
Ropiness in Bread—A Re-Emerging Spoilage Phenomenon
 , ,
, ,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. The Route of Endospores into the Bakery Environment
3. The Diversity of Species Inducing Rope Spoilage in Bread
4. Rope-Inducing Potential (RIP)
4.1. Methods of RIP Assessment
4.2. Factors Influencing RIP
5. Prevention of Rope Formation in Bread
6. Food Safety Concerns
7. Considerations for the Future
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Los, A.; Ziuzina, D.; Boehm, D.; Bourke, P. Effects of cold plasma on wheat grain microbiome and antimicrobial efficacy against challenge pathogens and their resistance. Int. J. Food Microbiol. 2020, 335, 108889. [Google Scholar] [CrossRef] [PubMed]
- Los, A.; Ziuzina, D.; Bourke, P. Current and Future Technologies for Microbiological Decontamination of Cereal Grains. J. Food Sci. 2018, 83, 1484–1493. [Google Scholar] [CrossRef] [PubMed]
- Arranz-Otaegui, A.; Carretero, L.G.; Ramsey, M.N.; Fuller, D.Q.; Richter, T. Archaeobotanical evidence reveals the origins of bread 14,400 years ago in northeastern Jordan. Proc. Natl. Acad. Sci. USA 2018, 115, 7925–7930. [Google Scholar] [CrossRef]
- Heiss, A.G.; Pouget, N.; Wiethold, J.; Delor-Ahü, A.; Le Goff, I. Tissue-based analysis of a charred flat bread (galette) from a Roman cemetery at Saint-Memmie (Dép. Marne, Champagne-Ardenne, north-eastern France). J. Archaeol. Sci. 2015, 55, 71–82. [Google Scholar] [CrossRef]
- Piperno, D.R.; Weiss, E.; Holst, I.; Nadel, D. Processing of wild cereal grains in the Upper Palaeolithic revealed by starch grain analysis. Nature 2004, 430, 670–673. [Google Scholar] [CrossRef]
- Smith, J.P.; Daifas, D.P.; El-Khoury, W.; Koukoutsis, J.; El-Khoury, A. Shelf Life and Safety Concerns of Bakery Products—A Review. Crit. Rev. Food Sci. Nutr. 2004, 44, 19–55. [Google Scholar] [CrossRef]
- Amos, A.J.; Kent-Jones, D.W. The “rope” spore content of flour and its significance. Analyst 1931, 56, 572–586. [Google Scholar] [CrossRef]
- Cauvain, S. Technology of Breadmaking, 3rd ed.; Springer International Publishing: Cham, Switzerland, 2015; ISBN 978-3-319-14686-7. [Google Scholar]
- Saranraj, P.; Sivasakthivelan, P. Microorganisms involved in spoilage of bread and its control measures. In Bread and Its Fortification: Nutrition and Health Benefits, 1st ed.; Rosell, C.M., Bajerska, J., El Sheikha, A.F., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 132–149. ISBN 9780429172557. [Google Scholar]
- Cohn, E.J.; Wolbach, S.B.; Henderson, L.J.; Cathcart, P.H. On the control of rope in bread. J. Gen. Physiol. 1918, 1, 221–230. [Google Scholar] [CrossRef]
- Watkins, E.J. Ropiness in flour and bread and its detection and prevention. J. Soc. Chem. Ind. 1906, 25, 350–357. [Google Scholar] [CrossRef]
- Cook, F.K.; Johnson, B.L. Microbiological Spoilage of Cereal Products. In Compendium of the Microbiological Spoilage of Foods and Beverages; Sperber, W.H., Doyle, M.P., Eds.; Springer: New York, NY, USA; London, UK, 2009; pp. 223–244. ISBN 978-1-4419-0825-4. [Google Scholar]
- Axel, C.; Zannini, E.; Arendt, E.K. Mold spoilage of bread and its biopreservation: A review of current strategies for bread shelf life extension. Crit. Rev. Food Sci. Nutr. 2017, 57, 3528–3542. [Google Scholar] [CrossRef]
- Rahman, M.; Simsek, S. Go clean label: Replacement of commercial dough strengtheners with hard red spring wheat flour in bread formulations. J. Food Sci. Technol. 2020, 57, 3581–3590. [Google Scholar] [CrossRef] [PubMed]
- Logan, N.A.; De Vos, P. Bacillus. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–163. ISBN 9781118960608. [Google Scholar]
- Postollec, F.; Mathot, A.-G.; Bernard, M.; Divanac’H, M.-L.; Pavan, S.; Sohier, D. Tracking spore-forming bacteria in food: From natural biodiversity to selection by processes. Int. J. Food Microbiol. 2012, 158, 1–8. [Google Scholar] [CrossRef]
- Christie, G.; Setlow, P. Bacillus spore germination: Knowns, unknowns and what we need to learn. Cell. Signal. 2020, 74, 109729. [Google Scholar] [CrossRef]
- Khanna, K.; Lopez-Garrido, J.; Pogliano, K. Shaping an Endospore: Architectural Transformations During Bacillus subtilis Sporulation. Annu. Rev. Microbiol. 2020, 74, 361–386. [Google Scholar] [CrossRef] [PubMed]
- Lavermicocca, P.; Valerio, F.; DeBellis, P.; Sisto, A.; Leguérinel, I. Chapter 16—Sporeforming bacteria associated with bread production: Spoilage and toxigenic potential. In Food Hygiene and Toxicology in Ready to Eat Foods; Kotzekidou, P., Ed.; Academic Press (Elsevier): Amsterdam, The Netherlands, 2016; pp. 275–293. ISBN 978-0-12-801916-0. [Google Scholar]
- André, S.; Vallaeys, T.; Planchon, S. Spore-forming bacteria responsible for food spoilage. Res. Microbiol. 2017, 168, 379–387. [Google Scholar] [CrossRef]
- Farmiloe, F.J.; Cornford, S.J.; Coppock, J.B.M.; Ingram, M. The survival of Bacillus subtilis spores in the baking of bread. J. Sci. Food Agric. 1954, 5, 292–304. [Google Scholar] [CrossRef]
- Thompson, J.M.; Dodd, C.E.; Waites, W.M. Spoilage of bread by bacillus. Int. Biodeterior. Biodegrad. 1993, 32, 55–66. [Google Scholar] [CrossRef]
- Thompson, J.; Waites, W.; Dodd, C. Detection of rope spoilage in bread caused by Bacillus species. J. Appl. Microbiol. 1998, 85, 481–486. [Google Scholar] [CrossRef]
- Rosenkvist, H. Contamination profiles and characterisation of Bacillus species in wheat bread and raw materials for bread production. Int. J. Food Microbiol. 1995, 26, 353–363. [Google Scholar] [CrossRef]
- Pepe, O.; Blaiotta, G.; Moschetti, G.; Greco, T.; Villani, F. Rope-Producing Strains of Bacillus spp. from Wheat Bread and Strategy for Their Control by Lactic Acid Bacteria. Appl. Environ. Microbiol. 2003, 69, 2321–2329. [Google Scholar] [CrossRef] [Green Version]
- Valerio, F.; De Bellis, P.; Di Biase, M.; Lonigro, S.; Giussani, B.; Visconti, A.; Lavermicocca, P.; Sisto, A. Diversity of spore-forming bacteria and identification of Bacillus amyloliquefaciens as a species frequently associated with the ropy spoilage of bread. Int. J. Food Microbiol. 2012, 156, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Rani, U.; Sharma, S.; Kumar, V. Bacillus Species: A Potential Plant Growth Regulator. In Bacilli and Agrobiotechnology: Phytostimulation and Biocontrol; Islam, M.T., Rahman, M.M., Pandey, P., Boehme, M.H., Haesaert, G., Eds.; Springer: Cham, Switzerland, 2019; pp. 29–47. ISBN 978-3-030-15174-4. [Google Scholar]
- Carlin, F. Origin of bacterial spores contaminating foods. Food Microbiol. 2011, 28, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, P. Zerealien und Nährmittel. In Handbuch Lebensmittelhygiene: Praxisleitfaden mit Wissenschaftlichen Grundlagen; Alter, T., Kleer, J., Kley, F., Eds.; Behr: Hamburg, Germany, 2005; pp. XIII.5–XII.6. ISBN 9783899471946. [Google Scholar]
- López, A.M.M.; Simsek, S. Pathogens control on wheat and wheat flour: A review. Cereal Chem. 2020, 98, 17–30. [Google Scholar] [CrossRef]
- Sabillón, L.; Bianchini, A. From Field to Table: A Review on the Microbiological Quality and Safety of Wheat-Based Products. Cereal Chem. 2016, 93, 105–115. [Google Scholar] [CrossRef]
- Berghofer, L.K.; Hocking, A.D.; Miskelly, D.; Jansson, E. Microbiology of wheat and flour milling in Australia. Int. J. Food Microbiol. 2002, 85, 137–149. [Google Scholar] [CrossRef]
- Sabillón, L.; Stratton, J.; Rose, D.J.; Regassa, T.; Bianchini, A. Microbial Load of Hard Red Winter Wheat Produced at Three Growing Environments across Nebraska, USA. J. Food Prot. 2016, 79, 646–654. [Google Scholar] [CrossRef]
- Chauhan, S.; Darvishzadeh, R.; Boschetti, M.; Pepe, M.; Nelson, A. Remote sensing-based crop lodging assessment: Current status and perspectives. ISPRS J. Photogramm. Remote Sens. 2019, 151, 124–140. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, H.; Yi, Y.; Ding, J.; Zhu, M.; Li, C.; Guo, W.; Feng, C.; Zhu, X. Effect of nitrogen levels and nitrogen ratios on lodging resistance and yield potential of winter wheat (Triticum aestivum L.). PLoS ONE 2017, 12, e0187543. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Sinigaglia, M.; Corbo, M.R. Microbial ecology of cereal and cereal-based foods. In Quantitative Microbiology in Food Processing: Modeling the Microbial Ecology; Sant’Ana, A.d.S., Ed.; Wiley: Chichester, UK, 2017; pp. 382–389. ISBN 9781118823071. [Google Scholar]
- Erkmen, O.; Bozoğlu, T.F. Spoilage of Cereals and Cereal Products. In Food Microbiology: Principles into Practice; Erkmen, O., Bozoğlu, T.F., Eds.; John Wiley & Sons Inc.: Chichester West Sussex, UK; Hoboken, NJ, USA, 2016; pp. 364–375. ISBN 9781119237761. [Google Scholar]
- Sabillón, L.; Stratton, J.; Rose, D.; Bianchini, A. Microbiological survey of equipment and wheat-milled fractions of a milling operation. Cereal Chem. 2020, 98, 44–51. [Google Scholar] [CrossRef]
- Cappelli, A.; Oliva, N.; Cini, E. Stone milling versus roller milling: A systematic review of the effects on wheat flour quality, dough rheology, and bread characteristics. Trends Food Sci. Technol. 2020, 97, 147–155. [Google Scholar] [CrossRef]
- Laca, A.; Mousia, Z.; Díaz, M.; Webb, C.; Pandiella, S.S. Distribution of microbial contamination within cereal grains. J. Food Eng. 2006, 72, 332–338. [Google Scholar] [CrossRef]
- Pereira, A.P.M.; Stradiotto, G.C.; Freire, L.; Alvarenga, V.O.; Crucello, A.; Morassi, L.L.; Silva, F.P.; Sant’Ana, A.S. Occurrence and enumeration of rope-producing spore forming bacteria in flour and their spoilage potential in different bread formulations. LWT 2020, 133, 110108. [Google Scholar] [CrossRef]
- Dexter, J.; Wood, P. Recent applications of debranning of wheat before milling. Trends Food Sci. Technol. 1996, 7, 35–41. [Google Scholar] [CrossRef]
- Chen, Y.-X.; Guo, X.-N.; Xing, J.-J.; Sun, X.-H.; Zhu, K.-X. Effects of wheat tempering with slightly acidic electrolyzed water on the microbial, biological, and chemical characteristics of different flour streams. LWT 2020, 118, 108790. [Google Scholar] [CrossRef]
- Serra, S.; De Simeis, D. New insights on the baker’s yeast-mediated hydration of oleic acid: The bacterial contaminants of yeast are responsible for the stereoselective formation of (R)-10-hydroxystearic acid. J. Appl. Microbiol. 2017, 124, 719–729. [Google Scholar] [CrossRef]
- Reale, A.; Di Renzo, T.; Succi, M.; Tremonte, P.; Coppola, R.; Sorrentino, E. Microbiological and Fermentative Properties of Baker’s Yeast Starter Used in Breadmaking. J. Food Sci. 2013, 78, M1224–M1231. [Google Scholar] [CrossRef]
- Gélinas, P. Compressed Baker’s Yeast: Mapping Patents on Post-Fermentation Processes. Compr. Rev. Food Sci. Food Saf. 2017, 16, 456–476. [Google Scholar] [CrossRef]
- Viljoen, B.; Lues, J. The microbial populations associated with post-fermented dough and compressed baker’s yeast. Food Microbiol. 1993, 10, 379–386. [Google Scholar] [CrossRef]
- Viljoen, C.R.; von Holy, A. Microbial populations associated with commercial bread production. J. Basic Microbiol. 1997, 37, 439–444. [Google Scholar] [CrossRef]
- Bailey, C.; Von Holy, A. Bacillus spore contamination associated with commercial bread manufacture. Food Microbiol. 1993, 10, 287–294. [Google Scholar] [CrossRef]
- Collins, N.E.; Ann, L.; Kirschner, M.; von Holy, A. Characterization of Bacillus isolates from ropey bread, bakery equipment and raw materials. S. Afr. J. Sci. 1991, 87, 62–66. [Google Scholar]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust demarcation of 17 distinct Bacillus species clades, proposed as novel Bacillaceae genera, by phylogenomics and comparative genomic analyses: Description of Robertmurraya kyonggiensis sp. nov. and proposal for an emended genus Bacillus limiting it only to the members of the Subtilis and Cereus clades of species. Int. J. Syst. Evol. Microbiol. 2020, 70, 5753–5798. [Google Scholar] [CrossRef] [PubMed]
- Dykes, G.A.; Kirschner, L.M.; von Holy, A. Differentiation of Bacillus isolates from ropey bread and the bakery environment using numerical taxonomy. S. Afr. J. Sci. 1994, 90, 302–307. [Google Scholar]
- Dunlap, C.A.; Bowman, M.J.; Zeigler, D.R. Promotion of Bacillus subtilis subsp. inaquosorum, Bacillus subtilis subsp. spizizenii and Bacillus subtilis subsp. stercoris to species status. Antonie van Leeuwenhoek 2019, 113, 1–12. [Google Scholar] [CrossRef]
- Ash, C.; Priest, F.G.; Collins, M.D. Molecular identification of rRNA group 3 bacilli (Ash, Farrow, Wallbanks and Collins) using a PCR probe test. Proposal for the creation of a new genus Paenibacillus. Antonie van Leeuwenhoek 1994, 64, 253–260. [Google Scholar] [CrossRef]
- Fritze, D.; Pukall, R. Reclassification of bioindicator strains Bacillus subtilis DSM 675 and Bacillus subtilis DSM 2277 as Bacillus atrophaeus. Int. J. Syst. Evol. Microbiol. 2001, 51, 35–37. [Google Scholar] [CrossRef]
- Jia, Y.; Fang, F. Improving applicability of urease from Bacillus amyloliquefaciens JP-21 by site-directed mutagenesis. Chin. J. Biotechnol. 2020, 36, 1640–1649. [Google Scholar] [CrossRef]
- Kim, Y.; Balaraju, K.; Jeon, Y. Biological characteristics of Bacillus amyloliquefaciens AK-0 and suppression of ginseng root rot caused by Cylindrocarpon destructans. J. Appl. Microbiol. 2016, 122, 166–179. [Google Scholar] [CrossRef]
- Leuschner, R.; O’Callaghan, M.; Arendt, E. Bacilli Spoilage in Part-baked and Rebaked Brown Soda Bread. J. Food Sci. 1998, 63, 915–918. [Google Scholar] [CrossRef]
- Kwak, M.-J.; Choi, S.-B.; Ha, S.-M.; Kim, E.H.; Kim, B.-Y.; Chun, J. Genome-based reclassification of Paenibacillus jamilae Aguilera et al. 2001 as a later heterotypic synonym of Paenibacillus polymyxa (Prazmowski 1880) Ash et al. 1994. Int. J. Syst. Evol. Microbiol. 2020, 70, 3134–3138. [Google Scholar] [CrossRef] [PubMed]
- Mols, M.; Abee, T. Role of Ureolytic Activity in Bacillus cereus Nitrogen Metabolism and Acid Survival. Appl. Environ. Microbiol. 2008, 74, 2370–2378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmi, F.M.; Elmitwalli, H.R.; Elnagdy, S.M.; El-Hagrassy, A.F. Calcium carbonate precipitation induced by ureolytic bacteria Bacillus licheniformis. Ecol. Eng. 2016, 90, 367–371. [Google Scholar] [CrossRef]
- Sorokulova, I.; Reva, O.; Smirnov, V.; Pinchuk, I.; Lapa, S.; Urdaci, M. Genetic diversity and involvement in bread spoilage of Bacillus strains isolated from flour and ropy bread. Lett. Appl. Microbiol. 2003, 37, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Vahabi, A.; Ramezanianpour, A.A.; Sharafi, H.; Zahiri, H.S.; Vali, H.; Noghabi, K.A. Calcium carbonate precipitation by strain Bacillus licheniformis AK01, newly isolated from loamy soil: A promising alternative for sealing cement-based materials. J. Basic Microbiol. 2013, 55, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Dhami, N.K.; Mukherjee, A.; Reddy, M.S. Micrographical, minerological and nano-mechanical characterisation of microbial carbonates from urease and carbonic anhydrase producing bacteria. Ecol. Eng. 2016, 94, 443–454. [Google Scholar] [CrossRef]
- Ma, M.; Mu, T.; Zhou, L. Identification of saprophytic microorganisms and analysis of changes in sensory, physicochemical, and nutritional characteristics of potato and wheat steamed bread during different storage periods. Food Chem. 2020, 348, 128927. [Google Scholar] [CrossRef]
- Norris, J.R.; Berkeley, R.C.W.; Logan, N.A.; O’Donnell, A.G.; Starr, M.P. The genera Bacillus and Sporolactobacillus. In The Prokaryotes: A Handbook on Habitats, Isolation, and Identification of Bacteria; Starr, M.P., Stolp, H., Trüper, H.G., Balows, A., Schlegel, H.G., Eds.; Springer: New York, NY, USA, 1981; pp. 1711–1742. ISBN 9780387088716. [Google Scholar]
- Patel, S.; Gupta, R.S. A phylogenomic and comparative genomic framework for resolving the polyphyly of the genus Bacillus: Proposal for six new genera of Bacillus species, Peribacillus gen. nov., Cytobacillus gen. nov., Mesobacillus gen. nov., Neobacillus gen. nov., Metabacillus gen. nov. and Alkalihalobacillus gen. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 406–438. [Google Scholar] [CrossRef]
- Sadeghi, A.; Mortazavi, S.A.; Bahrami, A.R.; Sadeghi, B. Design of multiplex PCR for simultaneous detection of rope-forming Bacillus strains in Iranian bread dough. J. Sci. Food Agric. 2012, 92, 2652–2656. [Google Scholar] [CrossRef]
- Mantzourani, I.; Plessas, S.; Saxami, G.; Alexopoulos, A.; Galanis, A.; Bezirtzoglou, E. Study of kefir grains application in sourdough bread regarding rope spoilage caused by Bacillus spp. Food Chem. 2014, 143, 17–21. [Google Scholar] [CrossRef]
- Biggel, M.; Jessberger, N.; Kovac, J.; Johler, S. Recent paradigm shifts in the perception of the role of Bacillus thuringiensis in foodborne disease. Food Microbiol. 2022, 105, 104025. [Google Scholar] [CrossRef] [PubMed]
- Russ, J.; Reeder, W.; Hatch, D. A rapid bacteriological method for predicting ropiness in bread. Cereal Sci. Today 1961, 6, 89–91. [Google Scholar]
- Volavsek, P.; Kirschner, L.A.M.; von Holy, A. Accelerated methods to predict the rope-inducing potential of bread raw materials. S. Afr. J. Sci. 1992, 88, 99–102. [Google Scholar]
- Li, Z.; Siepmann, F.B.; Tovar, L.E.R.; Chen, X.; Gänzle, M.G. Effect of copy number of the spoVA2mob operon, sourdough and reutericyclin on ropy bread spoilage caused by Bacillus spp. Food Microbiol. 2020, 91, 103507. [Google Scholar] [CrossRef] [PubMed]
- Chhanwal, N.; Anandharamakrishnan, C. Temperature- and Moisture-Based Modeling for Prediction of Starch Gelatinization and Crumb Softness during Bread-Baking Process. J. Texture Stud. 2014, 45, 462–476. [Google Scholar] [CrossRef]
- Berendsen, E.M.; Boekhorst, J.; Kuipers, O.P.; Wells-Bennik, M.H.J. A mobile genetic element profoundly increases heat resistance of bacterial spores. ISME J. 2016, 10, 2633–2642. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, E.M.; Koning, R.A.; Boekhorst, J.; De Jong, A.; Kuipers, O.P.; Wells-Bennik, M.H.J. High-Level Heat Resistance of Spores of Bacillus amyloliquefaciens and Bacillus licheniformis Results from the Presence of a spoVA Operon in a Tn1546 Transposon. Front. Microbiol. 2016, 7, 1912. [Google Scholar] [CrossRef]
- Li, Z.; Schottroff, F.; Simpson, D.J.; Gänzle, M.G. The Copy Number of the spoVA 2mob Operon Determines Pressure Resistance of Bacillus Endospores. Appl. Environ. Microbiol. 2019, 85, e01596-19. [Google Scholar] [CrossRef]
- De Filippis, F.; Parente, E.; Ercolini, D. Recent Past, Present, and Future of the Food Microbiome. Annu. Rev. Food Sci. Technol. 2018, 9, 589–608. [Google Scholar] [CrossRef]
- Erem, F.; Inan, M.; Budak, B.K.; Certel, M. Partial purification and characterization of an extracellular metallopeptidase produced by Bacillus amyloliquefaciens FE-K1. Trak. Univ. J. Nat. Sci. 2020, 21, 47–61. [Google Scholar] [CrossRef]
- Valerio, F.; Di Biase, M.; Huchet, V.; Desriac, N.; Lonigro, S.; Lavermicocca, P.; Sohier, D.; Postollec, F. Comparison of three Bacillus amyloliquefaciens strains growth behaviour and evaluation of the spoilage risk during bread shelf-life. Food Microbiol. 2015, 45, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Vaiciulyte-Funk, L.; Žvirdauskienė, R.; Šalomskienė, J.; Sarkinas, A. The effect of wheat bread contamination by the Bacillus genus bacteria on the quality and safety of bread. Zemdirb.-Agric. 2015, 102, 351–358. [Google Scholar] [CrossRef]
- Pattison, T.; Lindsay, D.; von Holy, A. In vitro growth response of bread-spoilage Bacillus strains to selected natural antimicrobials. J. Basic Microbiol. 2003, 43, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Fraberger, V.; Unger, C.; Kummer, C.; Domig, K.J. Insights into microbial diversity of traditional Austrian sourdough. LWT 2020, 127, 109358. [Google Scholar] [CrossRef]
- Viedma, P.M.; Abriouel, H.; Ben Omar, N.; López, R.L.; Gálvez, A. Inhibition of spoilage and toxigenic Bacillus species in dough from wheat flour by the cyclic peptide enterocin AS-48. Food Control 2010, 22, 756–761. [Google Scholar] [CrossRef]
- Digaitiene, A.; Hansen, Å.S.; Juodeikiene, G.; Eidukonyte, D.; Josephsen, J. Lactic acid bacteria isolated from rye sourdoughs produce bacteriocin-like inhibitory substances active against Bacillus subtilis and fungi. J. Appl. Microbiol. 2012, 112, 732–742. [Google Scholar] [CrossRef]
- Fraberger, V.; Ammer, C.; Domig, K.J. Functional Properties and Sustainability Improvement of Sourdough Bread by Lactic Acid Bacteria. Microorganisms 2020, 8, 1895. [Google Scholar] [CrossRef]
- Mantzourani, I.; Plessas, S.; Odatzidou, M.; Alexopoulos, A.; Galanis, A.; Bezirtzoglou, E.; Bekatorou, A. Effect of a novel Lactobacillus paracasei starter on sourdough bread quality. Food Chem. 2018, 271, 259–265. [Google Scholar] [CrossRef]
- Plessas, S.; Mantzourani, I.; Bekatorou, A. Evaluation of Pediococcus pentosaceus SP2 as Starter Culture on Sourdough Bread Making. Foods 2020, 9, 77. [Google Scholar] [CrossRef]
- Ghendov-Mosanu, A.; Cristea, E.; Patras, A.; Sturza, R.; Padureanu, S.; Deseatnicova, O.; Turculet, N.; Boestean, O.; Niculaua, M. Potential Application of Hippophae Rhamnoides in Wheat Bread Production. Molecules 2020, 25, 1272. [Google Scholar] [CrossRef]
- Mantzourani, I.; Terpou, A.; Alexopoulos, A.; Bezirtzoglou, E.; Plessas, S. Assessment of Ready-to-Use Freeze-dried Immobilized Biocatalysts as Innovative Starter Cultures in Sourdough Bread Making. Foods 2019, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Valerio, F.; De Bellis, P.; Lonigro, S.L.; Visconti, A.; Lavermicocca, P. Use of Lactobacillus plantarum fermentation products in bread-making to prevent Bacillus subtilis ropy spoilage. Int. J. Food Microbiol. 2008, 122, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Menteş, Ö.; Ercan, R.; Akçelik, M. Inhibitor activities of two Lactobacillus strains, isolated from sourdough, against rope-forming Bacillus strains. Food Control 2007, 18, 359–363. [Google Scholar] [CrossRef]
- Katina, K.; Sauri, M.; Alakomi, H.-L.; Mattila-Sandholm, T. Potential of Lactic Acid Bacteria to Inhibit Rope Spoilage in Wheat Sourdough Bread. LWT Food Sci. Technol. 2002, 35, 38–45. [Google Scholar] [CrossRef]
- Eliseeva, L.G.; Kokorina, D.S.; Zhirkova, E.V.; Smirova, S.A.; Nevskaya, E.V. The Quality and Microbiological Stability of Quinoa-enriched Wheat Bread. IOP Conf. Series: Earth Environ. Sci. 2021, 670, 012020. [Google Scholar] [CrossRef]
- Bücher, C.; Burtscher, J.; Domig, K.J. Propionic acid bacteria in the food industry: An update on essential traits and detection methods. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4299–4323. [Google Scholar] [CrossRef]
- Sudha, M.; Viswanath, P.; Siddappa, V.; Rajarathnam, S.; Shashirekha, M. Control of rope spore forming bacteria using carambola (Averrhoa carambola) fruit pomace powder in wheat bread preparation. Qual. Assur. Saf. Crops Foods 2016, 8, 555–564. [Google Scholar] [CrossRef]
- Dietrich, R.; Jessberger, N.; Ehling-Schulz, M.; Märtlbauer, E.; Granum, P.E. The Food Poisoning Toxins of Bacillus cereus. Toxins 2021, 13, 98. [Google Scholar] [CrossRef]
- European Food Safety Authority. European Centre for Disease Prevention and Control The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Jessberger, N.; Dietrich, R.; Granum, P.E.; Märtlbauer, E. The Bacillus cereus Food Infection as Multifactorial Process. Toxins 2020, 12, 701. [Google Scholar] [CrossRef]
- Rahnama, H.; Azari, R.; Yousefi, M.H.; Berizi, E.; Mazloomi, S.M.; Hosseinzadeh, S.; Derakhshan, Z.; Ferrante, M.; Conti, G.O. A systematic review and meta-analysis of the prevalence of Bacillus cereus in foods. Food Control 2022, 143, 109250. [Google Scholar] [CrossRef]
- De Bellis, P.; Minervini, F.; Di Biase, M.; Valerio, F.; Lavermicocca, P.; Sisto, A. Toxigenic potential and heat survival of spore-forming bacteria isolated from bread and ingredients. Int. J. Food Microbiol. 2015, 197, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Fangio, M.F.; Roura, S.I.; Fritz, R. Isolation and Identification of Bacillus spp. and Related Genera from Different Starchy Foods. J. Food Sci. 2010, 75, M218–M221. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, F.; Arslan, S. Molecular Characterization and Toxin Profiles of Bacillus spp. Isolated from Retail Fish and Ground Beef. J. Food Sci. 2019, 84, 548–556. [Google Scholar] [CrossRef]
- Possas, A.; Valero, A.; Pérez-Rodríguez, F. New software solutions for microbiological food safety assessment and management. Curr. Opin. Food Sci. 2022, 44, 100814. [Google Scholar] [CrossRef]
- Stavropoulou, E.; Bezirtzoglou, E. Predictive Modeling of Microbial Behavior in Food. Foods 2019, 8, 654. [Google Scholar] [CrossRef]
- Desvignes, V.; Buschhardt, T.; Guillier, L.; Sanaa, M. Quantitative microbial risk assessment for Salmonella in eggs. Food Model. J. 2019, 1, e39643. [Google Scholar] [CrossRef]
- Pahlavan, A.; Kamani, M.H.; Elhamirad, A.H.; Sheikholeslami, Z.; Armin, M.; Amani, H. Rapid quality assessment of bread using developed multivariate models: A simple predictive modeling approach. Prog. Agric. Eng. Sci. 2020, 16, 1–10. [Google Scholar] [CrossRef]
- Różyło, R.; Laskowski, J. Predicting bread quality (bread loaf volume and crumb texture). Pol. J. Food Nutr. Sci. 2011, 61, 61–67. [Google Scholar] [CrossRef]
- Gangawane, K.M.; Dwivedi, M. Advanced Computational Techniques for Heat and Mass Transfer in Food Processing, 1st ed.; CRC Press: Boca Raton, FL, USA, 2022; ISBN 9781000521450. [Google Scholar] [CrossRef]
- Qian, C.; Martin, N.; Wiedmann, M.; Trmčić, A. Development of a risk assessment model to predict the occurrence of late blowing defect in Gouda cheese and evaluate potential intervention strategies. J. Dairy Sci. 2022, 105, 2880–2894. [Google Scholar] [CrossRef]



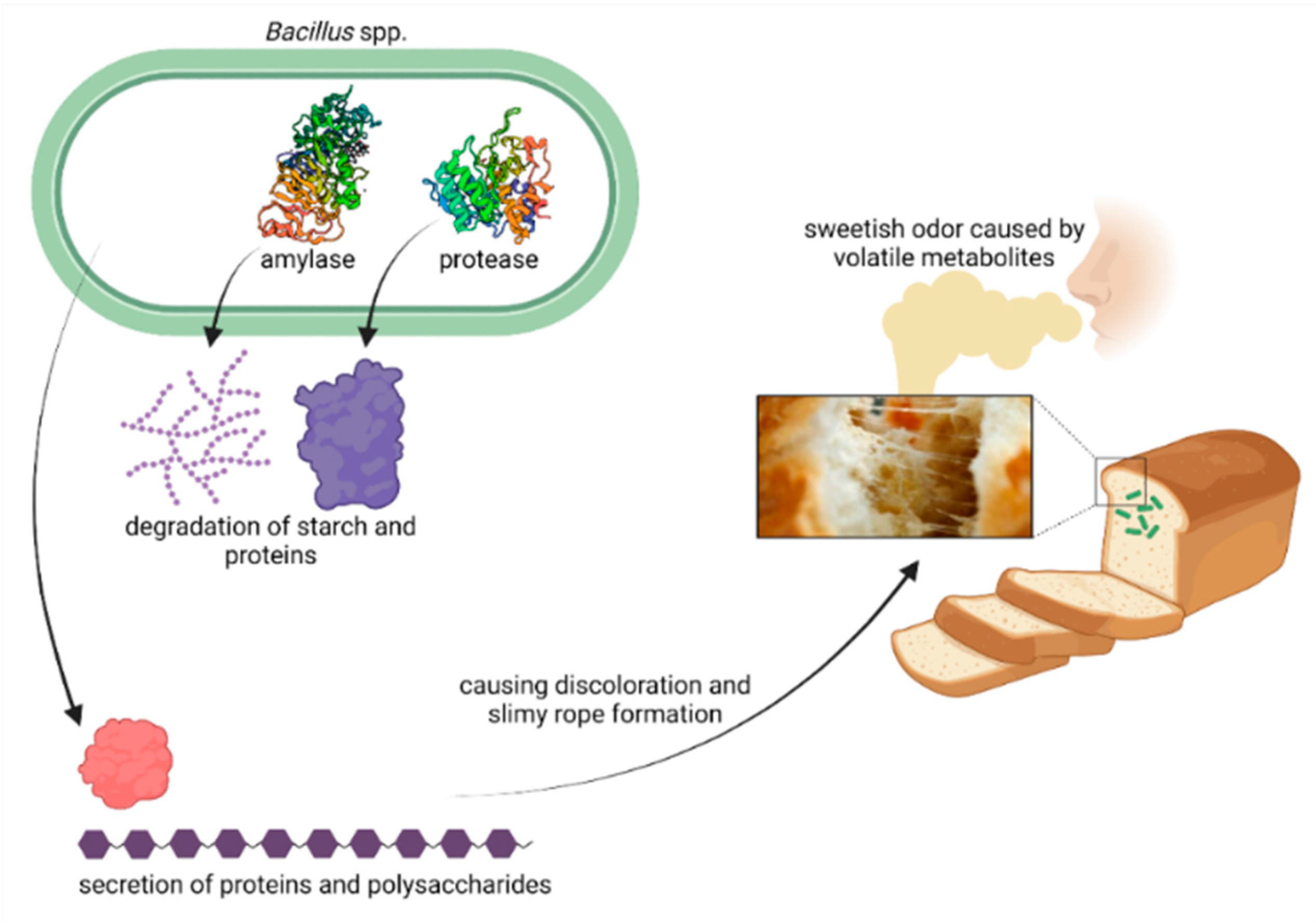{kind=link}
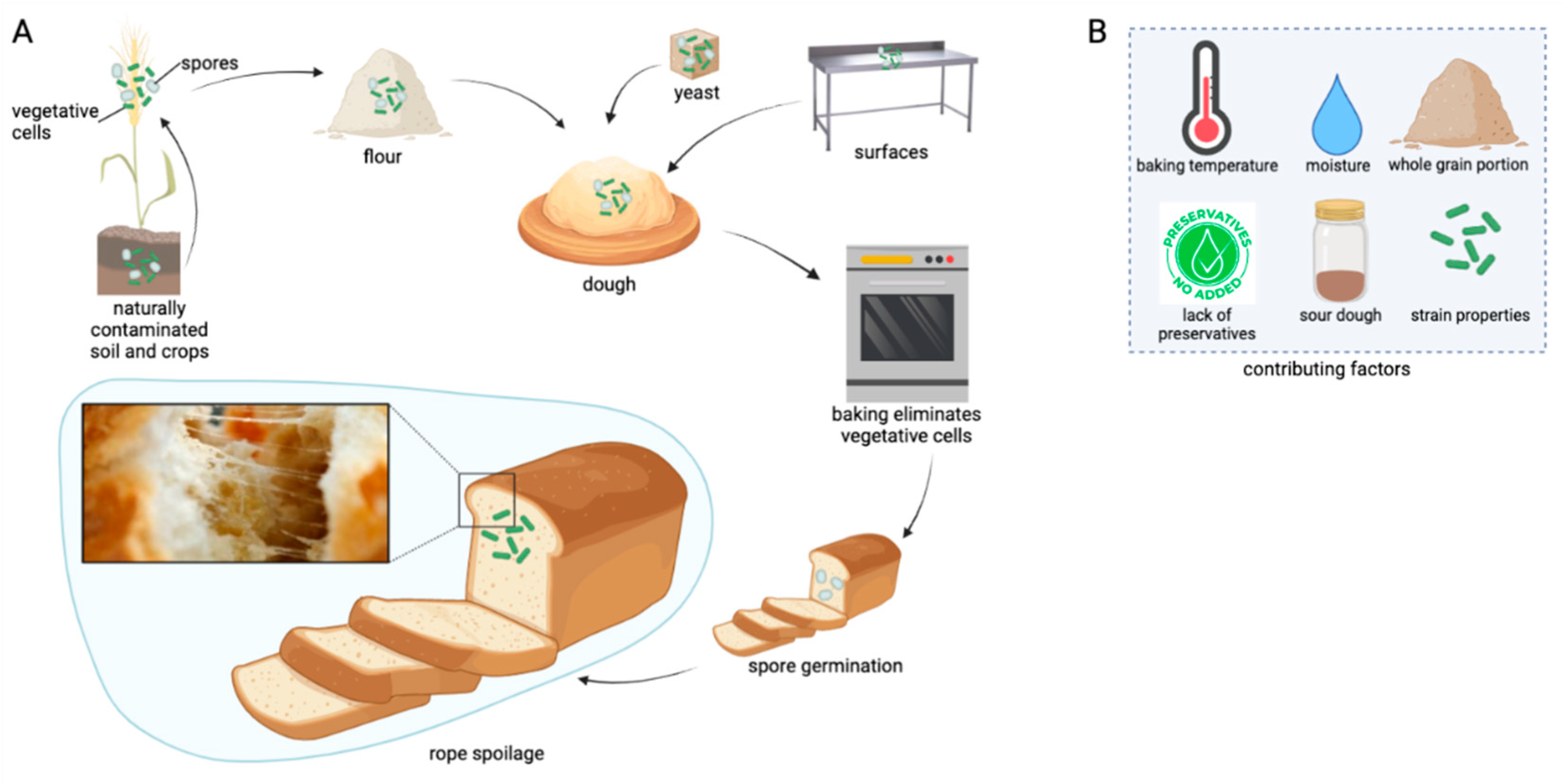{kind=link}
| Growth | Metabolism | Survival | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taxonomy | Optimum Growth Temperature [°C] | Minimum Growth Temperature [°C] | Maximum Growth Temperature [°C] | Growth at pH | NaCl Tolerance | Anaerobic Growth | Urease | Nitrate Reduction | Hydrolysis of Starch | Citrate | Propionate | Egg Yolk Reaction | Spore D100 Value [min] | References |
| B. amyloliquefaciens | 30–40 | 15 | 50 | 5.7 | 5–10% | − | + | d | + | + | n.a. | + | 23–44 | [19,26,50,51,52,57,58,59] |
| B. cereus group | 37 | 5 | 50 | 4.9–9.3 | n.a. | + | d | + | + | d | n.a | + | <10 | [24,26,40,41,41,42,43,44,45,46,47,48,49,50,51,52,60,61] |
| B. licheniformis | 37 | 15 | 50–55 | 5.7 | 7% | + | + | + | + | + | + | − | 56 | [19,24,26,41,50,51,52,59,60,62,63,64] |
| B. pumilus | 30 | 15 | 50–55 | 5.7 | 7% | + | + | + | + | + | + | − | 56 | [19,24,26,41,50,51,52,59,60,65] |
| B. sonorensis | 30 | 15 | 55 | n.a. | <5% | + | + | + | + | + | + | − | n.a. | [41,60] |
| B. subtilisa | 28–30 | 5-20 | 45–55 | 5.5–8.5 | 7–10% | Facultative | − | + | + | + | − | − | 14 | [7,15,24,25,26,41,50,51,52,53,54,60,63,66] |
| C. firmus b | 30–37 | n.a. | n.a. | n.a. | n.a. | n.a. | − | + | + | − | n.a. | n.a. | n.a. | [50,51,52] |
| N. circulans c | 30–37 | n.a. | n.a. | n.a. | n.a. | n.a. | − | n.a. | + | − | n.a. | n.a. | n.a. | [50,51,52] |
| P. polymyxa d | 30 | n.a. | n.a. | n.a. | n.a. | n.a. | − | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | [26,41,60] |
| Pr. megateriume | 30 | 3-15 | 35–45 | n.a. | 7% | − | + | d | + | + | n.a. | − | n.a. | [26,52,65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacher, N.; Burtscher, J.; Johler, S.; Etter, D.; Bender, D.; Fieseler, L.; Domig, K.J. Ropiness in Bread—A Re-Emerging Spoilage Phenomenon. Foods 2022, 11, 3021. https://doi.org/10.3390/foods11193021
Pacher N, Burtscher J, Johler S, Etter D, Bender D, Fieseler L, Domig KJ. Ropiness in Bread—A Re-Emerging Spoilage Phenomenon. Foods. 2022; 11(19):3021. https://doi.org/10.3390/foods11193021
Chicago/Turabian StylePacher, Nicola, Johanna Burtscher, Sophia Johler, Danai Etter, Denisse Bender, Lars Fieseler, and Konrad J. Domig. 2022. "Ropiness in Bread—A Re-Emerging Spoilage Phenomenon" Foods 11, no. 19: 3021. https://doi.org/10.3390/foods11193021