
Variation in Cell Wall Metabolism and Flesh Firmness of Four Apple Cultivars during Fruit Development

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Measurement of Physiological Characteristics
2.3. Analysis of CWMs
2.4. Determination of Hydrolytic Enzyme Activity
2.5. The Microstructure of Fruit Pulp Cells
2.6. Morphology Analysis of Apple Fruit Cell Wall
2.7. Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
2.8. Statistical Analysis
3. Results
3.1. Physiological Characteristics of ‘Ruixue’ and ‘Ruixianghong’ and Their Parents Apples during Fruit Development
3.2. Measurement of CWMs in the Four Apple Cultivars during Development
3.3. Measurement of Cell Wall Degrading Enzymes Activity in the Four Apple Cultivars during Development
3.4. Microstructure Analysis in the Four Apple Cultivars during Development
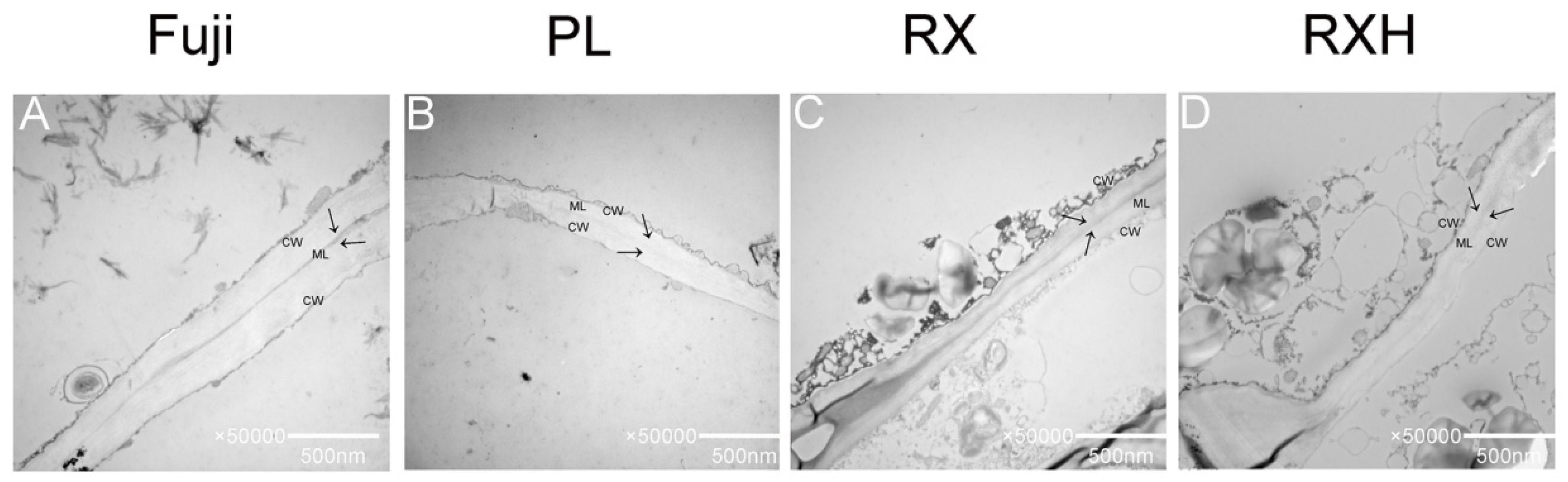
3.5. Ultrastructural Analysis of the Four Apple Cultivars during the Mature Stage
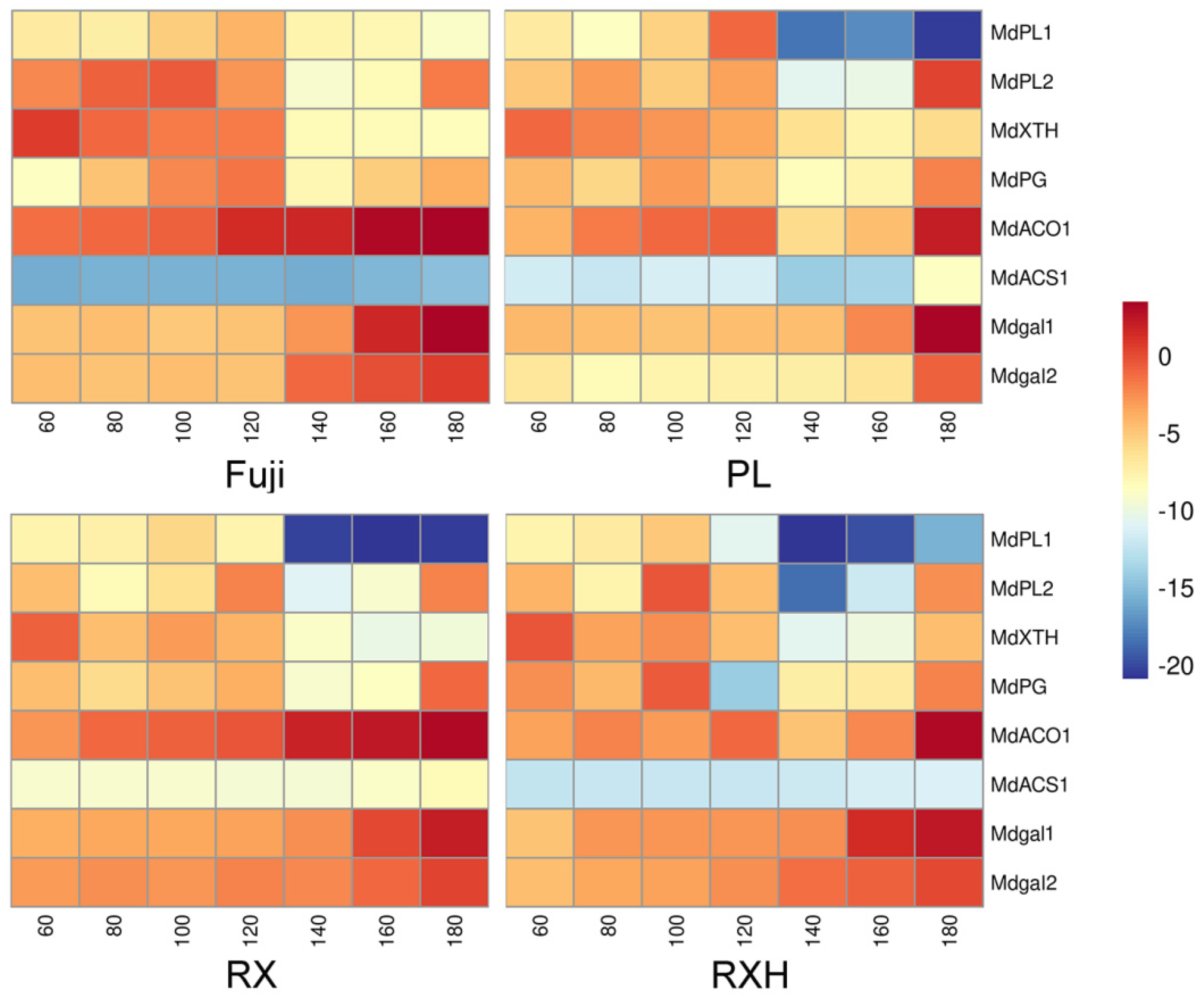
3.6. Expression of Cell Wall and Ethylene-Related Genes in the Four Apple Cultivars during Development
4. Discussion
4.1. Dynamic Variations of Physiological Characteristics of ‘Ruixue’ and ‘Ruixianghong’ Apples and Their Parent Apples during Fruit Development
4.2. Analysis of Changes in Cell Wall Components during Development in Four Cultivars
4.3. The Impact of Cell Wall Hydrolase of Softening on the Various Apple Cultivars during the Ripening Process
4.4. Microstructural Alterations of Fruit Firmness during Development in the Four Cultivars
4.5. Cell Wall Ultrastructure in the Different Apple Cultivars at Maturity
4.6. Analysis of Expression Patterns of Ethylene Synthesis and Hydrolysis Enzyme-Related Genes during Fruit Development Involved in Cell Wall Metabolism Pathways
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dheilly, E.; Gall, S.L.; Guillou, M.C.; Renou, J.P.; Bonnin, E.; Orsel, M.; Lahaye, M. Cell wall dynamics during apple development and storage involves hemicellulose modifications and related expressed genes. BMC Plant Biol. 2016, 16, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Cong, P.; He, J.; Bu, H.; Qin, S.; Lyu, D. Differential pulp cell wall structures lead to diverse fruit textures in apple (Malus domestica). Protoplasma 2022, 259, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Verhage, L. Apples and EARs: The role of an EAR domain in apple fruit firmness. Plant J. 2020, 103, 935–936. [Google Scholar] [CrossRef] [PubMed]
- Gwanpua, S.G.; van Buggenhout, S.; Verlinden, B.E.; Christiaens, S.; Shpigelman, A.; Vicent, V.; Kermani, Z.J.; Nicolai, B.M.; Hendrickx, M.; Geeraerd, A. Pectin modifications and the role of pectin-degrading enzymes during postharvest softening of Jonagold apples. Food Chem. 2014, 158, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Uluisik, S.; Chapman, N.H.; Smith, R.; Poole, M.; Adams, G.; Gillis, R.B.; Besong, T.M.D.; Sheldon, J.; Stiegelmeyer, S.; Perez, L.J.N.B. Genetic improvement of tomato by targeted control of fruit softening. Nat. Biotechnol. 2016, 34, 950–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Sun, S.; Wang, H.J.; Wang, K.T.; Yu, H.; Zhou, Z.; Xin, P.Y.; Chu, J.F.; Zhao, T.M.; Wang, H.Z.; et al. FIS1 encodes a GA2-oxidase that regulates fruit firmness in tomato. Nat. Commun. 2020, 11, 5844. [Google Scholar] [CrossRef]
- Yang, S.; Meng, Z.; Fan, J.; Yan, L.; Yang, Y.; Zhao, Z. Evaluation of the volatile profiles in pulp of 85 apple cultivars (Malus domestica) by HS–SPME combined with GC–MS. J. Food Meas. Charact. 2021, 15, 4215–4225. [Google Scholar] [CrossRef]
- Cybulska, J.; Zdunek, A.; Psonka-Antonczyk, K.M.; Stokke, B.T. The relation of apple texture with cell wall nanostructure studied using an atomic force microscope. Carbohydr. Polym. 2013, 92, 128–137. [Google Scholar] [CrossRef]
- Brummell, D.A.; Harpster, M.H. Cell wall metabolism in fruit softening and quality and its manipulation in transgenic plants. Plant Mol. Biol. 2001, 47, 311–340. [Google Scholar] [CrossRef]
- Wang, D.; Yeats, T.H.; Uluisik, S.; Rose, J.K.C.; Seymour, G.B. Fruit softening: Revisiting the role of pectin. Trends Plant Sci. 2018, 23, 302–310. [Google Scholar] [CrossRef]
- Leszczuk, A.; Chylińska, M.; Zdunek, A. Distribution of arabinogalactan proteins and pectins in the cells of apple (Malus × domestica) fruit during post-harvest storage. Ann. Bot. 2019, 123, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ornelas-Paz, J.; Quintana-Gallegos, B.M.; Escalante-Minakata, P.; Reyes-Hernandez, J.; Perez-Martinez, J.D.; Rios-Velasco, C.; Ruiz-Cruz, S. Relationship between the firmness of Golden Delicious apples and the physicochemical characteristics of the fruits and their pectin during development and ripening. J. Food Sci. Technol. 2018, 55, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Redgwell, R.J.; MacRae, E.; Hallett, I.; Fischer, M.; Perry, J.; Harker, R. In vivo and in vitro swelling of cell walls during fruit ripening. Planta 1997, 203, 162–173. [Google Scholar] [CrossRef]
- Kinnaert, C.; Daugaard, M.; Nami, F.; Clausen, M.H. Chemical synthesis of oligosaccharides related to the cell walls of plants and algae. Chem. Rev. 2017, 117, 11337–11405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volz, R.K.; Harker, F.R.; Lang, S. Firmness decline in ‘Gala’ apple during fruit development. J. Am. Soc. Hortic. Sci. 2003, 128, 797–802. [Google Scholar] [CrossRef]
- Goulao, L.F.; Santos, J.; de Sousa, I.; Oliveira, C.A. Patterns of enzymatic activity of cell wall-modifying enzymes during growth and ripening of apples. Postharvest Biol. Technol. 2007, 43, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Min, L.; Yanmin, Z.; Zongying, Z.; Xiaohao, J.; Rui, Z.; Daliang, L.; Liping, G.; Jing, Z.; Biao, W.; Yusen, W. Hypersensitive ethylene signaling and ZMdPG1 expression lead to fruit softening and dehiscence. PLoS ONE 2013, 8, e58745. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.; Peace, C.P.; Stella, S.; Serra, S.; Musacchi, S.; Bazzani, M.; Sansavini, S.; van de Weg, W.E. QTL dynamics for fruit firmness and softening around an ethylene-dependent polygalacturonase gene in apple (Malus × domestica Borkh.). J. Exp. Bot. 2010, 61, 3029–3039. [Google Scholar] [CrossRef] [Green Version]
- Moriya, S.; Kunihisa, M.; Okada, K.; Iwanami, H.; Abe, K. Identification of QTLs for flesh mealiness in apple (Malus × domestica Borkh.). Hortic. J. 2017, 86, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Wang, X.J.; Tan, G.F.; Zhou, W.Q.; Wang, G.L. Gibberellin and the plant growth retardant Paclobutrazol altered fruit shape and ripening in tomato. Protoplasma 2020, 257, 853–861. [Google Scholar] [CrossRef]
- Kou, X.; Feng, Y.; Yuan, S.; Zhao, X.; Wu, C.; Wang, C.; Xue, Z.J. Different regulatory mechanisms of plant hormones in the ripening of climacteric and non-climacteric fruits: A review. Plant Mol. Biol. 2021, 107, 477–497. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.F.; Hoffman, N.E. Ethylene biosynthesis and its regulation in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- Barry, C.S.; Giovannoni, J.J. Ethylene and fruit ripening. Plant Physiol. 2007, 26, 143–159. [Google Scholar] [CrossRef]
- Chang, H.Y.; Tong, C.B.S. Identification of candidate genes involved in fruit ripening and crispness retention through transcriptome analyses of a ‘Honeycrisp’ population. Plants 2020, 9, 1335. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Han, Z.; Sun, Y.; Wang, S.; Wang, T.; Wang, Y.; Xu, K.; Zhang, X.; Xu, X.; Han, Z. ERF4 affects fruit firmness through TPL4 by reducing ethylene production. Plant J. 2020, 103, 937–950. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Shen, F.; Wang, X.; Zheng, W.Y.; Xiao, C.; Deng, Y.; Wang, T.; Huang, Z.Y.; Zhou, Q.; Wang, Y.; et al. Role of MdERF3 and MdERF118 natural variations in apple flesh firmness/crispness retainability and development of QTL-based genomics-assisted prediction. Plant Biotechnol. J. 2021, 19, 1022–1037. [Google Scholar] [CrossRef]
- Ng, J.K.; Schröder, R.; Sutherland, P.W.; Hallett, I.; Hall, M.I.; Prakash, R.; Johnston, J.W. Cell wall structures leading to cultivar differences in softening rates develop early during apple (Malus × domestica) fruit growth. BMC Plant Biol. 2013, 13, 183. [Google Scholar] [CrossRef] [Green Version]
- Melton, L.D.; Smith, B.G. Isolation of plant cell walls and fractionation of cell wall polysaccharides. Curr. Protoc. Food Anal. Chem. 2001, E3.1.1–E3.1.23. [Google Scholar] [CrossRef]
- Brummell, D.A. Cell wall disassembly in ripening fruit. Funct. Plant Biol. 2006, 33, 103–119. [Google Scholar] [CrossRef]
- Ge, Y.; Bin, D.; Li, C.; Wei, M.; Chen, Y.; Li, X. Application of sodium silicate retards apple softening by suppressing the activity of enzymes related to cell wall degradation. J. Sci. Food Agric. 2019, 99, 1828–1833. [Google Scholar] [CrossRef]
- Chen, Y.; Hung, Y.C.; Chen, M.; Lin, H. Effects of acidic electrolyzed oxidizing water on retarding cell wall degradation and delaying softening of blueberries during postharvest storage. LWT 2017, 84, 650–657. [Google Scholar] [CrossRef]
- Li, Y.; He, H.; Hou, Y.; Kelimu, A.; Wu, F.; Zhao, Y.; Shi, L.; Zhu, X. Salicylic acid treatment delays apricot (Prunus armeniaca L.) fruit softening by inhibiting ethylene biosynthesis and cell wall degradation. Sci. Hortic. 2022, 300, 111061. [Google Scholar] [CrossRef]
- Wang, H.; Wang, S.; Fan, M.M.; Zhang, S.H.; Sun, L.L.; Zhao, Z.Y. Metabolomic insights into the browning of the peel of bagging ‘Rui Xue’ apple fruit. BMC Plant Biol. 2021, 21, 209. [Google Scholar] [CrossRef] [PubMed]
- Li, H.G.; Liu, J.L.; Zhang, X.; Zhu, Z.Z.; Yang, H.J.; Dang, M.L.; Zhao, Z.Y. Comparison of textural and ultrastructural characteristics of four apple cultivars with different textures during cold storage. Int. J. Food Prop. 2019, 22, 659–669. [Google Scholar] [CrossRef]
- Shi, Y.; Vrebalov, J.; Zheng, H.; Xu, Y.; Yin, X.; Liu, W.; Liu, Z.; Sorensen, I.; Su, G.; Ma, Q. A tomato LATERAL ORGAN BOUNDARIES transcription factor, SlLOB1, predominantly regulates cell wall and softening components of ripening. Proc. Nat. Acad. Sci. USA 2021, 118, e2102486118. [Google Scholar] [CrossRef]
- Kuang, J.F.; Wu, C.J.; Guo, Y.F.; Walther, D.; Shan, W.; Chen, J.Y.; Chen, L.; Lu, W.J. Deciphering transcriptional regulators of banana fruit ripening by regulatory network analysis. Plant Biotechnol. J. 2021, 19, 477–489. [Google Scholar] [CrossRef]
- Cheng, C.; Liu, J.; Wang, X.; Wang, Y.; Yuan, Y.; Yang, S. PpERF/ABR1 functions as an activator to regulate PpPG expression resulting in fruit softening during storage in peach (Prunus persica). Postharvest Biol. Technol. 2022, 189, 111919. [Google Scholar] [CrossRef]
- Song, L.Y.; Wang, Z.G.; Wang, Z.M.; Meng, G.; Zhai, R.; Cai, M.; Ma, F.W.; Xu, L.F. Screening of cell wall-related genes that are expressed differentially during ripening of pears with different softening characteristics. Postharvest Biol. Technol. 2016, 115, 1–8. [Google Scholar] [CrossRef]
- Yang, H.; Liu, J.; Dang, M.; Bo, Z.; Li, H. Analysis of β-Galactosidase during fruit development and ripening in two different texture types of apple cultivars. Front. Plant Sci. 2018, 9, 539. [Google Scholar] [CrossRef]
- Xie, F.; Yuan, S.; Pan, H.; Wang, R.; Cao, J.; Jiang, W. Effect of yeast mannan treatments on ripening progress and modification of cell wall polysaccharides in tomato fruit. Food Chem. 2017, 218, 509–517. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, Y.; Lin, H.; Lin, M.; Li, H.; Yuan, F.; Chen, Y.; Xiao, J. Effects of paper containing 1-MCP postharvest treatment on the disassembly of cell wall polysaccharides and softening in Younai plum fruit during storage. Food Chem. 2018, 264, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Ma, F.; Shi, S.; Qi, X.; Zhu, X.; Yuan, J. Changes and postharvest regulation of activity and gene expression of enzymes related to cell wall degradation in ripening apple fruit. Postharvest Biol. Technol. 2010, 56, 147–154. [Google Scholar] [CrossRef]
- Atkinson, R.G.; Sutherland, P.W.; Johnston, S.L.; Gunaseelan, K.; Hallett, I.C.; Mitra, D.; Brummell, D.A.; Schroder, R.; Johnston, J.W.; Schaffer, R.J. Down-regulation of POLYGALACTURONASE1 alters firmness, tensile strength and water loss in apple (Malus × domestica) fruit. BMC Plant Biol. 2012, 12, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Ren, S.; Lin, H.; Ni, L.I.; Lin, Y.; Zhuo, S. Characteristics of cell wall metabolism of harvested Chinese olive fruits with different maturities in response to chilling injury during cold storage. Food Sci. 2018, 39, 229–235. [Google Scholar] [CrossRef]
- Allan-Wojtas, P.; Sanford, K.; McRae, K.; Carbyn, S. An integrated microstructural and sensory approach to describe apple texture. J. Am. Soc. Hortic. Sci. 2003, 128, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.L.; Bennett, A.B. Role of cell wall hydrolases in fruit ripening. Annu. Rev. Plant Physiol. Mol. Biol. 1991, 42, 675–703. [Google Scholar] [CrossRef]
- Tong, C.; Krueger, D.; Vickers, Z.; Bedford, D.; Luby, J.; El-Shiekh, A.; Shackel, K.; Ahmadi, H. Comparison of softening-related changes during storage of ‘Honeycrisp’ apple, its parents, and ‘Delicious’. J. Am. Soc. Hortic. Sci. 1999, 124, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Xu, R.; Fang, Q.; Yuan, Y.; Cao, J.; Jiang, W. Analyses of microstructure and cell wall polysaccharides of flesh tissues provide insights into cultivar difference in mealy patterns developed in apple fruit. Food Chem. 2020, 321, 126707. [Google Scholar] [CrossRef]
- Shi, Z.; Yang, H.; Jiao, J.; Wang, F.; Lu, Y.; Deng, J. Effects of graft copolymer of chitosan and salicylic acid on reducing rot of postharvest fruit and retarding cell wall degradation in grapefruit during storage. Food Chem. 2019, 283, 92–100. [Google Scholar] [CrossRef]
- Tacken, E.; Ireland, H.; Gunaseelan, K.; Karunairetnam, S.; Wang, D.; Schultz, K.; Bowen, J.; Atkinson, R.G.; Johnston, J.W.; Putterill, J.; et al. The role of ethylene and cold temperature in the regulation of the apple POLYGALACTURONASE1 gene and fruit softening. Plant Physiol. 2010, 153, 294–305. [Google Scholar] [CrossRef]
- Smith, D.L.; Abbott, J.A.; Gross, K.C. Down-regulation of tomato beta-galactosidase 4 results in decreased fruit softening. Plant Physiol. 2002, 129, 1755–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunako, T.; Sakuraba, W.; Senda, M.; Akada, S.; Ishikawa, R.; Niizeki, M.; Harada, T. An allele of the ripening-specific 1-aminocyclopropane-1-carboxylic acid synthase gene (ACS1) in apple fruit with a long storage life. Plant Physiol. 1999, 119, 1297–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Liu, J.; Zang, N.; Yin, Z.; Wang, A. Effects of calcium application on apple fruit softening during storage revealed by proteomics and phosphoproteomics. Hortic. Plant J. 2022, 8, 15. [Google Scholar] [CrossRef]
- Soria-Guerra, R.E.; Rosales-Mendoza, S.; Gasic, K.; Wisniewski, M.E.; Band, M.; Korban, S.S. Gene expression is highly regulated in early developing fruit of apple. Plant Mol. Biol. Rep. 2011, 29, 885–897. [Google Scholar] [CrossRef]
- Zhu, Y.M.; Zheng, P.; Varanasi, V.; Shin, S.B.; Main, D.; Curry, E.; Mattheis, J.P. Multiple plant hormones and cell wall metabolism regulate apple fruit maturation patterns and texture attributes. Tree Genet. Genomes 2012, 8, 1389–1406. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Q.; Li, X.; Wang, L.; Wang, B.; Feng, Y.; Yang, H.; Zhao, Z. Variation in Cell Wall Metabolism and Flesh Firmness of Four Apple Cultivars during Fruit Development. Foods 2022, 11, 3518. https://doi.org/10.3390/foods11213518
Su Q, Li X, Wang L, Wang B, Feng Y, Yang H, Zhao Z. Variation in Cell Wall Metabolism and Flesh Firmness of Four Apple Cultivars during Fruit Development. Foods. 2022; 11(21):3518. https://doi.org/10.3390/foods11213518
Chicago/Turabian StyleSu, Qiufang, Xianglu Li, Lexing Wang, Bochen Wang, Yifeng Feng, Huijuan Yang, and Zhengyang Zhao. 2022. "Variation in Cell Wall Metabolism and Flesh Firmness of Four Apple Cultivars during Fruit Development" Foods 11, no. 21: 3518. https://doi.org/10.3390/foods11213518