
The Correlation Mechanism between Dominant Bacteria and Primary Metabolites during Fermentation of Red Sour Soup

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Sour Soup
2.2. Inoculant Fermentation
2.3. DNA Extraction, PCR Amplification and Metagenomic Sequencing
2.4. UPLC–MS/MS Conditions for Metabolome Analysis
2.4.1. Liquid Phase Conditions
2.4.2. Conditions for Mass Spectrometry
2.5. Statistical Analysis
3. Results
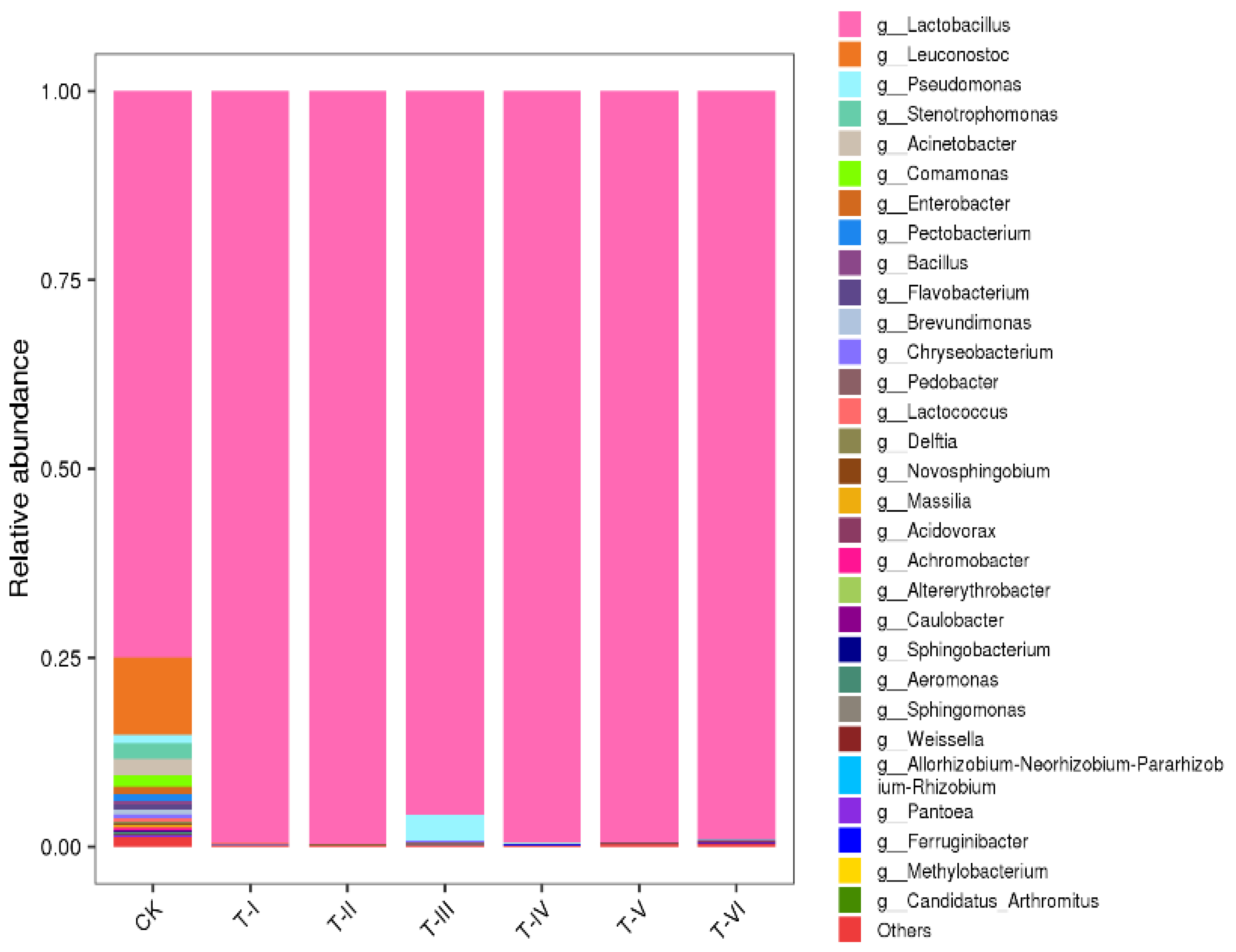
3.1. Metagenomic Sequencing and Relative Abundance of Different Microbial Communities
3.2. Alpha Diversity Indices of Bacterial Populations
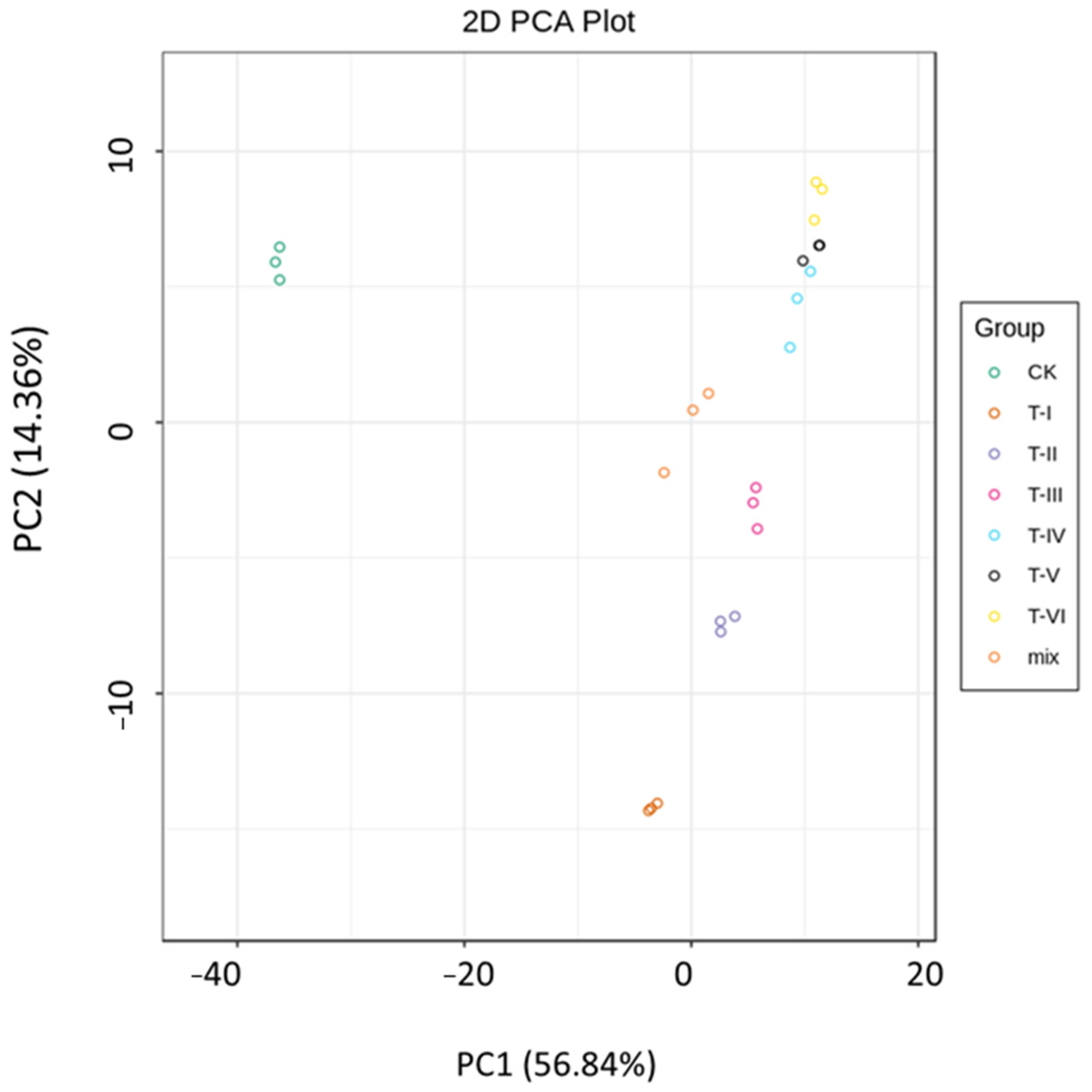
3.3. Beta Diversity Analyses of Bacterial Composition (through NMDS, PCA and PCOA)
3.4. Network Map of Dominant Species and Venn Diagram Analysis
3.5. LDA Effect Size (LEfSe) Analysis
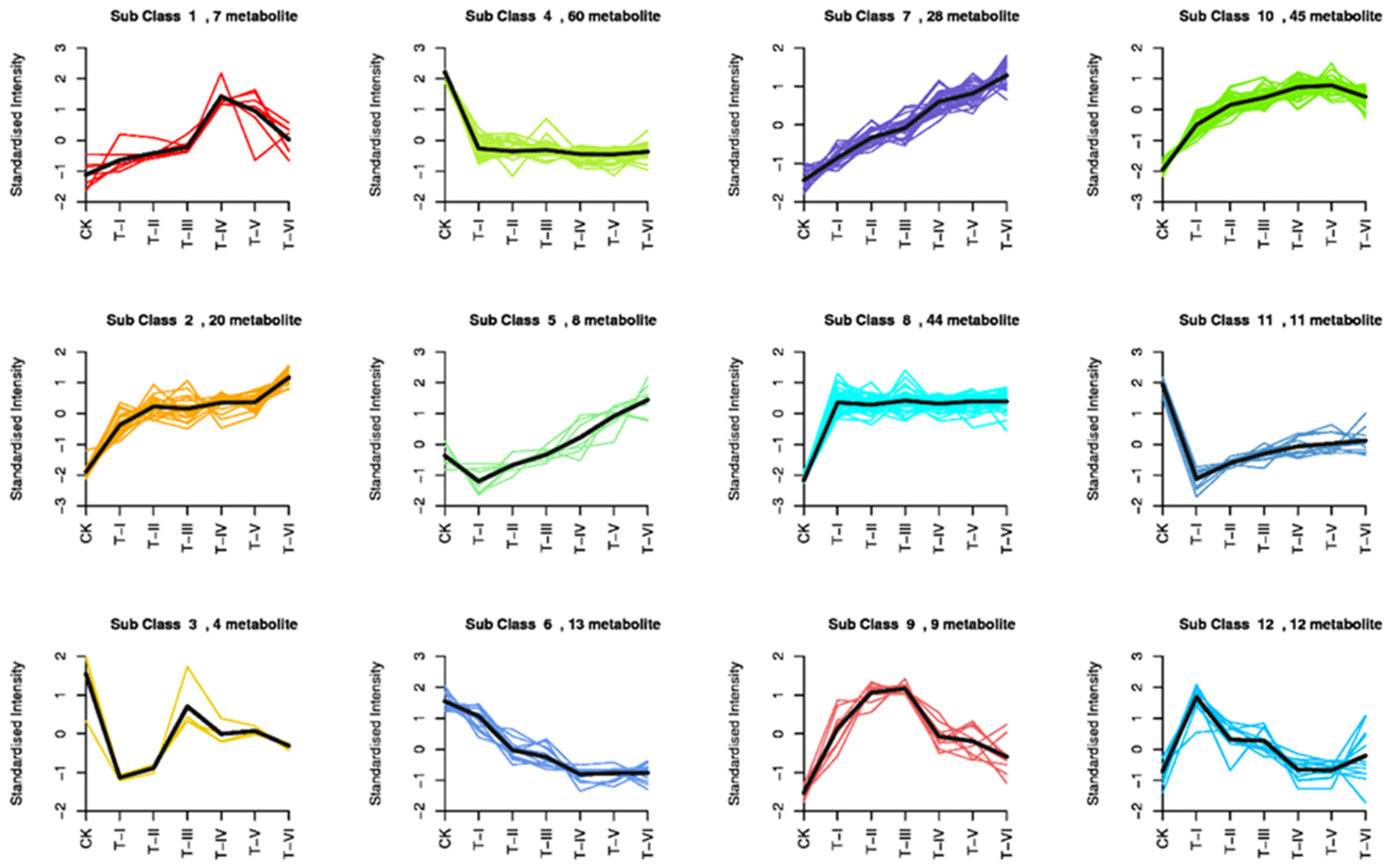
3.6. Analysis of the Major Metabolic Products
3.7. Orthogonal Least Multiplier-Differential Analysis (OPLS-DA)
3.8. Screening of Differential Metabolites
3.9. KEGG Classification and Differential Metabolite Enrichment Analysis
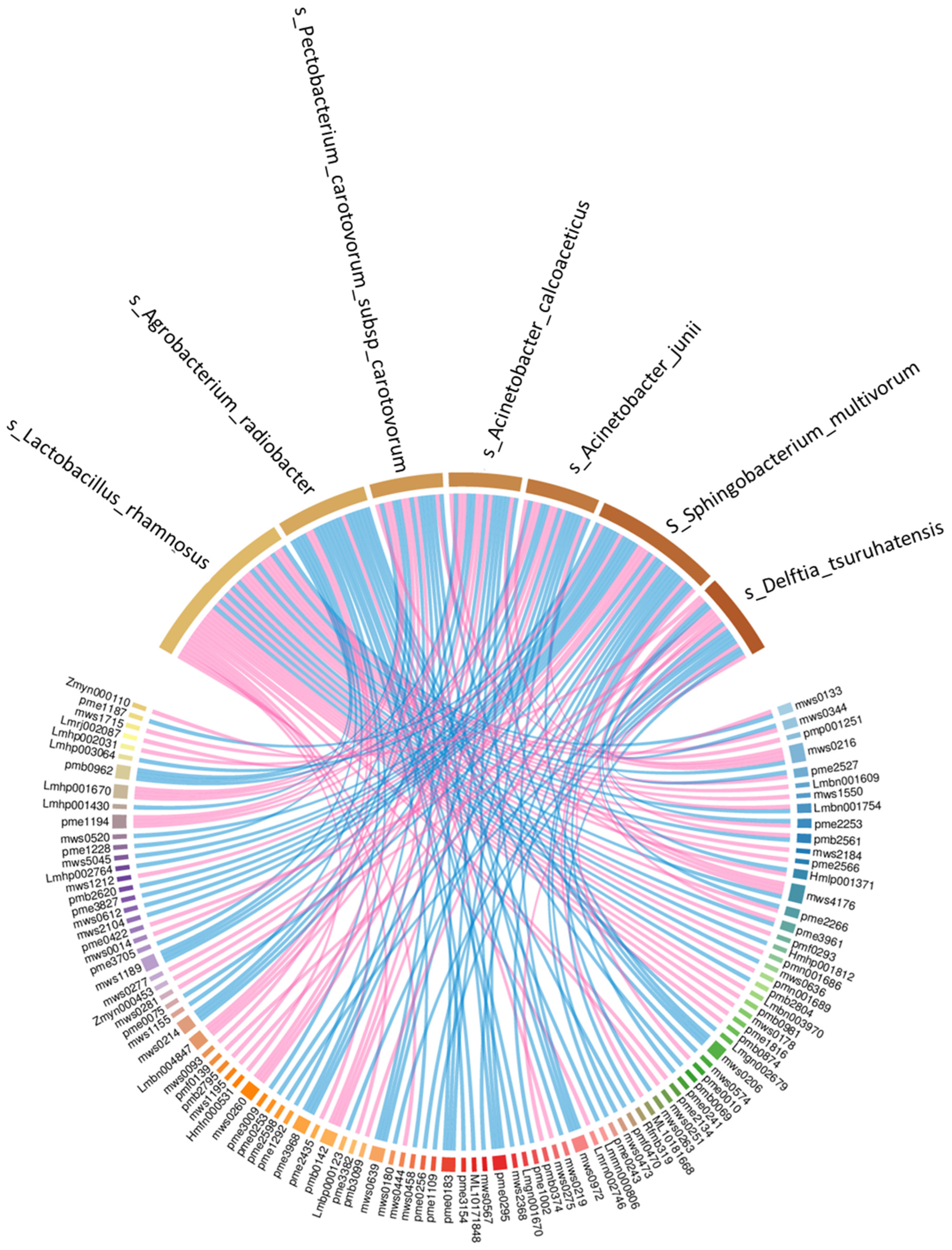
3.10. Correlation between Dominant Bacteria and Primary Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yangbo, H.; Yongfu, L.; Xingbang, L.; Guolin, L.; Zhaoyan, D.; Chaojun, C. Effects of thermal and nonthermal processing technology on the quality of red sour soup after storage. Food Sci. Nutr. 2021, 9, 3863–3872. [Google Scholar] [CrossRef]
- Fang, G.-Y.; Chai, L.-J.; Zhong, X.-Z.; Jiang, Y.-J. Deciphering the succession patterns of bacterial community and their correlations with environmental factors and flavor compounds during the fermentation of Zhejiang rosy vinegar. Int. J. Food Microbiol. 2021, 341, 109070. [Google Scholar] [CrossRef]
- Xun, W.; Liu, Y.; Li, W.; Ren, Y.; Xiong, W.; Xu, Z.; Zhang, N.; Miao, Y.; Shen, Q.; Zhang, R. Specialized metabolic functions of keystone taxa sustain soil microbiome stability. Microbiome 2021, 9, 35. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, Q.; He, L.; Li, C. Determination of the microbial communities of Guizhou Suantang, a traditional Chinese fermented sour soup, and correlation between the identified microorganisms and volatile compounds. Food Res. Int. 2020, 138, 109820. [Google Scholar] [CrossRef]
- Xiong, K.; Han, F.; Wang, Z.; Du, M.; Chen, Y.; Tang, Y.; Wang, Z. Screening of dominant strains in red sour soup from Miao nationality and the optimization of inoculating fermentation conditions. Food Sci. Nutr. 2021, 9, 261–271. [Google Scholar] [CrossRef]
- Rakhmanova, A.; Khan, Z.; Shah, K. A mini review fermentation and preservation: Role of lactic acid bacteria. MOJ Food Process. Technol. 2018, 6, 414–417. [Google Scholar] [CrossRef]
- Juodeikiene, G.; Bartkiene, E.; Viskelis, P.; Urbonaviciene, D.; Eidukonyte, D.; Bobinas, C. Fermentation processes using lactic acid bacteria producing bacteriocins for preservation and improving functional properties of food products. Adv. Appl. Biotechnol. 2012, 63–100. [Google Scholar]
- Lin, L.; Wu, J.; Chen, X.; Huang, L.; Zhang, X.; Gao, X. The Role of the Bacterial Community in Producing a Peculiar Smell in Chinese Fermented Sour Soup. Microorganisms 2020, 8, 1270. [Google Scholar] [CrossRef]
- Yang-bo, H.; Yong-fu, L.; Ding-jiang, Z.; Zhao-yan, D.; Qian, L.; Bin, S.; Zhu-xi, T. Analysis on the Flavor Difference of Red Sour Soup Including Urn Odor and Normal Flavor Samples with Electronic Nose and Gas Chromatography-Ion Mobility Spectroscopy. Food Indust. Tech. 2020, 41, 216–221, 227. [Google Scholar] [CrossRef]
- Liu, Z.; Li, J.; Wei, B.; Huang, T.; Xiao, Y.; Peng, Z.; Xie, M.; Xiong, T. Bacterial community and composition in Jiang-shui and Suan-cai revealed by high-throughput sequencing of 16S rRNA. Int. J. Food Microbiol. 2019, 306, 108271. [Google Scholar] [CrossRef]
- Papagianni, M. Recent advances in solid-state fermentation applications for the food industry. Curr. Biochem. Eng. 2014, 1, 2–8. [Google Scholar] [CrossRef]
- Wang, S.; Chen, H.; Sun, B. Recent progress in food flavor analysis using gas chromatography–ion mobility spectrometry (GC–IMS). Food Chem. 2020, 315, 126158. [Google Scholar] [CrossRef]
- Diez-Simon, C.; Mumm, R.; Hall, R.D. Mass spectrometry-based metabolomics of volatiles as a new tool for understanding aroma and flavour chemistry in processed food products. Metabolomics 2019, 15, 41. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.F.; Zhou, B.; Ressom, H.W. Metabolite identification and quantitation in LC-MS/MS-based metabolomics. Trends Analyt. Chem. 2012, 32, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Wang, D.; Li, X.; Li, Z.; Chao, J.; Qin, X. Metabolomic study of the fever model induced by baker’s yeast and the antipyretic effects of aspirin in rats using nuclear magnetic resonance and gas chromatography–mass spectrometry. J. Pharm. Biomed. Anal. 2013, 81, 168–177. [Google Scholar] [CrossRef]
- Wouters, D.; Grosu-Tudor, S.; Zamfir, M.; De Vuyst, L. Applicability of Lactobacillus plantarum IMDO 788 as a starter culture to control vegetable fermentations. J. Sci. Food Agric. 2013, 93, 3352–3361. [Google Scholar] [CrossRef]
- Silva, C.C.; Silva, S.P.; Ribeiro, S.C. Application of bacteriocins and protective cultures in dairy food preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Raj, T.; Chandrasekhar, K.; Kumar, A.N.; Kim, S.-H. Recent biotechnological trends in lactic acid bacterial fermentation for food processing industries. Syst. Microbiol. Biomanuf. 2021, 2, 14–40. [Google Scholar] [CrossRef]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [Green Version]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665. [Google Scholar] [CrossRef]
- Sharma, R.; Garg, P.; Kumar, P.; Bhatia, S.K.; Kulshrestha, S. Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods. Fermentation 2020, 6, 106. [Google Scholar] [CrossRef]
- Abbasiliasi, S.; Tan, J.S.; Ibrahim, T.A.T.; Bashokouh, F.; Ramakrishnan, N.R.; Mustafa, S.; Ariff, A.B. Fermentation factors influencing the production of bacteriocins by lactic acid bacteria: A review. Rsc Adv. 2017, 7, 29395–29420. [Google Scholar] [CrossRef]
- Kuley, E.; Ozogul, F.; Ozogul, Y.; Akyol, I. The function of lactic acid bacteria and brine solutions on biogenic amine formation by foodborne pathogens in trout fillets. Food Chem. 2011, 129, 1211–1216. [Google Scholar] [CrossRef]
- Bartkiene, E.; Bartkevics, V.; Mozuriene, E.; Krungleviciute, V.; Novoslavskij, A.; Santini, A.; Rozentale, I.; Juodeikiene, G.; Cizeikiene, D. The impact of lactic acid bacteria with antimicrobial properties on biodegradation of polycyclic aromatic hydrocarbons and biogenic amines in cold smoked pork sausages. Food Control 2017, 71, 285–292. [Google Scholar] [CrossRef]
- Wedajo, B. Lactic acid bacteria: Benefits, selection criteria and probiotic potential in fermented food. J. Prob. Health 2015, 3, 100129. [Google Scholar] [CrossRef]
- Sindhu, S.C.; Khetarpaul, N. Probiotic fermentation of indigenous food mixture: Effect on antinutrients and digestibility of starch and protein. J. Food Compos. Anal. 2001, 14, 601–609. [Google Scholar] [CrossRef]
- Jiao, A.; Xu, X.; Jin, Z. Research progress on the brewing techniques of new-type rice wine. Food Chem. 2017, 215, 508–515. [Google Scholar] [CrossRef]
- Liu, N.; Miao, S.; Qin, L. Regulatory Mechanisms of L-lactic acid and Taste Substances in Chinese Acid Rice Soup (Rice-acid) fermented with a Lacticaseibacillus paracasei and Kluyveromyces marxianus. Front. Microbiol. 2021, 12, 1078. [Google Scholar] [CrossRef]
- Xu, X.; Wu, B.; Zhao, W.; Pang, X.; Lao, F.; Liao, X.; Wu, J. Correlation between autochthonous microbial communities and key odorants during the fermentation of red pepper (Capsicum annuum L.). Food Microbiol. 2020, 91, 103510. [Google Scholar] [CrossRef]
- Miyamae, Y.; Kurisu, M.; Han, J.; Isoda, H.; Shigemori, H. Structure-activity relationship of caffeoylquinic acids on the accelerating activity on ATP production. Chem. Pharm. Bull. 2011, 59, 502–507. [Google Scholar] [CrossRef] [Green Version]
- Gaby, S.K. Vitamin Intake and Health: A scientific Review; CRC Press: Boca Raton, FL, USA, 1990; pp. 71–103. [Google Scholar]
- Schectman, G.; Byrd, J.C.; Hoffmann, R. Ascorbic acid requirements for smokers: Analysis of a population survey. Am. J. Clin. Nutr. 1991, 53, 1466–1470. [Google Scholar] [CrossRef] [PubMed]
- Cioroi, M. Study on L-ascorbic acid contents from exotic fruits. Cercet. Agron. Mold. 2007, 1, 23–27. [Google Scholar]
- Miyake, M.; Kirisako, T.; Kokubo, T.; Miura, Y.; Morishita, K.; Okamura, H.; Tsuda, A. Randomised controlled trial of the effects of L-ornithine on stress markers and sleep quality in healthy workers. Nutr. J. 2014, 13, 53. [Google Scholar] [CrossRef] [Green Version]
- Pangallo, D.; Kraková, L.; Puškárová, A.; Šoltys, K.; Bučková, M.; Koreňová, J.; Budiš, J.; Kuchta, T. Transcription activity of lactic acid bacterial proteolysis-related genes during cheese maturation. Food Microbiol. 2019, 82, 416–425. [Google Scholar] [CrossRef]
- Wang, C.; Sun, J.; Lassabliere, B.; Yu, B.; Liu, S.Q. Coffee flavour modification through controlled fermentation of green coffee beans by Lactococcus lactis subsp. cremoris. LWT 2020, 120, 108930. [Google Scholar] [CrossRef]
- Shimizu, Y.; Sakuraba, H.; Doi, K.; Ohshima, T. Molecular and functional characterization of D-3-phosphoglycerate dehydrogenase in the serine biosynthetic pathway of the hyperthermophilic archaeon Sulfolobus tokodaii. Arch. Biochem. Biophys. 2008, 470, 120–128. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Code | Fermentation Period |
|---|---|
| CK | Samples fermented for 0 Days |
| T-I | Samples fermented for 5 Days |
| T-II | Samples fermented for 10 Days |
| T-III | Samples fermented for 15 Days |
| T-IV | Samples fermented for 20 Days |
| T-V | Samples fermented for 25 Days |
| T-VI | Samples fermented for 30 Days |
| Parameter | CK | T-I | T-II | T-III | T-IV | T-V | T-VI | SEM | p Value |
|---|---|---|---|---|---|---|---|---|---|
| Chao1 | 80.37 | 49.14 | 80.71 | 46.72 | 63.22 | 74.50 | 58.93 | 15.10 | 0.53 |
| ACE | 82.21 | 50.14 | 77.40 | 47.84 | 66.43 | 75.07 | 68.39 | 11.81 | 0.33 |
| Observed Species | 49.00 a | 24.33 b | 31.33 ab | 26.33 b | 34.33 ab | 43.33 ab | 47.00 a | 4.21 | 0.001 |
| Shannon | 1.79 a | 0.70 c | 0.10 c | 0.29 bc | 0.10 c | 0.11 c | 0.15 c | 0.10 c | 0.001 |
| Simpson | 0.43 a | 0.24 b | 0.02 c | 0.08 c | 0.02 c | 0.02 c | 0.02 c | 0.03 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Liu, Z.; Xie, L.; Li, L.; Zhou, W.; Zhao, L. The Correlation Mechanism between Dominant Bacteria and Primary Metabolites during Fermentation of Red Sour Soup. Foods 2022, 11, 341. https://doi.org/10.3390/foods11030341
Zhou X, Liu Z, Xie L, Li L, Zhou W, Zhao L. The Correlation Mechanism between Dominant Bacteria and Primary Metabolites during Fermentation of Red Sour Soup. Foods. 2022; 11(3):341. https://doi.org/10.3390/foods11030341
Chicago/Turabian StyleZhou, Xiaojie, Zhiqi Liu, Le Xie, Liangyi Li, Wenhua Zhou, and Liangzhong Zhao. 2022. "The Correlation Mechanism between Dominant Bacteria and Primary Metabolites during Fermentation of Red Sour Soup" Foods 11, no. 3: 341. https://doi.org/10.3390/foods11030341