
Characterization of the Reduced IgE Binding Capacity in Boiled and Autoclaved Soybeans through Proteomic Approaches

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Pre-Treatment of Boiled and Autoclaved Soybeans
2.3. Preparation of Soybean Proteins
2.4. Determination of IgE Binding Capacity by Indirect ELISA
2.5. SDS-PAGE
2.6. LC/MS-MS
2.7. Statistical Analysis
3. Results and Discussion
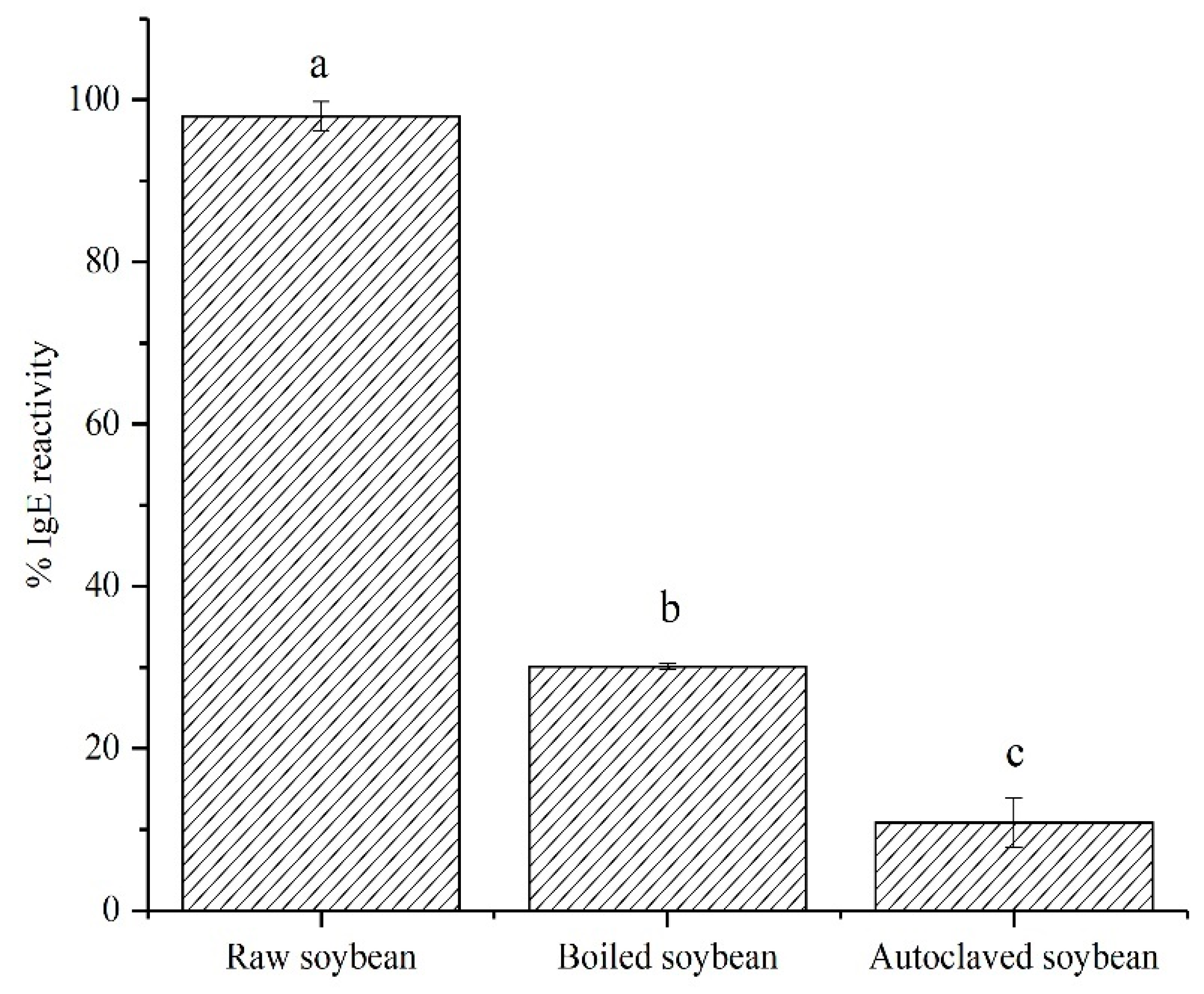
3.1. The IgE Binding Capacity of Boiled and Autoclaved Soybeans
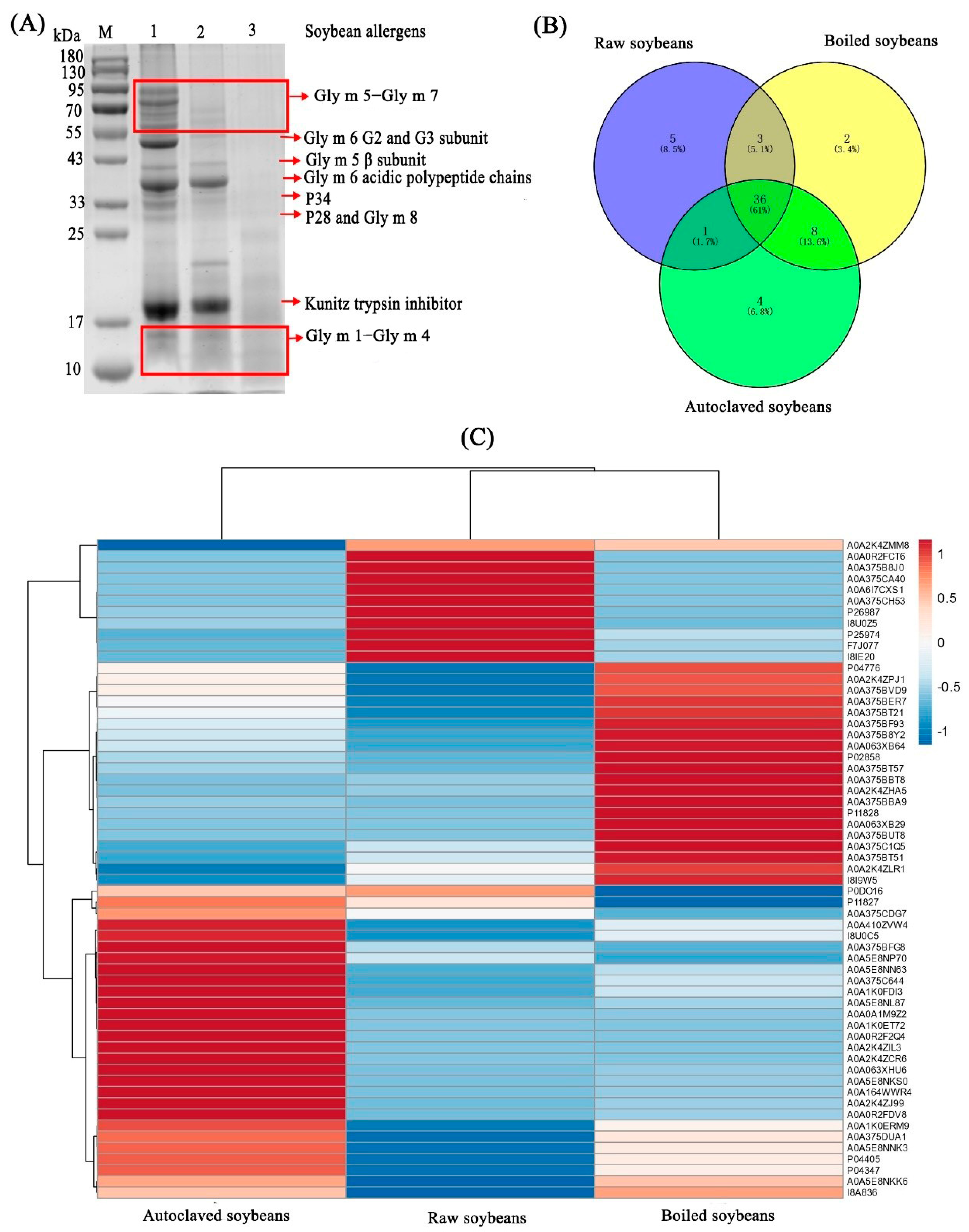
3.2. Changes in Protein Profiles of Extracted Protein
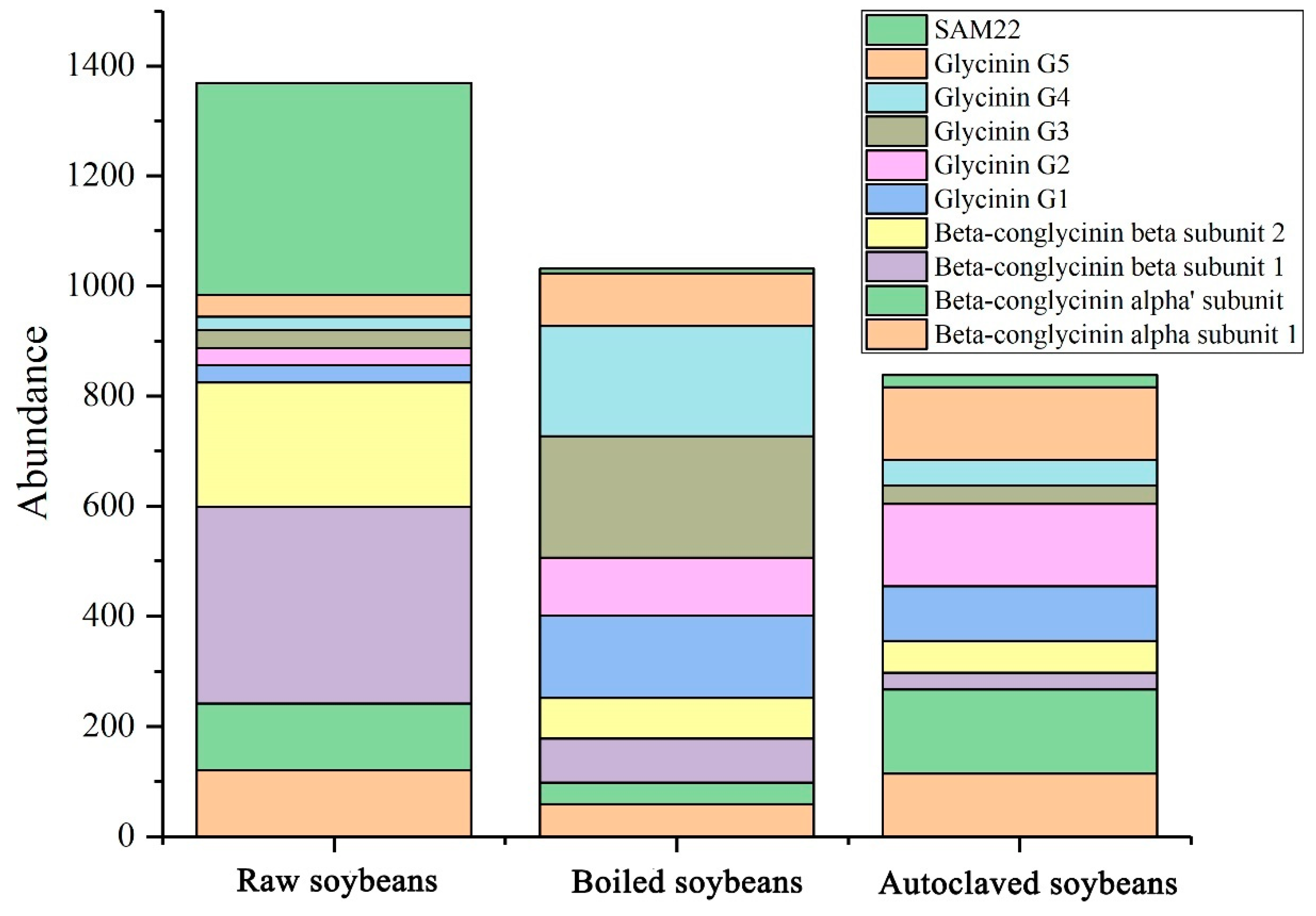
3.3. Changes in Identified Soybean Allergens
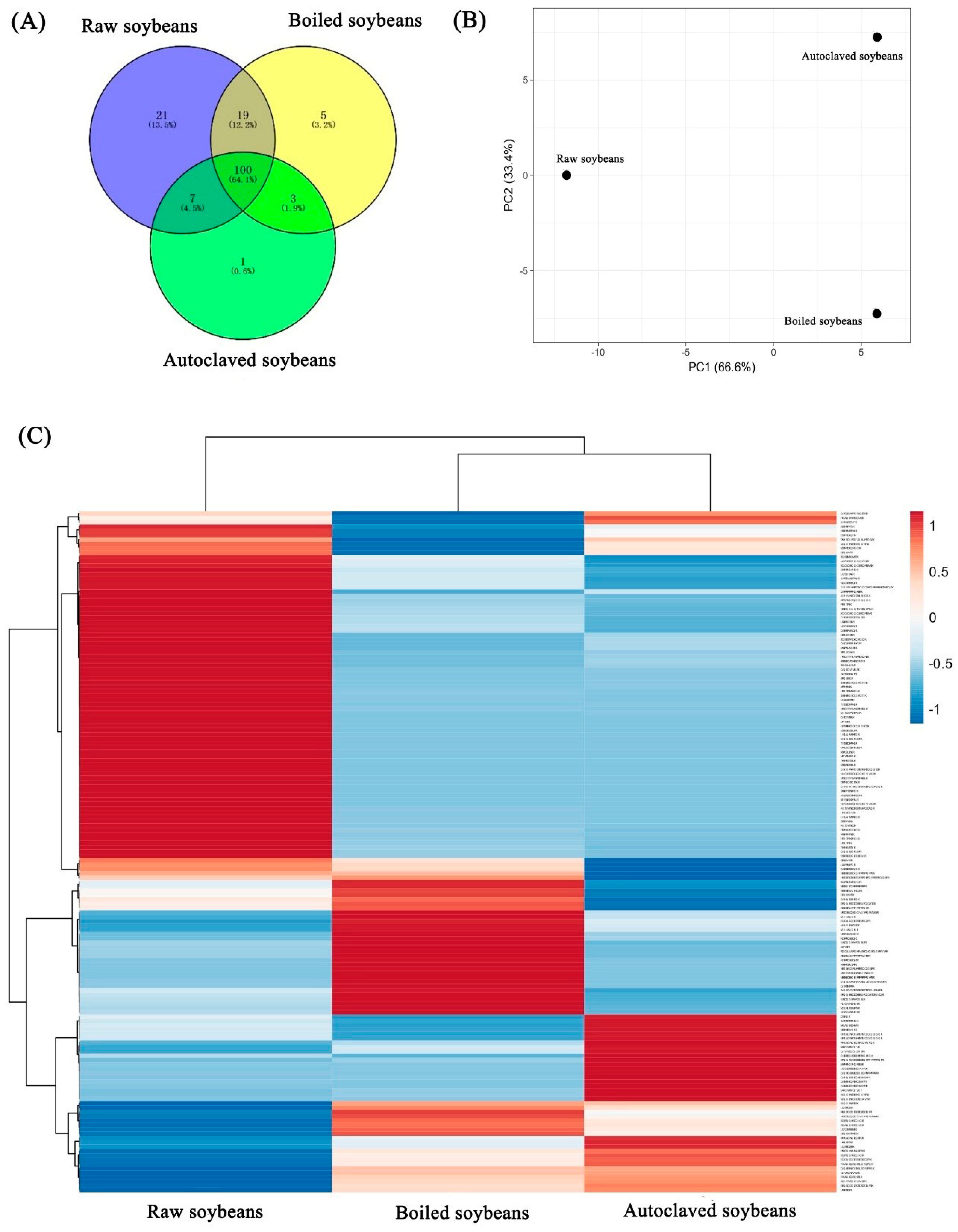
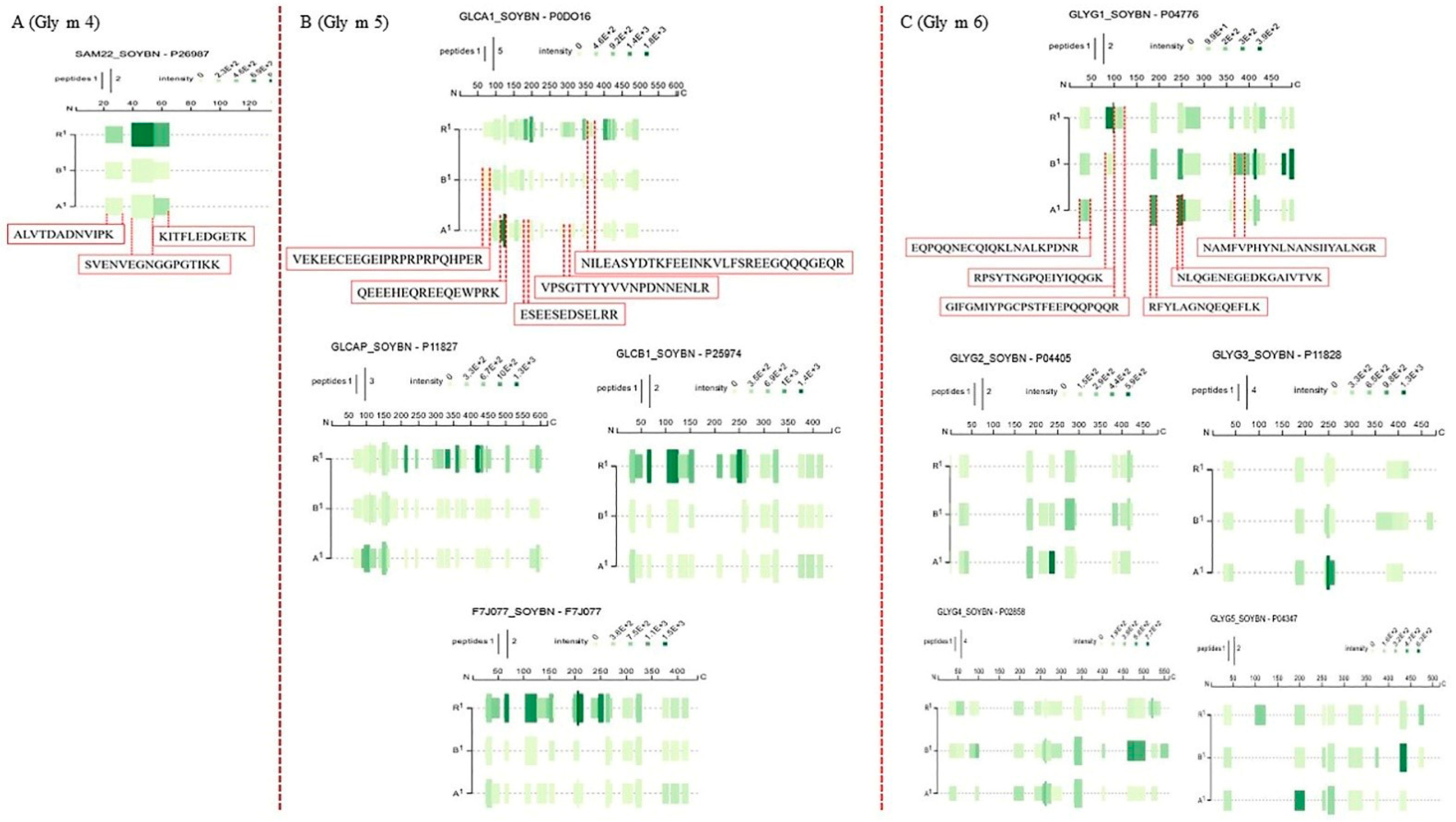
3.4. Changes in Peptide Profiles in Identified Soybean Allergens
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geng, T.; Liu, K.; Frazier, R.; Shi, L.; Bell, E.; Glenn, K.; Ward, J.M. Development of a Sandwich ELISA for Quantification of Gly m 4, a Soybean Allergen. J. Agric. Food Chem. 2015, 63, 4947–4953. [Google Scholar] [CrossRef] [PubMed]
- Pi, X.; Sun, Y.; Fu, G.; Wu, Z.; Cheng, J. Effect of processing on soybean allergens and their allergenicity. Trends Food Sci. Technol. 2021, 118, 316–327. [Google Scholar] [CrossRef]
- Burney, P.; Summers, C.; Chinn, S.; Hooper, R.; van Ree, R.; Lidholm, J. Prevalence and distribution of sensitization to foods in the European Community Respiratory Health Survey: A EuroPrevall analysis. Allergy 2010, 65, 1182–1188. [Google Scholar] [CrossRef]
- Ballmer-Weber, B.K.; Holzhauser, T.; Scibilia, J.; Mittag, D.; Zisa, G.; Ortolani, C.; Oesterballe, M.; Poulsen, L.K.; Vieths, S.; Bindslev-Jensen, C. Clinical characteristics of soybean allergy in Europe: A double-blind, placebo-controlled food challenge study. J. Allergy Clin. Immunol. 2007, 119, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- The World Health Organization and International Union of Immunological Societies (WHO/IUIS) Allergen Nomenclature Subcommittee. Available online: http://www.allergen.org/index.php (accessed on 1 September 2021).
- Amnuaycheewa, P.; de Mejia, E.G. Purification, characterisation, and quantification of the soy allergen profilin (Gly m 3) in soy products. Food Chem. 2010, 119, 1671–1680. [Google Scholar] [CrossRef]
- Chi, C.-H.; Cho, S.-J. Improvement of bioactivity of soybean meal by solid-state fermentation with Bacillus amyloliquefaciens versus Lactobacillus spp. and Saccharomyces cerevisiae. LWT-Food Sci. Technol. 2016, 68, 619–625. [Google Scholar] [CrossRef]
- Jimenez-Saiz, R.; Benede, S.; Molina, E.; Lopez-Exposito, I. Effect of processing technologies on the allergenicity of food products. Crit. Rev. Food Sci. Nutr. 2015, 55, 1902–1917. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.W.; Hsu, C.K.; Yang, Y.F. Effect of thermal treatments on anti-nutritional factors and antioxidant capabilities in yellow soybeans and green-cotyledon small black soybeans. J. Sci. Food Agric. 2014, 94, 1794–1801. [Google Scholar] [CrossRef]
- Pi, X.; Sun, Y.; Guo, X.; Chen, Q.; Cheng, J.; Guo, M. Effects of thermal sterilization on the allergenicity of soybeans. LWT 2022, 154, 112678. [Google Scholar] [CrossRef]
- Xia, J.; Zu, Q.; Yang, A.; Wu, Z.; Li, X.; Tong, P.; Yuan, J.; Wu, Y.; Fan, Q.; Chen, H. Allergenicity reduction and rheology property of Lactobacillus-fermented soymilk. J. Sci. Food Agric. 2019, 99, 6841–6849. [Google Scholar] [CrossRef]
- Wilson, S.; Martinez-Villaluenga, C.; De Mejia, E.G. Purification, thermal stability, and antigenicity of the immunodominant soybean allergen P34 in soy cultivars, ingredients, and products. J. Food Sci. 2008, 73, T106–T114. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B.; Maleki, S.J.; Rodriguez, J.; Burbano, C.; Muzquiz, M.; Jimenez, M.A.; Pedrosa, M.M.; Cuadrado, C.; Crespo, J.F. Heat and pressure treatments effects on peanut allergenicity. Food Chem. 2012, 132, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B.; Cuadrado, C.; Rodriguez, J.; Hart, J.; Burbano, C.; Crespo, J.F.; Novak, N. Potential changes in the allergenicity of three forms of peanut after thermal processing. Food Chem. 2015, 183, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Li, S.; Xu, H.; Zhang, T.; Lin, X.; Wu, X. Effect of Covalent Interaction with Chlorogenic Acid on the Allergenic Capacity of Ovalbumin. J. Agric. Food Chem. 2018, 66, 9794–9800. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, G.; Tu, Z.; Wang, H.; Hu, Y.; Mao, J.; Zhang, J. Insight into the Mechanism of Reduced IgG/IgE Binding Capacity in Ovalbumin as Induced by Glycation with Monose Epimers through Liquid Chromatography and High-Resolution Mass Spectrometry. J. Agric. Food Chem. 2020, 68, 6065–6075. [Google Scholar] [CrossRef]
- Liu, G.-X.; Tu, Z.-C.; Yang, W.; Wang, H.; Zhang, L.; Ma, D.; Huang, T.; Liu, J.; Li, X. Investigation into allergenicity reduction and glycation sites of glycated beta-lactoglobulin with ultrasound pretreatment by high-resolution mass spectrometry. Food Chem. 2018, 252, 99–107. [Google Scholar] [CrossRef]
- Yang, W.; Tu, Z.; Wang, H.; Zhang, L.; Xu, S.; Niu, C.; Yao, H.; Kaltashov, I.A. Mechanism of Reduction in IgG and IgE Binding of beta-Lactoglobulin Induced by Ultrasound Pretreatment Combined with Dry-State Glycation: A Study Using Conventional Spectrometry and High-Resolution Mass Spectrometry. J. Agric. Food Chem. 2017, 65, 8018–8027. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.; Sun, X.; Jiang, S.; Guo, M. Characterization of the milk fat globule membrane proteome in colostrum and mature milk of Xinong Saanen goats. J. Dairy Sci. 2020, 103, 3017–3024. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, N.; Wang, C.; Cheng, J.; Guo, M. Proteomic analysis of differentially expressed whey proteins in Saanen goat milk from different provinces in China using a data-independent acquisition technique. J. Dairy Sci 2021, 104, 10513–10527. [Google Scholar] [CrossRef]
- Huang, X.; Zhu, Z.; Feng, H.; Zhang, Q.; Zhang, H. Simultaneous determination of multi-allergens in surimi products by LC-MS/MS with a stable isotope-labeled peptide. Food Chem. 2020, 320, 126580. [Google Scholar] [CrossRef]
- Yoshimitsu, M.; Kiyota, K.; Kajimura, K.; Yamano, T. Development of an LC-MS/MS-based analytical method for quantification of soybean allergen Gly m 4 in soybean grains and processed foods. Food Agric. Immunol. 2018, 30, 25–33. [Google Scholar] [CrossRef]
- Rui, X.; Huang, J.; Xing, G.; Zhang, Q.; Li, W.; Dong, M. Changes in soy protein immunoglobulin E reactivity, protein degradation, and conformation through fermentation with Lactobacillus plantarum strains. LWT-Food Sci. Technol. 2019, 99, 156–165. [Google Scholar] [CrossRef]
- He, W.; He, K.; Sun, F.; Mu, L.; Liao, S.; Li, Q.; Yi, J.; Liu, Z.; Wu, X. Effect of heat, enzymatic hydrolysis and acid-alkali treatment on the allergenicity of silkworm pupa protein extract. Food Chem. 2021, 343, 128461. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, X.; Song, X.; Guo, M. Effects of kefir grains from different origins on proteolysis and volatile profile of goat milk kefir. Food Chem. 2021, 339, 128099. [Google Scholar] [CrossRef] [PubMed]
- Bloom, K.A.; Huang, F.R.; Bencharitiwong, R.; Bardina, L.; Ross, A.; Sampson, H.A.; Nowak-Węgrzyn, A. Effect of Heat Treatment on Milk and Egg Proteins Allergenicity. Pediatr. Allergy Immunol. 2014, 25, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Blanc, F.; Vissers, Y.M.; Adel-Patient, K.; Rigby, N.M.; Mackie, A.R.; Gunning, A.P.; Wellner, N.K.; Skov, P.S.; Przybylski-Nicaise, L.; Ballmer-Weber, B.; et al. Boiling peanut Ara h 1 results in the formation of aggregates with reduced allergenicity. Mol. Nutr. Food Res. 2011, 55, 1887–1894. [Google Scholar] [CrossRef]
- Pi, X.; Fu, G.; Dong, B.; Yang, Y.; Wan, Y.; Xie, M. Effects of fermentation with Bacillus natto on the allergenicity of peanut. LWT 2021, 141, 110862. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, Q.; Zhang, T.; Cai, Q.; Chen, Q. Thermal processing effects on peanut allergen Ara h 2 allergenicity in mice and its antigenic epitope structure. Food Chem. 2016, 212, 657–662. [Google Scholar] [CrossRef]
- Kim, K.-B.-W.-R.; Lee, S.-Y.; Song, E.-J.; Kim, K.-E.; Ahn, D.-H. Effect of Heat and Autoclave on Allergenicity of Porcine Serum Albumin. Food Sci. Biotechnol. 2011, 20, 455–459. [Google Scholar] [CrossRef]
- Tong, P.; Gao, J.; Chen, H.; Li, X.; Zhang, Y.; Jian, S.; Wichers, H.; Wu, Z.; Yang, A.; Liu, F. Effect of heat treatment on the potential allergenicity and conformational structure of egg allergen ovotransferrin. Food Chem. 2012, 131, 603–610. [Google Scholar] [CrossRef]
- Costa, J.; Bavaro, S.L.; Benede, S.; Diaz-Perales, A.; Bueno-Diaz, C.; Gelencser, E.; Klueber, J.; Larre, C.; Lozano-Ojalvo, D.; Lupi, R.; et al. Are Physicochemical Properties Shaping the Allergenic Potency of Plant Allergens? Clin. Rev. Allergy Immunol. 2020, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Kasera, R.; Singh, A.B.; Kumar, R.; Lavasa, S.; Prasad, K.N.; Arora, N. Effect of thermal processing and gamma-irradiation on allergenicity of legume proteins. Food Chem. Toxicol. 2012, 50, 3456–3461. [Google Scholar] [CrossRef] [PubMed]
- Geng, T.; Stojšin, D.K.; Liu, K.; Schaalje, B.; Postin, C.; Ward, J.; Wang, Y.; Liu, Z.L.; Li, B.; Glenn, K. Natural Variability of Allergen Levels in Conventional Soybeans: Assessing Variation across North and South America from Five Production Years. J. Agric. Food Chem. 2017, 65, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Berneder, M.; Bublin, M.; Hoffmann-Sommergruber, K.; Hawranek, T.; Lang, R. Allergen chip diagnosis for soy-allergic patients: Gly m 4 as a marker for severe food-allergic reactions to soy. Int. Arch. Allergy Immunol. 2013, 161, 229–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husslik, F.; Mittag, D.; von Loetzen, C.S.; Hartl, M.; Vogel, L.; Ballmer-Weber, B.; Kleine-Tebbe, J.; Treudler, R.; Simon, J.-C.; Hanschmann, K.-M.; et al. Analysis of the IgE epitope profile of soybean allergen Gly m 4. Clin. Transl. Allergy 2014, 4, 42. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Teng, D.; Yang, Y.; Wang, X.; Wang, J. Development of a competitive ELISA for the detection of soybean α subunit of β-conglycinin. Process Biochem. 2012, 47, 280–287. [Google Scholar] [CrossRef]
- Sun, X.; Shan, X.; Yan, Z.; Zhang, Y.; Guan, L. Prediction and characterization of the linear IgE epitopes for the major soybean allergen beta-conglycinin using immunoinformatics tools. Food Chem. Toxicol. 2013, 56, 254–260. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pi, X.; Sun, Y.; Deng, X.; Xin, D.; Cheng, J.; Guo, M. Characterization of the Reduced IgE Binding Capacity in Boiled and Autoclaved Soybeans through Proteomic Approaches. Foods 2022, 11, 479. https://doi.org/10.3390/foods11030479
Pi X, Sun Y, Deng X, Xin D, Cheng J, Guo M. Characterization of the Reduced IgE Binding Capacity in Boiled and Autoclaved Soybeans through Proteomic Approaches. Foods. 2022; 11(3):479. https://doi.org/10.3390/foods11030479
Chicago/Turabian StylePi, Xiaowen, Yuxue Sun, Xiaomin Deng, Dawei Xin, Jianjun Cheng, and Mingruo Guo. 2022. "Characterization of the Reduced IgE Binding Capacity in Boiled and Autoclaved Soybeans through Proteomic Approaches" Foods 11, no. 3: 479. https://doi.org/10.3390/foods11030479