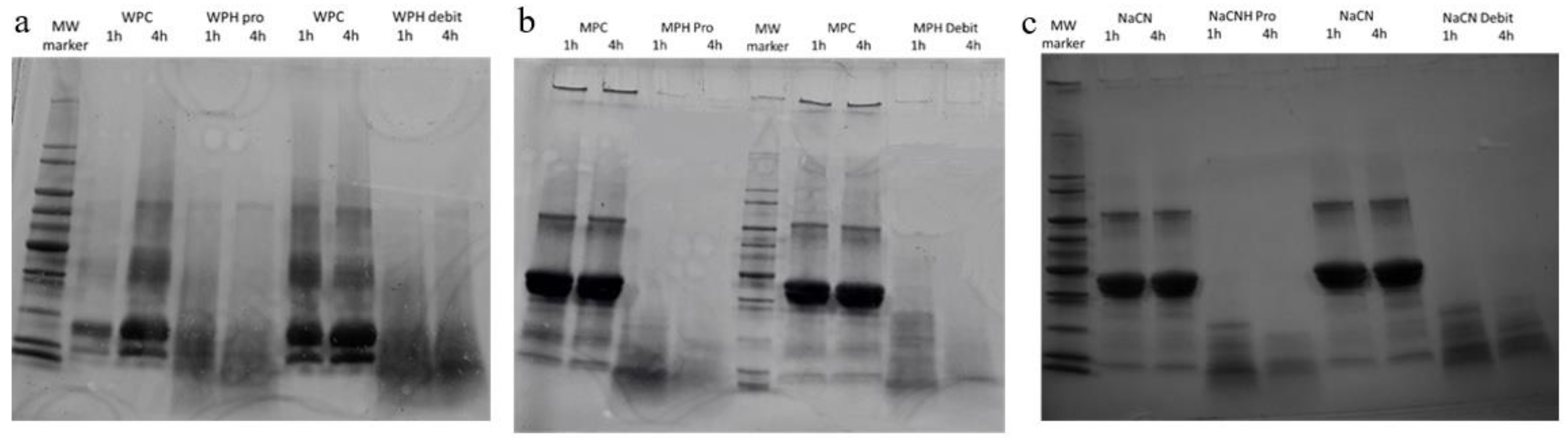
2.2.4. Molecular mass Distribution
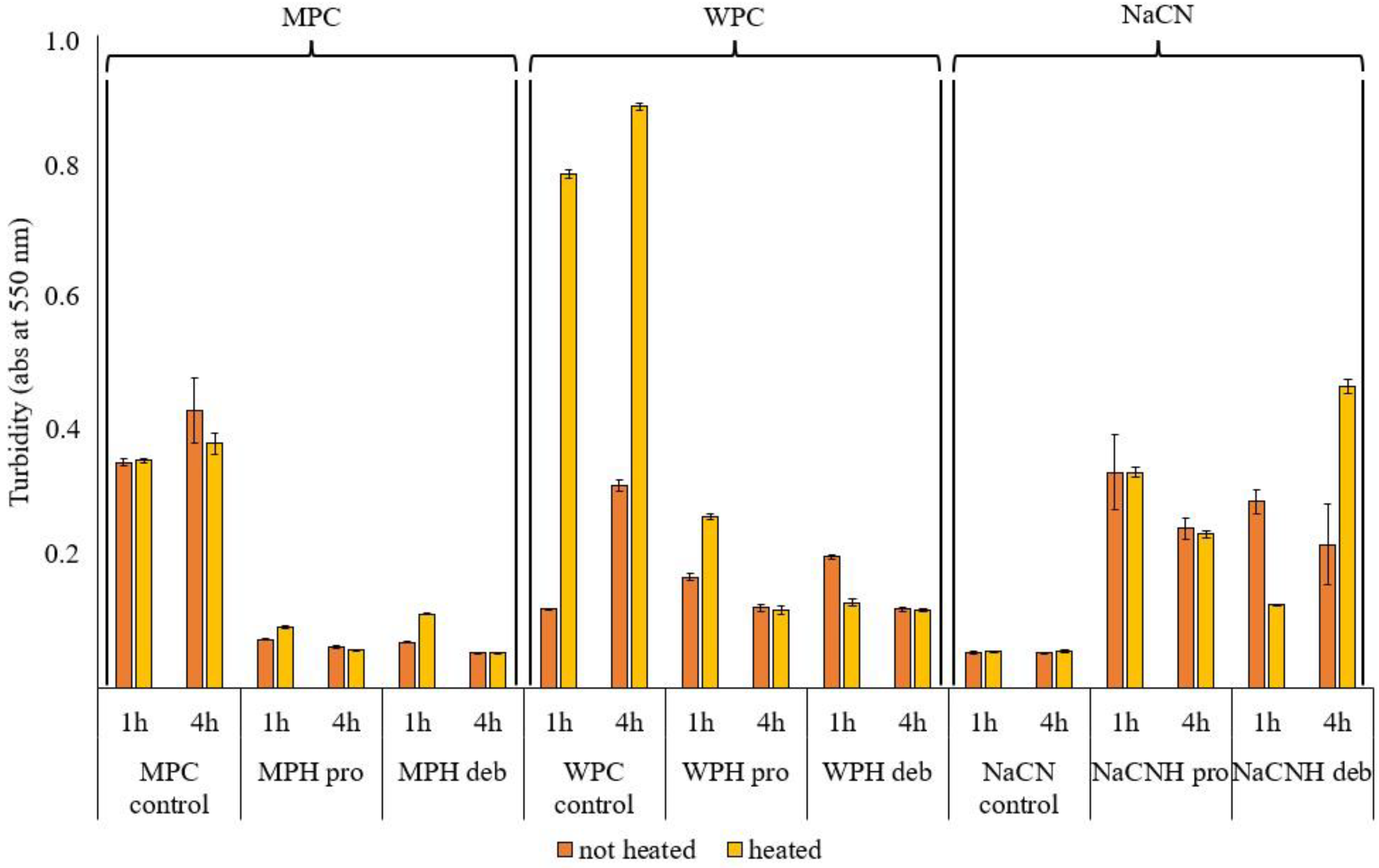
2.2.5. Turbidity (A550nm) Measurements
2.2.6. Determination of Apparent Viscosity (ηapp)
2.2.7. Statistical Analysis
3. Results and Discussion
3.1. Hydrolysis of Milk Protein Substrates
3.2. Physicochemical Characteristics of the Hydrolysates
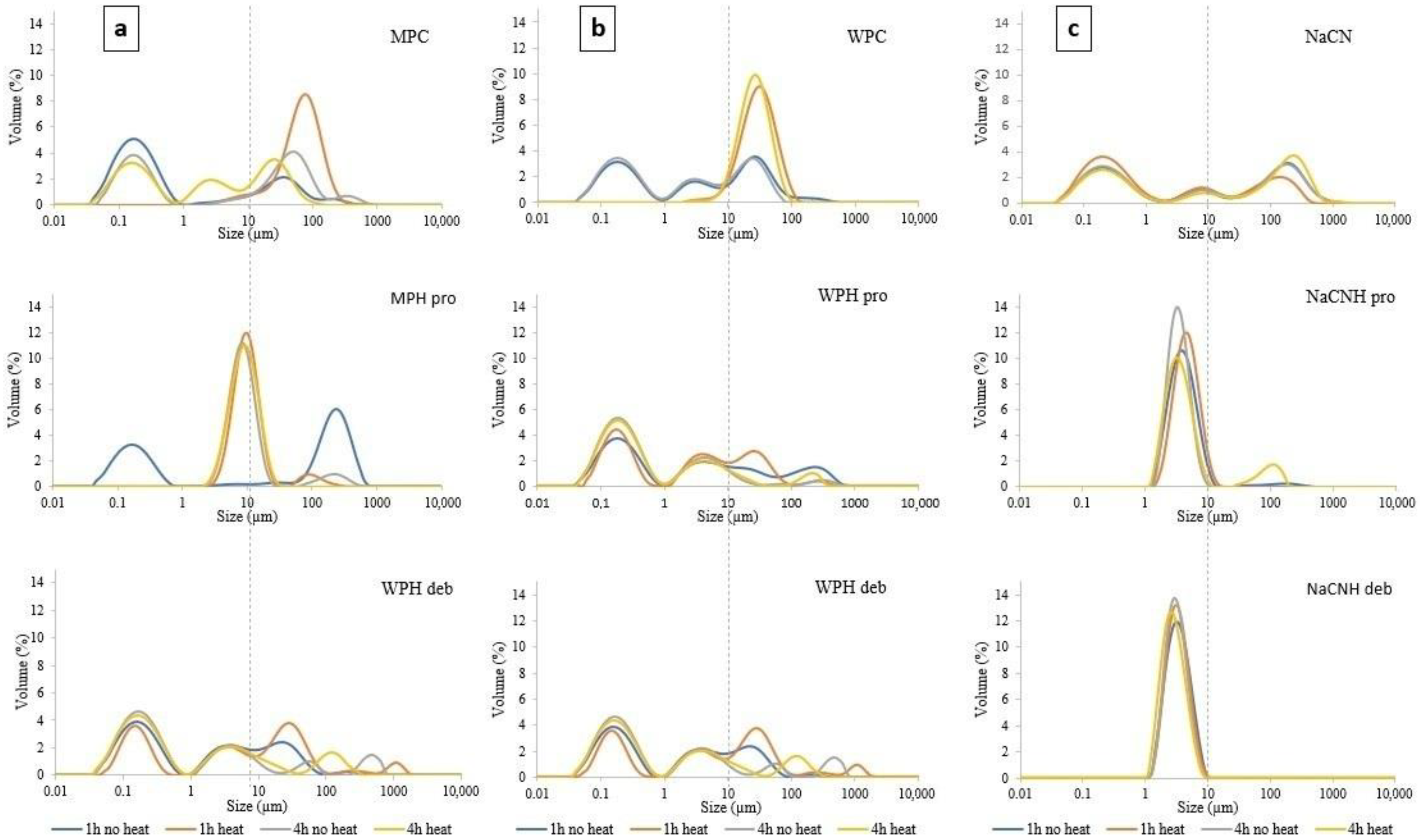
3.2.1. Particle Size
3.2.2. Turbidity
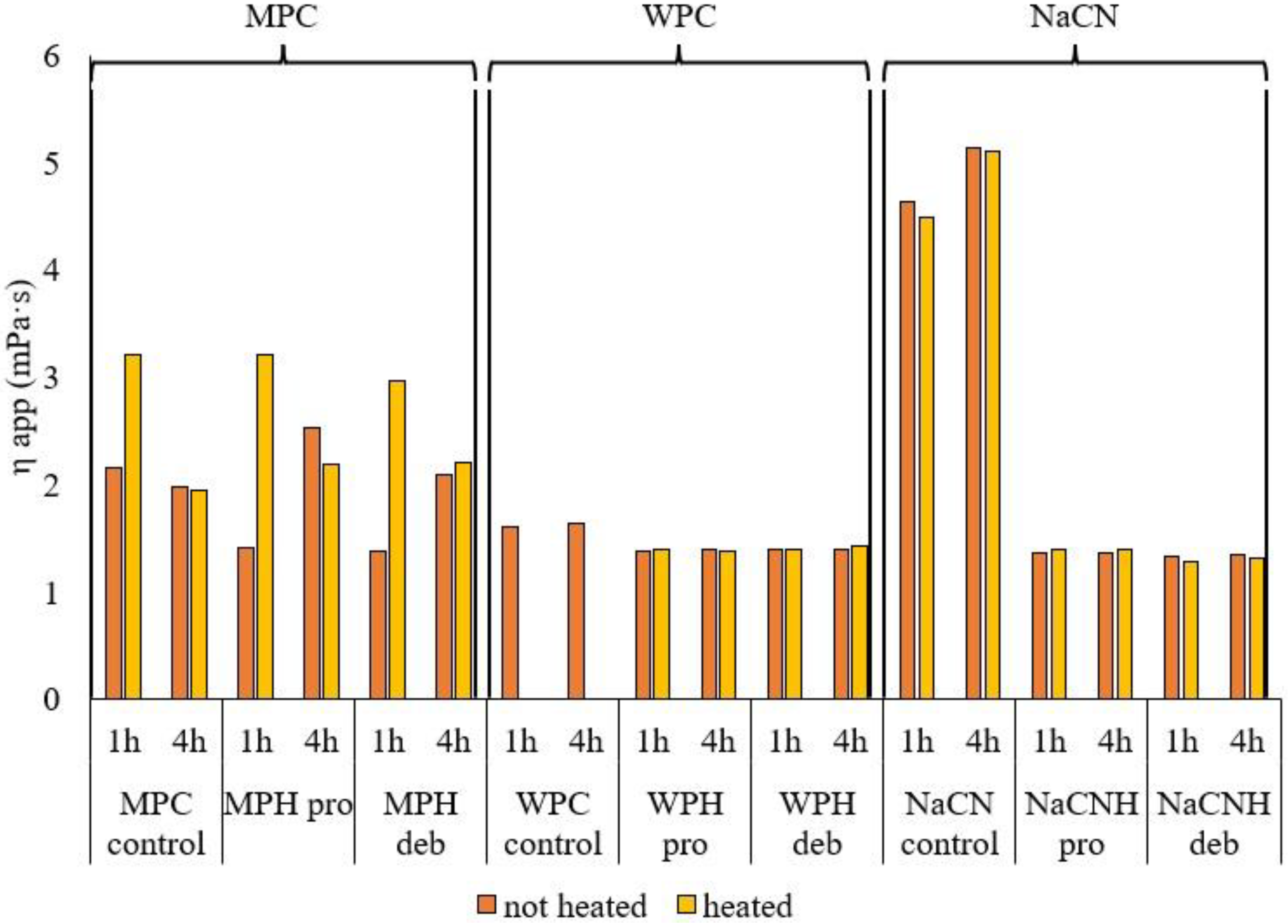
3.2.3. Apparent Viscosity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Carr, A.; Golding, M. Functional milk proteins production and utilization: Casein-based ingredients. In Advanced Dairy Chemistry, 4th ed.; McSweeney, P.L.H., O’Mahony, J.A., Eds.; Springer: Cork, Ireland, 2016; Volume 1B, pp. 35–66. [Google Scholar]
- Nongonierma, A.B.; FitzGerald, R.J. The scientific evidence for the role of milk protein-derived bioactive peptides in humans: A Review. J. Funct. Foods 2015, 17, 640–656. [Google Scholar] [CrossRef] [Green Version]
- Cheison, S.C.; Kulozik, U. Impact of the environmental conditions and substrate pre-treatment on whey protein hydrolysis: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 418–453. [Google Scholar] [CrossRef]
- Abd El-Salam, M.H.; El-Shibiny, S. Preparation, properties, and uses of enzymatic milk protein hydrolysates. Crit. Rev. Food Sci. Nutr. 2017, 57, 1119–1132. [Google Scholar] [CrossRef]
- Izadi, A.; Khedmat, L.; Mojtahedi, S.Y. Nutritional and therapeutic perspectives of camel milk and its protein hydrolysates: A review on versatile biofunctional properties. J. Funct. Foods 2019, 60, 103441. [Google Scholar] [CrossRef]
- Mulvihill, D.M. Production, functional properties and utilization of milk protein products. In Advanced Dairy Chemistry; Fox, P.F., Ed.; Elsevier Science Publishers: London, UK, 1992; Volume 1, pp. 369–404. [Google Scholar] [CrossRef]
- Spellman, D.; McEvoy, E.; O’Cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Pelegrine, D.H.G.; Gasparetto, C.A. Whey proteins solubility as function of temperature and pH. LWT-Food Sci. Technol. 2005, 38, 77–80. [Google Scholar] [CrossRef]
- Agarwal, S.; Beausire, R.L.; Patel, S.; Patel, H. Innovative uses of milk protein concentrates in product development. J. Food Sci. 2015, 80, A23–A29. [Google Scholar] [CrossRef]
- De Castro-Morel, M.; Harper, W.J. Basic functionality of commercial milk protein concentrates. Milchwissenschaft 2002, 57, 367–370. [Google Scholar]
- Meena, G.S.; Singh, A.K.; Panjagari, N.R.; Arora, S. Milk protein concentrates: Opportunities and challenges. J. Food Sci. Technol. 2017, 54, 3010–3024. [Google Scholar] [CrossRef]
- Khalesi, M.; FitzGerald, R.J. Insolubility in milk protein concentrates: Potential causes and strategies to minimize its occurrence. Crit. Rev. Food Sci. Nutr. 2021, 15, 1–17. [Google Scholar] [CrossRef]
- Panyam, D.; Kilara, A. Enhancing the functionality of food proteins by enzymatic modification. Trends Food Sci. Technol. 1996, 71, 120–125. [Google Scholar] [CrossRef]
- Flanagan, J.; FitzGerald, R.J. Functionality of Bacillus proteinase hydrolysates of sodium caseinate. Int. Dairy J. 2002, 12, 737–748. [Google Scholar] [CrossRef]
- Tuncturk, Y.; Zorba, O. The effects of enzymatic hydrolysis of casein on apparent yield stress and some emulsion properties. Food Hydrocoll. 2006, 20, 475–482. [Google Scholar] [CrossRef]
- De Castro, R.J.S.; Bagagli, M.P.; Sato, H.H. Improving the functional properties of milk proteins: Focus on the specificities of proteolytic enzymes. Curr. Opin. Food Sci. 2015, 1, 64–69. [Google Scholar] [CrossRef]
- Cui, Q.; Sun, Y.; Zhou, Z.; Cheng, J.; Guo, M. Effects of enzymatic hydrolysis on physicochemical properties and solubility and bitterness of milk protein hydrolysates. Foods 2021, 10, 2462. [Google Scholar] [CrossRef]
- Ryan, G.; Nongonierma, A.B.; O’Regan, J.; FitzGerald, R.J. Functional properties of bovine milk protein isolate and associated enzymatic hydrolysates. Int. Dairy J. 2018, 81, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Turgeon, S.L.; Gauthier, S.F.; Paquin, P. Interfacial and Emulsifying Properties of Whey Peptide Fractions Obtained with a Two-step Ultrafiltration Process. J. Agric. Food Chem. 1991, 39, 70–73. [Google Scholar] [CrossRef]
- Mutilangi, W.A.M.; Panyam, D.; Kilara, A. Functional Properties of Hydrolysates from Proteolysis of Heat-denatured Whey Protein Isolate. J. Food Sci. 1996, 61, 270–274. [Google Scholar] [CrossRef]
- Severin, S.; Xia, W.S. Enzymatic hydrolysis of whey proteins by two different proteases and their effect on the functional properties of resulting protein hydrolysates. J. Food Biochem. 2006, 30, 77–97. [Google Scholar] [CrossRef]
- Cermeño, M.; Felix, M.; Connolly, A.; Brennan, E.; Coffey, B.; Ryan, E.; FitzGerald, R.J. Role of carbohydrate conjugation on the emulsification and antioxidant properties of intact and hydrolysed whey protein concentrate. Food Hydrocoll. 2019, 88, 170–179. [Google Scholar] [CrossRef] [Green Version]
- Walsh, D.J.; Russell, K.; FitzGerald, R.J. Stabilisation of sodium caseinate hydrolysate foams. Food Res. Int. 2008, 41, 43–52. [Google Scholar] [CrossRef]
- Banach, J.C.; Lin, Z.; Lamsal, B.P. Enzymatic modification of milk protein concentrate and characterization of resulting functional properties. LWT-Food Sci. Technol. 2013, 54, 397–403. [Google Scholar] [CrossRef]
- McIntyre, I.; Carolan, A.; O’Sullivan, M.; Jacquier, J.; Hutchings, S.; Murray, B.; O’Riordan, D. Incorporation of bioactive dairy hydrolysate influences the stability and digestion behaviour of milk protein stabilised emulsions. Food Funct. 2018, 9, 5813–5823. [Google Scholar] [CrossRef]
- Chobert, J.; Bertrand-Harb, C.J.; Nicolas, M. Solubility and Emulsifying Properties of Caseins and Whey Proteins Modified Enzymatically by Trypsin. J. Agric. Food Chem. 1988, 36, 883–892. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, N.; Ashaolu, T.J. Whey proteins and peptides in health-promoting functions—A review. Int. Dairy J. 2021, 126, 105269. [Google Scholar] [CrossRef]
- Le Maux, S.; Nongonierma, A.B.; Barre, C.; FitzGerald, R.J. Enzymatic generation of whey protein hydrolysates under pH-controlled and non pH-controlled conditions: Impact on physicochemical and bioactive properties. Food Chem. 2016, 199, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dermiki, M.; FitzGerald, R.J. Physicochemical and gelling properties of whey protein hydrolysates generated at 5 and 50 °C using Alcalase® and Neutrase®, effect of total solids and incubation time. Int. Dairy J. 2020, 110, 104792. [Google Scholar] [CrossRef]
- Le Maux, S.; Nongonierma, A.B.; FitzGerald, R.J. Peptide composition and dipeptidyl peptidase IV inhibitory properties of β-lactoglobulin hydrolysates having similar extents of hydrolysis while generated using different enzyme-to-substrate ratios. Food Res. Int. 2017, 99, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Spellman, D.; Kenny, P.; O’Cuinn, G.; FitzGerald, R.J. Aggregation properties of whey protein hydrolysates generated with Bacillus licheniformis proteinase activities. J. Agric. Food Chem. 2005, 53, 1258–1265. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Control of the Proteolytic Reaction and of the Level of Bitterness in Protein Hydrolysis Processes. J. Chem. Technol. Biotechnol. 1984, 34, 215–222. [Google Scholar] [CrossRef]
- Rajarathnam, E.; Nongonierma, A.B.; O’Sullivan, D.; Flynn, C.; FitzGerald, R.J. Impact of enzyme preparation and degree of hydrolysis on peptide profile and nitrogen solubility of sodium caseinate hydrolysates. Int. J. Food Sci. Technol. 2016, 51, 2123–2131. [Google Scholar] [CrossRef] [Green Version]
- Le Maux, S.; Nongonierma, A.B.; Lardeux, C.; FitzGerald, R.J. Impact of enzyme inactivation conditions during the generation of whey protein hydrolysates on their physicochemical and bioactive properties. Int. J. Food Sci. Technol. 2018, 53, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; FitzGerald, R.J. Tryptophan-containing milk protein-derived dipeptides inhibit xanthine oxidase. Peptides 2012, 37, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Loughlin, I.B.; Murray, B.A.; Kelly, P.M.; FitzGerald, R.J.; Brodkorb, A. Enzymatic Hydrolysis of Heat-Induced Aggregates of Whey Protein Isolate. J. Agric. Food Chem. 2012, 60, 4895–4904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addinsoft. XLSTAT Statistical and Data Analysis Solution 2020. New York, NY, USA. Available online: https://www.xlstat.com (accessed on 16 May 2020).
- Spellman, D.; O’Cuinn, G.; FitzGerald, R.J. Physicochemical and sensory characteristics of whey protein hydrolysates generated at different total solids levels. J. Dairy Res. 2005, 72, 138–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FitzGerald, R.J.; O’Cuinn, G. Enzymatic debittering of food protein hydrolysates. Biotechnol. Adv. 2006, 24, 234–237. [Google Scholar] [CrossRef]
- Whitehurst, R.J.; Van Oort, M. Enzymes in Food Technology, 2nd ed.; Wiley Blackwell: Singapore, 2009. [Google Scholar]
- Power, O.; Jakeman, P.; FitzGerald, R.J. Antioxidative peptides: Enzymatic production, in vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids 2013, 44, 797–820. [Google Scholar] [CrossRef]
- Svendsen, I.; Breddam, K. Isolation and amino acid sequence of a glutamic acid specific endopeptidase from Bacillus licheniformis. Eur. J. Biochem. 1992, 204, 165–171. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Enzymes exogenous to milk in dairy technology. Proteinases. In Encyclopedia of Dairy Science, 2nd ed.; Fuquay, J.W., Fox, P.F., Mc Sweeney, P.J.H., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 289–296. [Google Scholar]
- McDonagh, D.; FitzGerald, R.J. Production of caseinophosphopeptides (CPPs) from sodium caseinate using a range of commercial protease preparations. Int. Dairy J. 1998, 8, 39–45. [Google Scholar] [CrossRef]
- Bhandari, D.; Rafiq, S.; Gat, Y.; Gat, P.; Waghmare, R.; Kumar, V. A review on bioactive peptides: Physiological functions, bioavailability and safety. Int. J. Pept. Res. Ther. 2020, 26, 139–150. [Google Scholar] [CrossRef]
- Adjonu, R.; Doran, G.; Torley, P.; Agboola, S. Whey protein peptides as components of nanoemulsions: A review of emulsifying and biological functionalities. J. Food Eng. 2014, 122, 15–27. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, L.; Shi, Y.; Akhtar, M.; Chen, J.; Ettelaie, R. Emulsifying and emulsion stabilizing properties of soy protein hydrolysates, covalently bonded to polysaccharides: The impact of enzyme choice and the degree of hydrolysis. Food Hydrocoll. 2021, 113, 106519. [Google Scholar] [CrossRef]
- Karami, Z.; Akbari-Adergani, B. Bioactive food derived peptides: A review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Morais, H.A.; Silvestre, M.P.C.; Viviane, D.M.; Mauro, R.S.; Silva, A.S.; Silvera, J.N. Correlation between the degree of hydrolysis and the peptide profile of whey protein of hydrolysis and peptide profile of whey protein concentrate hydrolysate: Effect of the enzyme type and reaction time. Am. J. Food Technol. 2013, 8, 1–16. [Google Scholar] [CrossRef]
- Gani, A.; Broadway, A.A.; Ahmad, M.; Ashwar, B.A.; Wani, A.A. Enzymatic hydrolysis of whey and casein protein- effect on functional, rheological, textural and sensory properties of breads. J. Food Sci. Technol. 2015, 52, 7697–7709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahler, H.; Friess, W.; Grauschopf, U.; Kiese, S. Protein Aggregation: Pathways, Induction Factors and Analysis. J. Pharm. Sci. 2008, 98, 2909–2934. [Google Scholar] [CrossRef]
- Ewert, J.; Luz, A.; Volk, V.; Stressler, T.; Fischer, L. Enzymatic production of emulsifying whey protein hydrolysates without the need of heat inactivation. J. Sci. Food Agric. 2019, 99, 3443–3450. [Google Scholar] [CrossRef]
- Iametti, S.; De Gregori, B.; Vecchio, G.; Bonomi, F. Modifications occur at different structural levels during the heat denaturation of β-lactoglobulin. Eur. J. Biochem. 1996, 237, 106–112. [Google Scholar] [CrossRef]
- Slattery, H.; Fitzgerald, R.J. Functional Properties and Bitterness of Sodium Caseinate Hydrolysates Prepared with a Bacillus Proteinase. J. Food Sci. 1998, 63, 418–422. [Google Scholar] [CrossRef]
- Nam, S.-H.; Wagh, A.; Martini, S.; Walsh, M.K. Sensory characterisation of a high-protein beverage. Int. J. Dairy Technol. 2017, 70, 432–438. [Google Scholar] [CrossRef]
- Lesme, H.; Rannou, C.; Loisel, C.; Famelart, M.H.; Bouhallab, S.; Prost, C. Controlled whey protein aggregates to modulate the texture of fat-free set-type yoghurts. Int. Dairy J. 2019, 92, 28–36. [Google Scholar] [CrossRef]
- Gélébart, P.; Riaublanc, A.; Famelart, M.H.; Jonchère, C.; Beaumal, V.; Anton, M.; Garnier, C. Protein aggregates modulate the texture of emulsified and acidified acid milk gels. Food Hydrocoll. 2019, 93, 176–188. [Google Scholar] [CrossRef]
- Jung, S.; Murphy, P.A.; Johnson, L.A. Physicochemical and Functional Properties of Soy Protein Substrates Modified by Low Levels of Protease Hydrolysis. J. Food Sci. 2006, 70, C180–C185. [Google Scholar] [CrossRef]
- Diniz, F.M.; Martin, A.M. Effects of the Extent of Enzymatic Hydrolysis on Functional Properties of Shark Protein Hydrolysate. LWT-Food Sci. Technol. 1997, 30, 266–272. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Otte, J.; Madsen, J.S.; Qvist, K.B. Effects of limited proteolysis on gelation and gel properties of whey protein isolate. J. Dairy Sci. 1995, 78, 2119–2128. [Google Scholar] [CrossRef]
- Otte, J.; Ju, Z.Y.; Skriver, A.; Qvist, K.B. Effects of limited proteolysis on the microstructure of heat-induced whey protein gels at varying pH. J. Dairy Sci. 1996, 79, 782–790. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Enzyme | DH (%) at Different Incubation Times | |
|---|---|---|---|
| 1 h | 4 h | ||
| MPC | Control | 0.92 ± 0.21 e | 1.04 ± 0.37 e |
| Pro | 7.05 ± 0.40 c | 11.65 ± 0.98 b | |
| Deb | 8.17 ± 0.58 c | 15.74 ± 1.36 a | |
| WPC | Control | 2.11 ± 0.15 d | 2.26 ± 0.07 d |
| Pro | 7.12 ± 0.50 c | 9.54 ± 1.09 b | |
| Deb | 6.56 ± 0.38 c | 12.04 ± 0.73 a | |
| NaCN | Control | 0.30 ± 0.26 e | 0.66 ± 0.26 e |
| Pro | 7.18 ± 0.67 d | 11.67 ± 0.67 b | |
| deb | 9.48 ± 0.88 c | 17.78 ± 1.00 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruppi, A.; Dermiki, M.; Spigno, G.; FitzGerald, R.J. Impact of Enzymatic Hydrolysis and Heat Inactivation on the Physicochemical Properties of Milk Protein Hydrolysates. Foods 2022, 11, 516. https://doi.org/10.3390/foods11040516
Gruppi A, Dermiki M, Spigno G, FitzGerald RJ. Impact of Enzymatic Hydrolysis and Heat Inactivation on the Physicochemical Properties of Milk Protein Hydrolysates. Foods. 2022; 11(4):516. https://doi.org/10.3390/foods11040516
Chicago/Turabian StyleGruppi, Alice, Maria Dermiki, Giorgia Spigno, and Richard J. FitzGerald. 2022. "Impact of Enzymatic Hydrolysis and Heat Inactivation on the Physicochemical Properties of Milk Protein Hydrolysates" Foods 11, no. 4: 516. https://doi.org/10.3390/foods11040516