
Potential Health Benefits of Yeast-Leavened Bread Containing LAB Pediococcus pentosaceus Fermented Pitaya (Hylocereus undatus): Both In Vitro and In Vivo Aspects
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and methods
2.1. Materials, Chemicals and Microorganisms
2.2. Pitaya Fermentation and Bread Preparation
2.2.1. pH and Total Titratable Acidity (TTA) of Fermented Pitaya
2.2.2. Sugar Composition of Fermented Pitaya
2.2.3. Determination of Total Phenolic Content (TPC), Total Flavonoid Content (TFC), and Phenolic Acids of Fermented Pitaya
2.2.4. Determination of α-Amylase Activity and β-Glucosidase Activity in Dough
2.2.5. Changes in the Dietary Fiber Content following the Dough Proofing
2.2.6. In Vitro Starch Digestibility of the Bread
2.3. Customized Bread Diets, Animal and Study Design
2.3.1. Preparation of Customized Bread Diet
2.3.2. Animal Study Design
2.4. Determination of Changes in Gut Microbiota Composition
2.5. Analysis of the Short-Chain Fatty Acids (SCFAs) Content in Feces and Serum
2.6. Analysis of the Bio-Parameters in the Serum Samples
2.7. Statistical Analysis
3. Results and Discussion
3.1. Effect of the Fermentation on Sugar and Phytochemical Content of the Pitaya Substrate
3.2. Bioactive Components of the Dough and Its Corresponding Bread Supplemented with Pitaya Fermentates
3.2.1. Beta-Glucosidase and α-Amylase Activity in Dough
3.2.2. Difference in the Dietary Fiber Content among the Dough Samples
3.2.3. In Vitro Starch Digestibility of the Bread
3.2.4. Difference in the Bioactive Components among the Breads
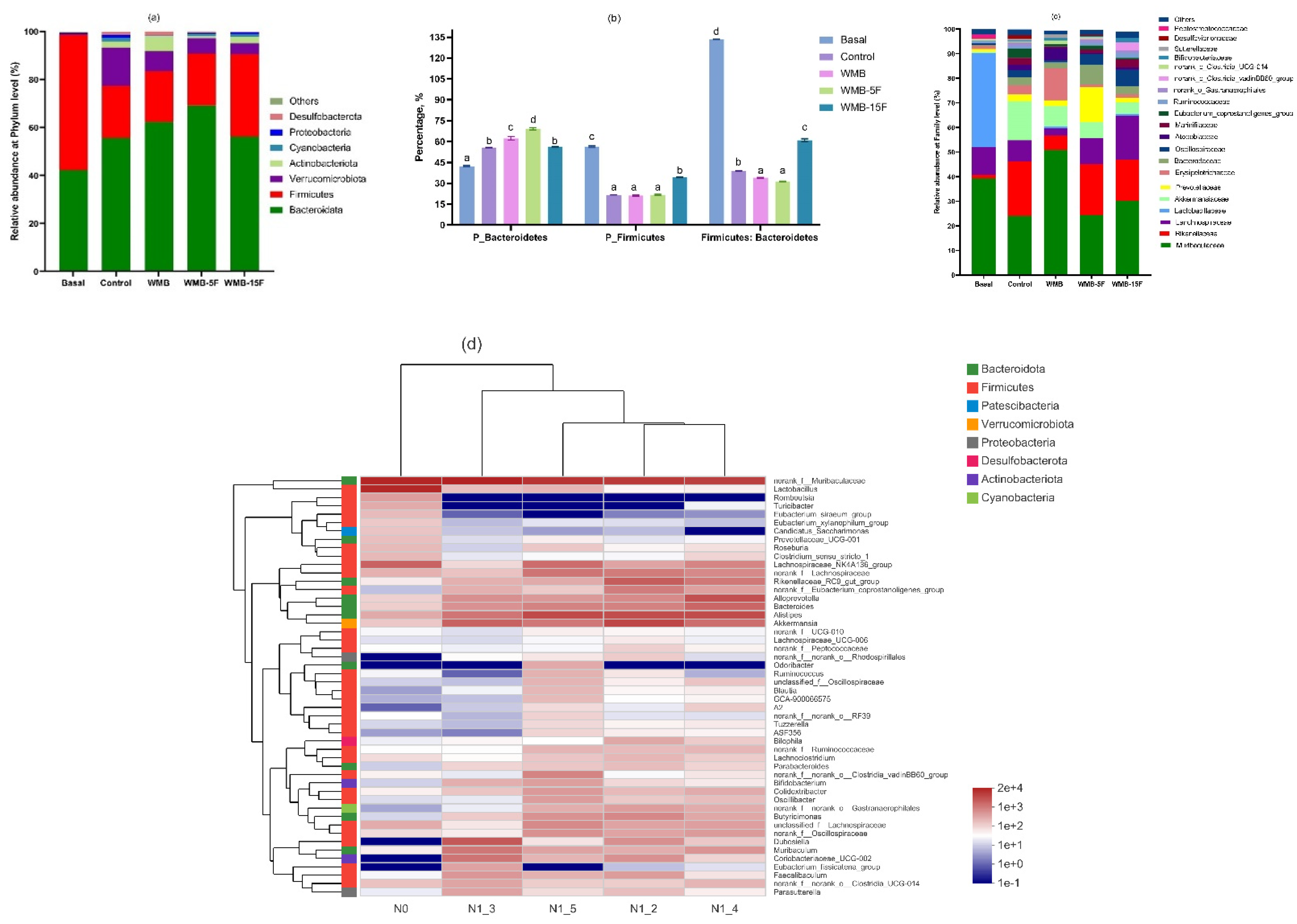
3.3. Effect of Diet Based on Bread Supplemented with Fermented Pitaya on Gut Microbiota in Mice
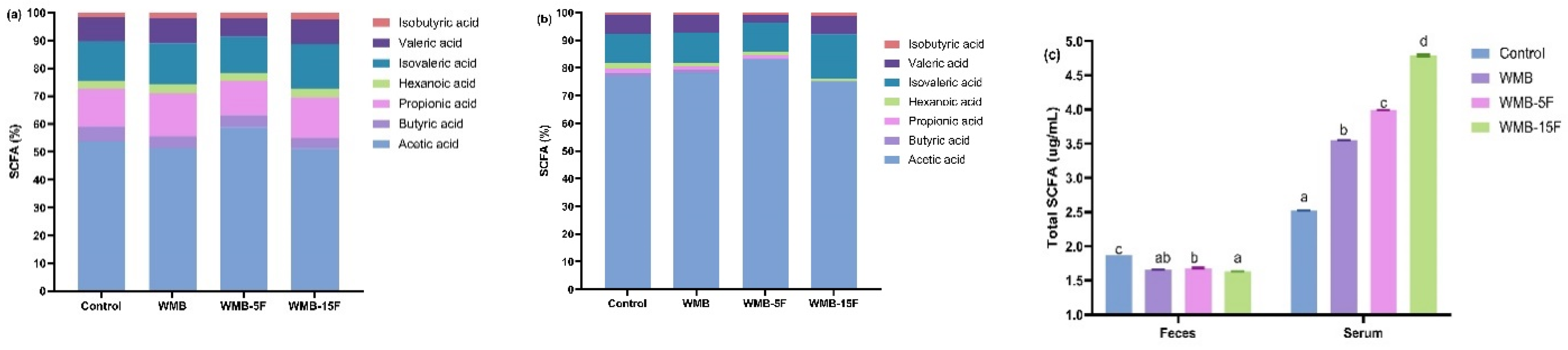
3.4. Effect of Diet Based on Bread Supplemented with Fermented Pitaya on SCFAs Production in Serum and Feces of the Mice
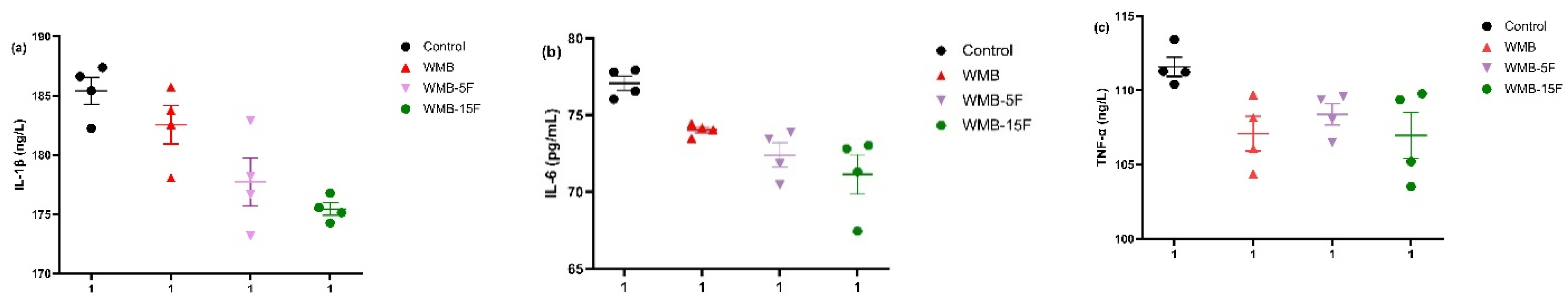
3.5. Effect of Diet Based on Bread Supplemented with Fermented Pitaya on the Serum Levels of Pro-Inflammatory Cytokines
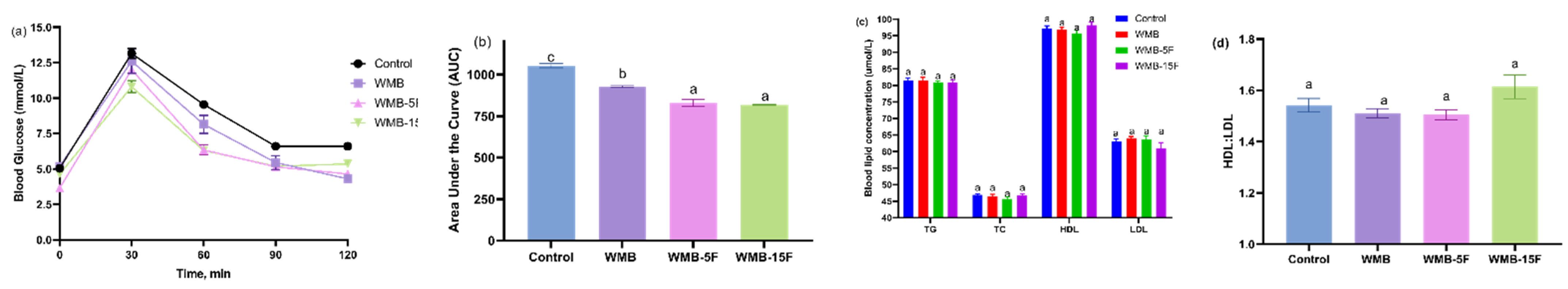
3.6. Effect of Diet Based on Bread Supplemented with Fermented Pitaya on Glucose Tolerance and Serum Lipid Profile
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poutanen, K.; Flander, L.; Katina, K. Sourdough and cereal fermentation in a nutritional perspective. Food Microbiol. 2009, 26, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Arora, K.; Ameur, H.; Polo, A.; Di Cagno, R.; Rizzello, C.G.; Gobbetti, M. Thirty years of knowledge on sourdough fermentation: A systematic review. Trends Food Sci. Technol. 2021, 108, 71–83. [Google Scholar] [CrossRef]
- Zhang, B.; Omedi, J.O.; Zheng, J.; Huang, W.; Jia, C. Exopolysaccharides in sourdough fermented by Weissella confusa QS813 protected protein matrix and quality of frozen gluten-red bean dough during freeze-thaw cycles. Food Biosci. 2021, 43, 101180. [Google Scholar] [CrossRef]
- Siepmann, F.B.; Ripari, V.; Waszczynskyj, N.; Spier, M.R. Overview of Sourdough Technology: From Production to Marketing. Food Bioprocess Technol. 2018, 11, 242–270. [Google Scholar] [CrossRef]
- Slavin, J.L.; Lloyd, B. Health Benefits of Fruits and Vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Suh, D.H.; Lee, S.; Heo, D.Y.; Kim, Y.; Cho, S.K.; Lee, L.; Lee, C.H. Metabolite profiling of red and white pitayas (Hylocereus polyrhizus and Hylocereus undatus) for comparing betalain biosynthesis and antioxidant activity. J. Agric. Food Chem. 2014, 62, 8764–8771. [Google Scholar] [CrossRef]
- Poolsup, N.; Suksomboon, N.; Paw, N.J. Effect of dragon fruit on glycemic control in prediabetes and type 2 diabetes: A systematic review and meta-analysis. PLoS ONE 2017, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Holanda, M.O.; Lira, S.M.; da Silva, J.Y.G.; Marques, C.G.; Coelho, L.C.; Lima, C.L.S.; Costa, J.T.G.; da Silva, G.S.; Santos, G.B.M.; Zocolo, G.J.; et al. Intake of pitaya (Hylocereus polyrhizus (F.A.C. Weber) Britton & Rose) beneficially affects the cholesterolemic profile of dyslipidemic C57BL/6 mice. Food Biosci. 2021, 42, 101181. [Google Scholar]
- Song, H.; Zheng, Z.; Wu, J.; Lai, J.; Chu, Q.; Zheng, X. White pitaya (Hylocereus undatus) juice attenuates insulin resistance and hepatic steatosis in diet-induced obese mice. PLoS ONE 2016, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Morais, S.G.G.; da Silva, G.C.B.; dos Santos, M.L.; Martín-Belloso, O.; Magnani, M. Effects of probiotics on the content and bioaccessibility of phenolic compounds in red pitaya pulp. Food Res. Int. 2019, 126, 108681. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and vegetables, as a source of nutritional compounds and phytochemicals: Changes in bioactive compounds during lactic fermentation. Food Res. Int. 2018, 104, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Omedi, J.O.; Huang, W.; Zheng, J. Effect of sourdough lactic acid bacteria fermentation on phenolic acid release and antifungal activity in pitaya fruit substrate. LWT-Food Sci. Technol. 2019, 111, 309–317. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2017, 49, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Min, J.; Lee, J.P.; Baek, S.C.; Kim, S.G.; Jo, Y.D.; Kim, J.; Kim, H. Characterization of two extracellular β-glucosidases produced from the cellulolytic fungus Aspergillus sp. YDJ216 and their potential applications for the hydrolysis of flavone glycosides. Int. J. Biol. Macromol. 2018, 111, 595–603. [Google Scholar]
- Michlmayr, H.; Kneifel, W. β-glucosidase activities of lactic acid bacteria: Mechanisms, impact on fermented food and human health. FEMS Microbiol. Lett. 2014, 352, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Omedi, J.O.; Huang, J.; Huang, W.; Zheng, J.; Zeng, Y. Suitability of pitaya fruit fermented by sourdough LAB strains for bread making: Its impact on dough physicochemical, rheo-fermentation properties and antioxidant, antifungal and quality performance of bread. Heliyon 2021, 7, 08290. [Google Scholar] [CrossRef]
- Hernandez, J.L.; Gonzalez-Castro, M.J.; Alba, I.N.; de la Cruz Garcia, C. High-Performace Liquid Chromatographic Determination of Mono- and Oligosaccharides in Vegetables with Evaporative Light-Scattering Detection and Refractive Index Detection. J. Chromatogr. Sci. 1998, 36, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Guergoletto, K.B.; Costabile, A.; Flores, G.; Garcia, S.; Gibson, G.R. In vitro fermentation of juçara pulp (Euterpe edulis) by human colonic microbiota. Food Chem. 2016, 196, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shang, P.; Qin, F.; Zhou, Q.; Gao, B.; Huang, H.; Yang, H.; Shi, H.; Yu, L. Chemical composition and antioxidative and anti-inflammatory properties of ten commercial mung bean samples. LWT-Food Sci. Technol. 2013, 54, 171–178. [Google Scholar] [CrossRef]
- Liang, L.; Omedi, J.O.; Huang, W.; Zheng, J.; Zeng, Y.; Huang, J.; Zhang, B.; Zhou, L.; Li, N.; Gao, T.; et al. Antioxidant, flavor profile and quality of wheat dough bread incorporated with kiwifruit fermented by β-glucosidase producing lactic acid bacteria strains. Food Biosci. 2021, 46, 101450. [Google Scholar] [CrossRef]
- Wang, Y.C.; Hu, H.F.; Ma, J.W.; Yan, Q.J.; Liu, H.J.; Jiang, Z.Q. A novel high maltose-forming α-amylase from Rhizomucor miehei and its application in the food industry. Food Chem. 2020, 305, 125447. [Google Scholar] [CrossRef] [PubMed]
- AACC. 2000 AACC International Method 32-07.01, 11th ed.; AACC International: St. Paul, MN, USA; Available online: https://www.cerealsgrains.org/resources/Methods/Pages/32Fiber.aspx (accessed on 1 October 2022).
- Rathod, R.P.; Annapure, U.S. Physicochemical properties, protein and starch digestibility of lentil based noodle prepared by using extrusion processing. LWT-Food Sci. Technol. 2017, 80, 121–130. [Google Scholar] [CrossRef]
- Scortichini, S.; Boarelli, M.C.; Silvi, S.; Fiorini, D. Development and validation of a GC-FID method for the analysis of short chain fatty acids in rat and human faeces and in fermentation fluids. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2020, 1143, 121972. [Google Scholar] [CrossRef]
- Tangerman, A.; Nagengast, F.M. A gas chromatographic analysis of fecal short-chain fatty acids, using the direct injection method. Anal. Biochem. 1996, 236, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Zhang, Y.; Wang, L. Structure characteristics of a water-soluble polysaccharide purified from dragon fruit (Hylocereus undatus) pulp. Carbohydr. Polym. 2016, 146, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Delcour, J.A.; Bruneel, C.; Derde, L.J.; Gomand, S.V.; Pareyt, B.; Putseys, J.A.; Wilderjans, E.; Lamberts, L. Fate of Starch in Food Processing: From Raw Materials to Final Food Products. Annu. Rev. Food Sci. Technol. 2010, 1, 87–111. [Google Scholar] [CrossRef]
- Tomas, M.; Rocchetti, G.; Ghisoni, S.; Giuberti, G.; Capanoglu, E.; Lucini, L. Effect of different soluble dietary fibres on the phenolic profile of blackberry puree subjected to in vitro gastrointestinal digestion and large intestine fermentation. Food Res. Int. 2019, 130, 108954. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The controversial role of human gut lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Hinsu, A.T.; Pandit, R.J.; Patel, S.H.; Psifidi, A.; Tomley, F.M.; Da, S.K.; Blake, D.P.; Joshi, C.G. Genome reconstruction of a novel carbohydrate digesting bacterium from the chicken caecal microflora. Meta Gene. 2019, 20, 100543. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhang, K.; Yang, H. Pectin Alleviates High Fat (Lard) Diet-Induced Nonalcoholic Fatty Liver Disease in Mice: Possible Role of Short-Chain Fatty Acids and Gut Microbiota Regulated by Pectin. J. Agric. Food Chem. 2018, 66, 8015–8025. [Google Scholar] [CrossRef] [PubMed]
- Brasili, E.; Hassimotto, N.M.A.; Del Chierico, F.; Marini, F.; Quagliariello, A.; Sciubba, F.; Miccheli, A.; Putignani, L.; Lajolo, F. Daily Consumption of Orange Juice from Citrus sinensis L. Osbeck cv. Cara Cara and cv. Bahia Differently Affects Gut Microbiota Profiling as Unveiled by an Integrated Meta-Omics Approach. J. Agric. Food Chem. 2021, 67, 1381–1391. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Jasso, G.M.; Camacho-Díaz, B.H.; Arenas Ocampo, M.L.; Jiménez-Ferrer, J.E.; Mora-Escobedo, R.; Osorio-Díaz, P. Prebiotic effects of a mixture of agavins and green banana flour in a mouse model of obesity. J. Funct. Foods. 2019, 64, 103685. [Google Scholar] [CrossRef]
- Arias, M.; Cobo, M.; Jaime-Sánchez, P.; Pastor, J.; Marijuan, P.; Pardo, J.; Rezusta, A.; Del Campo, R. Gut microbiota and systemic inflammation changes after bread consumption: The ingredients and the processing influence. J. Funct. Foods. 2017, 32, 98–105. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pitaya Substrate | ||
|---|---|---|
| Characteristic | Before Fermentation | After Fermentation |
| pH | 4.89 ± 0.06 b | 3.74 ± 0.42 a |
| TTA (mL) | 6.5 ± 0.07 a | 7.3 ± 0.07 b |
| Sugars (mg/mL) | ||
| Glucose | 1.88 ± 0.01 a | 2.26 ± 0.00 b |
| Fructose | 5.95 ± 0.00 b | 5.56 ± 0.00 a |
| Maltose | 5.10 ± 0.02 a | 6.40 ± 0.00 b |
| Total | 12.93 ± 0.03 a | 14.22 ± 0.00 b |
| Phytochemical content | ||
| TPC (mg GAE/mL) | 1.25 ± 0.20 a | 1.51 ± 0.20 b |
| TFC (mg RE/g) | 1.48 ± 0.04 a | 1.66 ± 0.04 b |
| Phenolic acids (100 µg/mL) | ||
| Gallic acid | - | 2.37 ± 0.04 |
| Protocatechuic acid | - | 0.76 ± 0.03 |
| Chlorogenic acid | - | 0.52 ± 0.01 |
| Caffeic acid | - | 5 × 10−4 ± 2 × 10−5 |
| Syringic acid | - | 0.51 ± 0.01 |
| p-hydroxybenzoic acid | 3228.89 ± 0.15 b | 2056.31 ± 0.30 a |
| Ferulic acid | 6.19 ± 0.00 b | 1.67 ± 0.00 a |
| Dough Analysis | Bread Analysis | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Enzyme Activity (U/g) | Dietary Fiber Composition (mg/g) | Bioactive Components | In Vitro Starch Digestibility (mg/100 g) | ||||||||||||||
| β-Glucosidase | α-Amylase | IDF | SDF | TDF | Total Sugars (mg/g Bread) | TPC (mg GAE/mL) | TFC (mg RE/g) | SDF (mg/g) | TS | RDS | SDS | ||||||
| 0 min | 90 min | 0 min | 90 min | 0 min | 90 min | 0 min | 90 min | 0 min | 90 min | ||||||||
| Control | 2.17 ± 0.03 aA | 2.30 ± 0.01 bB | 7.34 ± 0.73 aA | 7.63 ± 0.14 aA | 0.097 ± 0.001 a | 0.027 ± 0.001 a | 0.069 ± 0.000 b | 0.025 ± 0.001 a | 0.166 ± 0.001 b | 0.053 ± 0.000 a | 7.09 ± 0.08 a | 1.65 ± 0.07 | 2.28 ± 0.01 a | 0.043 ± 0.000 b | 424.57 ± 11.78 d | 411.54 ± 12.03 e | 13.03 ± 0.25 a |
| WMB | 2.47 ± 0.01 cB | 2.20 ± 0.01 aA | 8.42 ± 0.08 bA | 8.57 ± 0.12 bA | 0.105 ± 0.001 c | 0.028 ± 0.001 a | 0.073 ± 0.001 b | 0.044 ± 0.005 b | 0.178 ± 0.000 c | 0.072 ± 0.005 b | 8.16 ± 0.03 ab | 1.83 ± 0.13 | 2.29 ± 0.04 a | 0.043 ± 0.000 b | 213.19 ± 24.09 a | 197.84 ± 24.09 a | 15.35 ± 0.00 a |
| WMB-5F | 2.12 ± 0.01 aA | 2.22 ± 0.01 aB | 8.26 ± 0.19 bA | 8.99 ± 0.01 cB | 0.100 ± 0.000 b | 0.051 ± 0.001 d | 0.045 ± 0.002 a | 0.051 ± 0.001 c | 0.146 ± 0.002 a | 0.103 ± 0.002 c | 9.14 ± 0.54 b | 1.74 ± 0.07 | 2.45 ± 0.01 b | 0.045 ± 0.000 c | 263.08 ± 2.12 b | 247.37 ± 0.60 b | 15.71 ± 1.51 a |
| WMB-15F | 2.39 ± 0.03 bA | 2.33 ± 0.02 cA | 8.48 ± 0.07 bA | 9.72 ± 0.11 dB | 0.104 ± 0.001 c | 0.037 ± 0.001 b | 0.067 ± 0.001 b | 0.097 ± 0.002 d | 0.171 ± 0.001 c | 0.134 ± 0.003 d | 9.20 ± 0.91 b | 1.71 ± 0.11 | 2.54 ± 0.03 b | 0.046 ± 0.000 d | 307.75 ± 17.01 c | 285.61 ± 18.53 c | 22.13 ± 1.51 b |
| WMB-25F | 2.57 ± 0.05 dB | 2.46 ± 0.00 dA | 10.22 ± 0.06 cA | 10.69 ± 0.20 eB | 0.105 ± 0.000 c | 0.042 ± 0.000 c | 0.072 ± 0.005 b | 0.142 ± 0.004 e | 0.177 ± 0.004 c | 0.184 ± 0.004 e | 8.72 ± 0.16 b | 1.79 ± 0.06 | 2.51 ± 0.11 b | 0.042 ± 0.000 a | 329.30 ± 0.78 c | 294.67 ± 1.79 d | 34.63 ± 1.01 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omedi, J.O.; Li, N.; Chen, C.; Cheng, X.; Huang, J.; Zhang, B.; Gao, T.; Liang, L.; Zhou, Z.; Huang, W. Potential Health Benefits of Yeast-Leavened Bread Containing LAB Pediococcus pentosaceus Fermented Pitaya (Hylocereus undatus): Both In Vitro and In Vivo Aspects. Foods 2022, 11, 3416. https://doi.org/10.3390/foods11213416
Omedi JO, Li N, Chen C, Cheng X, Huang J, Zhang B, Gao T, Liang L, Zhou Z, Huang W. Potential Health Benefits of Yeast-Leavened Bread Containing LAB Pediococcus pentosaceus Fermented Pitaya (Hylocereus undatus): Both In Vitro and In Vivo Aspects. Foods. 2022; 11(21):3416. https://doi.org/10.3390/foods11213416
Chicago/Turabian StyleOmedi, Jacob Ojobi, Ning Li, Cheng Chen, Xin Cheng, Jing Huang, Binle Zhang, Tiecheng Gao, Li Liang, Zhongkai Zhou, and Weining Huang. 2022. "Potential Health Benefits of Yeast-Leavened Bread Containing LAB Pediococcus pentosaceus Fermented Pitaya (Hylocereus undatus): Both In Vitro and In Vivo Aspects" Foods 11, no. 21: 3416. https://doi.org/10.3390/foods11213416