
Detecting the Mechanism of Action of Antimicrobial Peptides by Using Microscopic Detection Techniques


Abstract
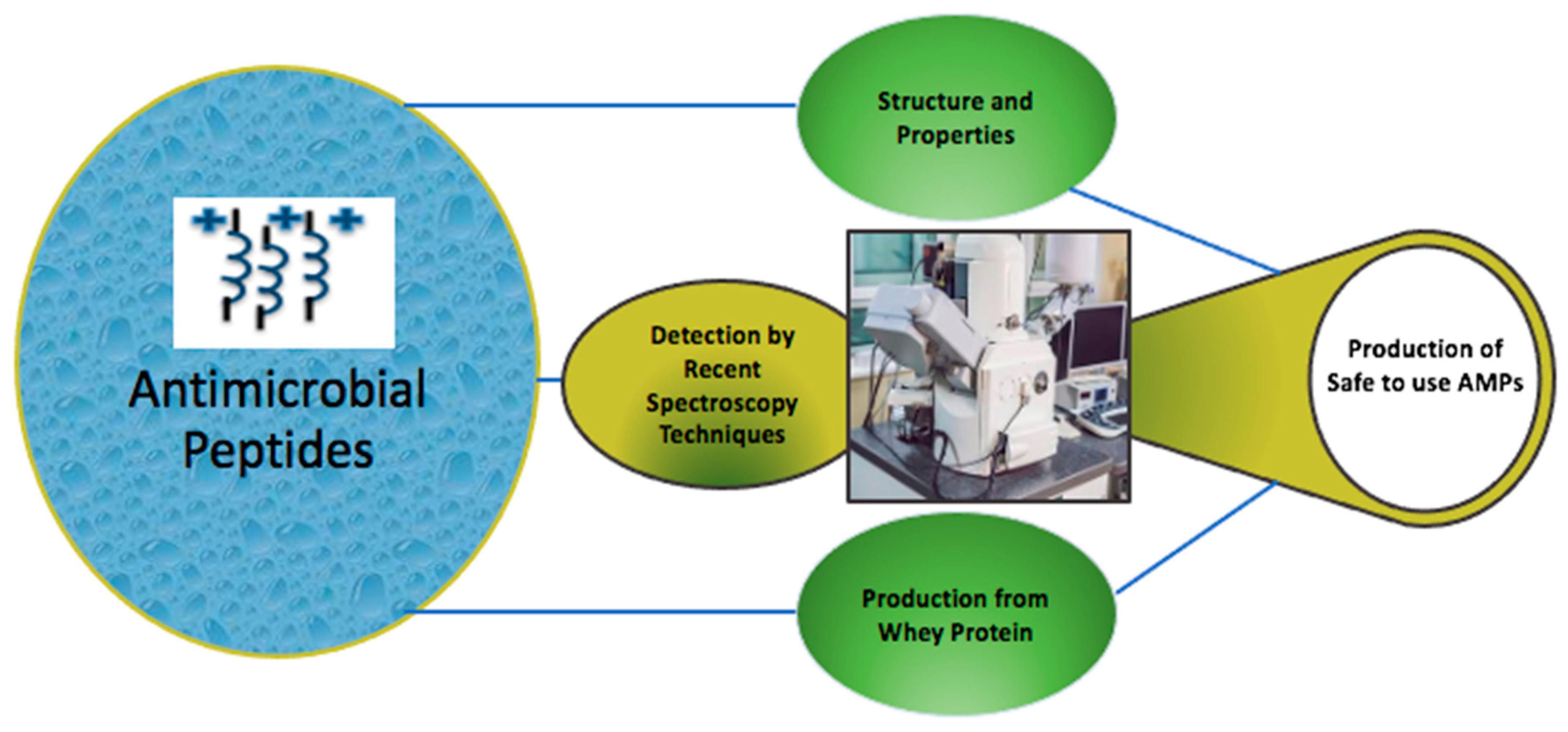
:1. Introduction
2. Structure and Properties of AMPs
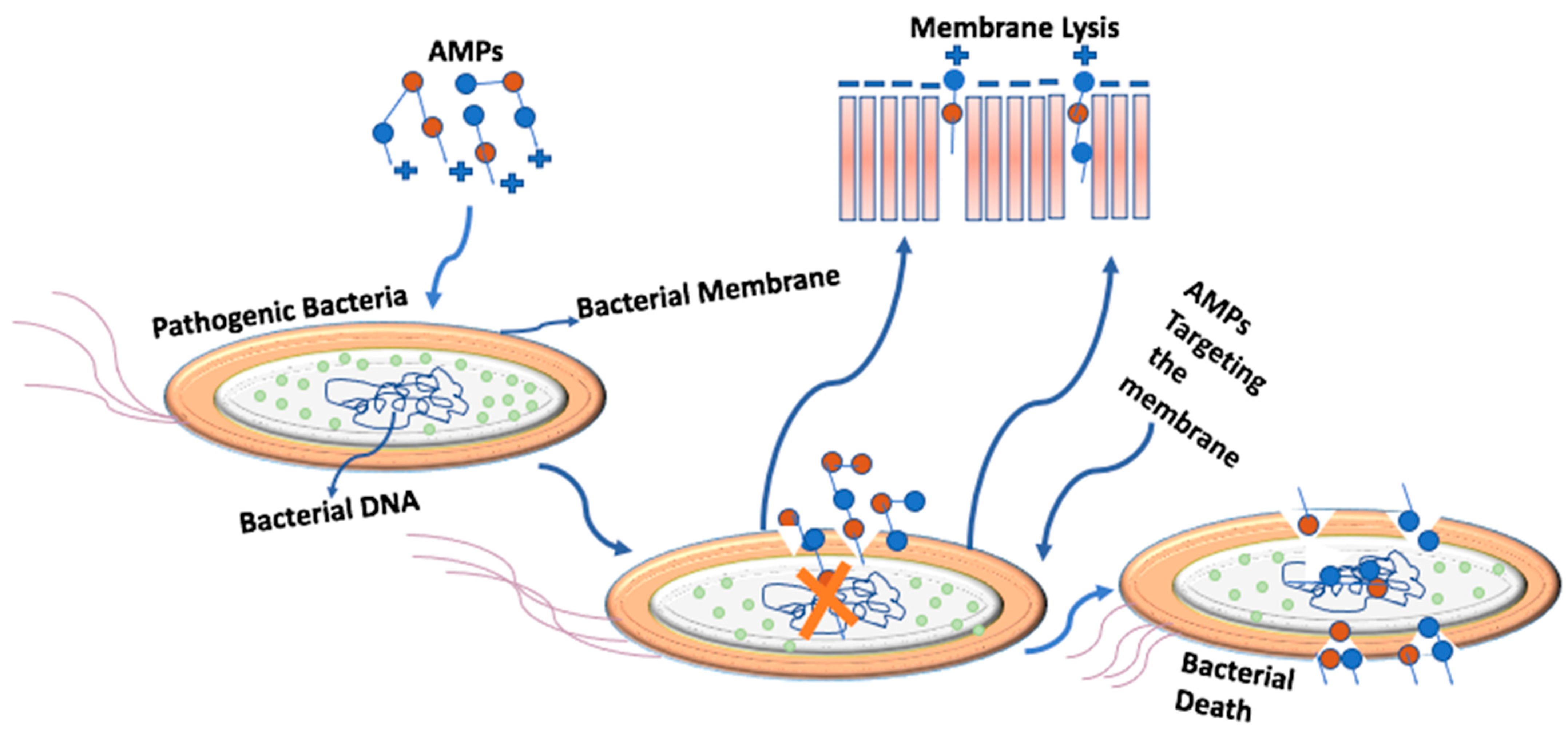
3. The Mechanism of Targeting the Bacterial Cell
4. Recently Tested Peptides

{kind=link}
{kind=link}
{kind=link}
| Name | Source | Secondary Structure | Pathogenic Bacteria | Toxicity | Efficiency | Reference |
|---|---|---|---|---|---|---|
| Monomeric peptide MG2 (Magainin 2) | Skin of African clawed frog | Alpha-helical | Escherichia coli | Low toxicity | Targets the membrane and forms toroidal pores | [28] |
| Peptaibol peptide-Alamethicin | fungus | Alpha-helical | ___ | Toxic | Targets bi-layer lipid membranes by forming pores | [32] |
| Cationic AMP PG-1 (Porcine protegrin-1) | porcine neutrophils | Beta-hairpin | Broad spectrum pathogens | Cytotoxic | Targets the membrane by forming pores | [26] |
| Aurein 1.2 (GLFDIIKKIAESF-NH2) | Skin of Australian bell frogs | Alpha-helical | ___ | ___ | Targets the bacterial membrane | [33] |
| Cysteine-rich HBD2 (Human beta-defensin 2) | Human skin | Beta-strand and Alpha-helical | Pseudomonas aeruginosa | Non-toxic | Targets the bacterial membrane and inhibits biofilm formation | [29] |
| Cathelicidin LL-37 | Human skin, epithelial cells | Cationic amphipathic Alpha-helical | Broad spectrum pathogens | Non-Toxic | Targets the Cationic and anionic membranes of pathogens | [30] |
| Nisin | Lactococcus lactis | Looped | Streptococcus mutans | Non-toxic | Targets the pathogen’s lipid bi-layer membrane | [34] |
| 45 analogs of AMP indolicidin | Neutrophil blood cells of cows | Alpha-helical | Gram+ and gram− bacteria | Toxic at high concentration | ___ | [35] |
| AMPs of the temporin family (Temporin-She) | The skin of the Sahara Frog | Non-amphipathic α-helical peptide α-helical peptide | Leishmania infantum, and Staphylococcus aureus | Toxic | Targets Gram+ and gram− bacteria by damaging the lipid chain of the membrane | [36] |
| Pro10-1D | Designed in the lab | Alpha-helical | Escherichia coli, and Acinetobacter baumannii | Non-toxic | Targets the bacterial membrane and damages the cell efficiently | [11] |
| AP-64 | Human | Alpha-helical | Escherichia coli DH5α, Escherichia coli O157:H7, Vibrio cholerae, and Pseudomonas aeruginosa | Cytotoxic | Targets the membrane | [37] |
| LL-14 | lysosomes and polymorphonuclear leukocytes | Helical | Escherichia coli, Salmonella typhi, klebsiella pneumoniae, Staphylococcus aureus | LL-14 Non-toxic | Membrane depolarization and cell death | [38] |
| Protein-glutamine gamma-glutamyltransferase 2 | Hemoglobin of blood clam | Alpha helical | Escherichia coli | Less Toxic | Targets membrane by making nano pores through which cellular material leaks out | [39] |
| Crustin (rCrus1) | shrimp | Alpha helical | Gram+ bacteria | ___ | Damages the cellular machinery in target cells | [40] |
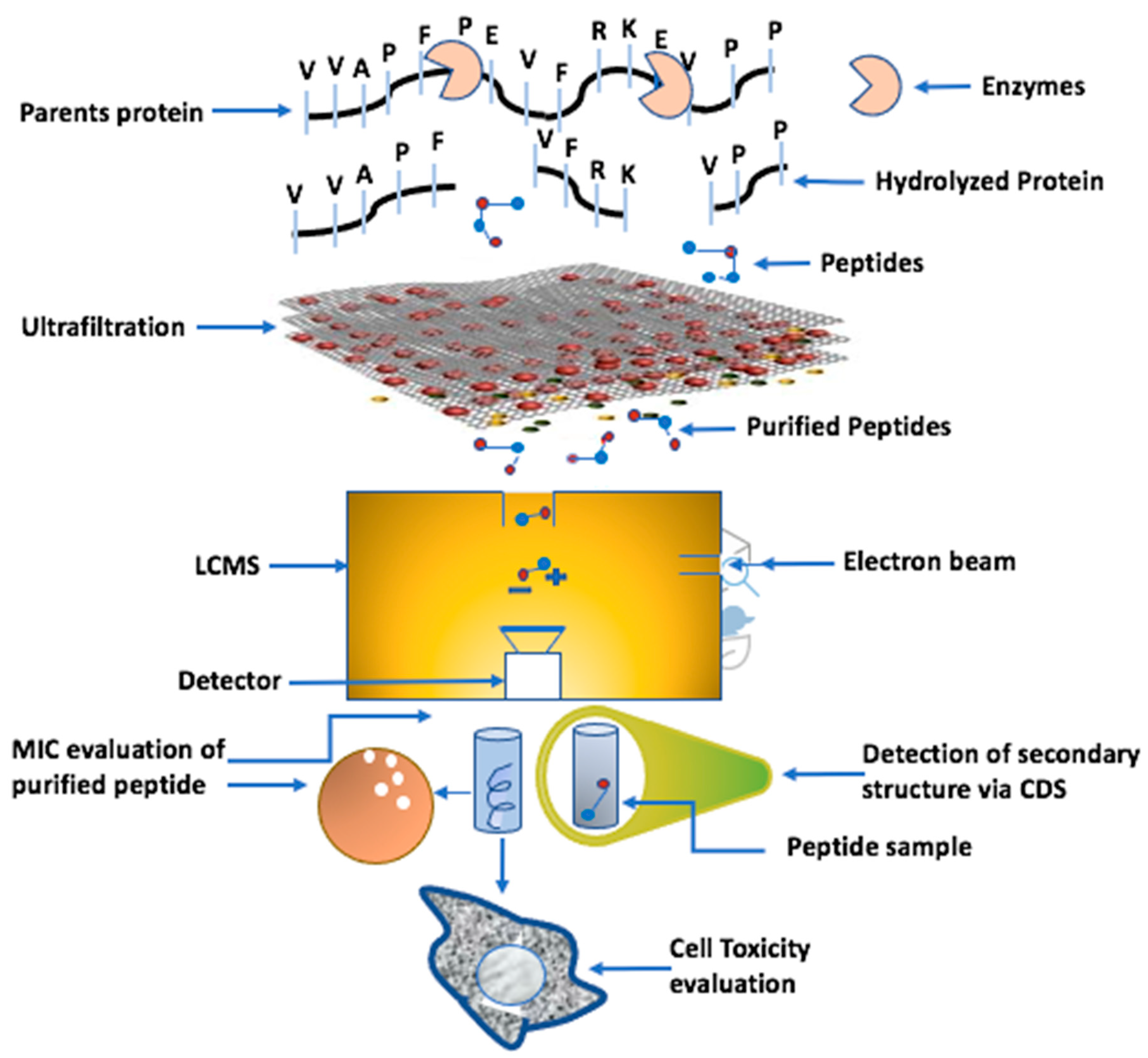
5. Recent Developments in the Use of Microscopy Techniques to Highlight AMP Targeting
| Pathogens | AMPs | Microscopic Technique | Observations | Reference |
|---|---|---|---|---|
| Pseudomonas Aeruginosa | DP7, (synthetic) | Gel retardation assay | DP7 targets the membrane protein and damages bacterial membrane | [41] |
| Staphylococcus aureus | Porcine beta defensin 2, (synthetic) | Transmission electron microscopy | AMP targets the cell membrane and then enters the cytoplasm | [42] |
| Staphylococcus aureus | Temporin-She, (extracted from frog, mildly cationic with charge of +2) | Scanning electron microscopy | AMP targets the anionic cell membrane | [36] |
| Escherichia coli, and Acinetobacter baumannii | Pro10-1D (synthetic with charge of +4) | Scanning electron microscopy | Targets the bacterial membrane | [11] |
| Escherichia coli | P6.2 (synthetic) | Atomic force microscopy | Targets the pathogen at the membrane level | [43] |
| Methicillin-resistant Staphylococcus aureus | PVP (synthetic) | Fluorescence microscopy | Increasse membrane permeability and causes cell lysis | [44] |
| Escherichia coli | Temporin L (extracted from frog skin) | Transmission electron microscopy | After interacting with the membrane protein, this AMP forms nanopores | [23] |
| Staphylococcus aureus and Staphylococcus epidermidis. | Cecropin, Magainin 2, and melittin | Field Emission Scanning Electron Microscopy | These AMPs target the membrane and form pores on it | [45] |
| Pseudomonas aeruginosa | undecapeptides (AMP21-24) | Field emission scanning electron microscopy | Targets the membrane | [38] |
| Pseudomonas fluorescens | Temporin-L (extracted from frog) | Confocal Laser Scanning Microscopy | Targets the bacterial biofilm | [46] |
| Escherichia coli | cecropin A (extracted from honeycomb moth) | Scanning electron microscopy | Disrupts bacterial membrane and targets the biofilm | [47] |
| Mycobacterium smegmatis, and Mycobacterium tuberculosis | HHC-8, and MM-10 (Synthetic) | Scanning electron microscopy | Targets the membrane and makes it permeable to penetrate | [48] |
| Escherichia coli | Pa-Methionine aminopeptidase 2 and Pa-Methionine aminopeptidase 2 1.9 (Synthetic) | Atomic force microscopy | Cationic AMPs target the anionic membrane and cause cell death | [49] |
| Gram+ bacteria | rCrus1 (Extracted from shrimp) | Scanning electron microscope and Transmission electron microscope | This AMP causes membrane leakage and structure damage In the pathogen | [40] |
| Escherichia coli | Protamine (extracted from salmon sperm) and OH-CATH-30 (Synthesized) | Electron Microscopy | AMP adheres to the target membrane | [50] |
| Escherichia coli DH5α, Escherichia coli O157:H7, Vibrio cholerae, and Pseudomonas aeruginosa | AP-64 (extracted from human lacking cysteine) | scanning electron microscopy | Targets the membrane | [37] |
| Salmonella typhi TY2 | LL-14, VV-14 and ββ-14 (synthetic) | Field emission scanning electron microscopy | Targets the membrane, causes depolarization and eventually cell lysis | [51] |
| Escherichia coli | Protein-glutamine gamma-glutamyltransferase 2 (extracted from blood clam hemoglobin) | Transmission electron microscopy | Increases membrane permeability | [39] |
| Escherichia coli | Disulfide-rich β-defensin AvBD103b (extracted from avian defensin) | Real time Fluorescence microscopy | Targets the outer and cytoplasm membrane and disrupts homeostasis | [52] |
| Enterococcus faecalis, Klebsiella pneumoniae, and Pseudomonas aeruginosa | Synoeca-MP (extracted from the venom of Synoeca surinama) | Atomic force microscopy | Targets the membrane | [53] |
| Eschericia coli and Staphylococcus aureus | Arginine-rich peptide Bac8c2,5Leu (synthetic) | Scanning electron microscopy | Effective in targeting pathogens | [54] |
| Enterococcus hirae | SAAP-148 (synthetic) | Fluorescence Microscopy | Disrupts the Anionic membrane and cell shrinkage | [55] |
| Porphyromonas gingivalis | DP7 (synthetic) | Transmission electron microscopy | Targets the bacterial membrane and inhibits biofilm formation | [56] |
| Nosocomial bacterial pathogens | Pardaxin, MSI-78, dermaseptin-PC, and Cecropin B (Synthetic) | Helium ion microscopy | Targets the membrane | [57] |
| Streptococcus agalactiae | NZX and P2 (extracted from fungal defensin) | Scanning electron microscopy | Targets the cell wall and disrupts the membrane | [58] |
| Bacillus circulans | BaCf3 (extracted from Bacillus amyloliquefaciens) | Scanning electron microscopy | Targets the membrane by pore formation | [59] |
| Staphylococcus aureus, Streptococcus agalactiae,Vibrio harveyi, Vibrio alginolyticus, Escherichia coli, and Edwardsiella tarda | TroNKL-27 (extracted from golden pompano) | Scanning electron microscopy | Targets the pathogen and degrades the DNA after penetrating | [60] |
| Staphylococcus aureus | Cruzioseptins (extracted from splendid treefrog) | Fluorescence Microscopy | Targets the bacterial membrane | [61] |
| Methicillin-resistant Staphylococcus aureus, Escherichia coli | Proline-rich antimicrobial peptides | Scanning electron microscopy | Efficient in targeting the membrane | [62] |
| Staphylococcus aureus | LCMHC (extracted from Larimichthys crocea) | Transmission electron microscopy | Targets the cell membrane | [63] |
| Cutibacterium acnes | WSKK11 and WSRR11 | Scanning electron microscopy, transmission electron microscopy | Efficient in targeting the pathogens | [64] |
6. The Production of AMPs from Whey Protein
7. Conclusions and Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Silveira, R.F.; Roque-Borda, C.A.; Vicente, E.F. Antimicrobial peptides as a feed additive alternative to animal production, food safety and public health implications: An overview. Anim. Nutr. 2021, 7, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-Y.; Hu, Q.; Xu, F.; Ding, S.-Y.; Zhu, K. Characterization of Bacillus cereus in dairy products in China. Toxins 2020, 12, 454. [Google Scholar] [CrossRef]
- Huang, H.W.; Charron, N.E. Understanding membrane-active antimicrobial peptides. Q. Rev. Biophys. 2017, 50, e10. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.R.P.; Guimarães, M.S.; Rabelo, J.; Belén, L.H.; Perecin, C.J.; Farías, J.G.; Santos, J.H.P.M.; Rangel-Yagui, C.O. Recent advances in the design of antimicrobial peptide conjugates. J. Mater. Chem. B 2022, 10, 3587–3600. [Google Scholar] [CrossRef] [PubMed]
- Parchebafi, A.; Tamanaee, F.; Ehteram, H.; Ahmad, E.; Nikzad, H.; Haddad Kashani, H.J. The dual interaction of antimicrobial peptides on bacteria and cancer cells; mechanism of action and therapeutic strategies of nanostructures. Microb. Cell Factories 2022, 21, 118. [Google Scholar] [CrossRef]
- Mishra, A.K.; Choi, J.; Moon, E.; Baek, K.-H. Tryptophan-rich and proline-rich antimicrobial peptides. Molecules 2018, 23, 815. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Feng, L.; Wang, Y.; Yang, J.; Sun, Y.-F.; Li, Y.-W.; Ye, Z.-H.; Lin, H.-B.; Yang, K. Overview of the preparation method, structure and function, and application of natural peptides and polypeptides. Biomed. Pharmacother. 2022, 153, 113493. [Google Scholar] [CrossRef] [PubMed]
- Smithers, G.W. Whey-ing up the options–Yesterday, today and tomorrow. Int. Dairy J. 2015, 48, 2–14. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial peptides: Classification, design, application and research progress in multiple fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Krishnan, M.; Choi, J.; Jang, A.; Kim, Y. A novel peptide antibiotic, Pro10-1D, designed from insect defensin shows antibacterial and anti-inflammatory activities in sepsis models. Int. J. Mol. Sci. 2020, 21, 6216. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, G.; Cui, X.; Chen, J.; Yu, Q.; Zong, C.; Zhao, Y.; Xu, M.; Zhou, S.; Xu, H. Mechanistic investigation of a self-assembling peptide against Escherichia coli. Langmuir 2020, 36, 9800–9809. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Javor, S.; Gan, B.H.; Köhler, T.; Reymond, J.-L. The antibacterial activity of peptide dendrimers and polymyxin B increases sharply above pH 7.4. Chem. Commun. 2021, 57, 5654–5657. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, D.; Jena, R.; Choudhury, P.K.; Pattnaik, R.; Mohapatra, S.; Saini, M.R. Milk derived antimicrobial bioactive peptides: A review. Int. J. Food Prop. 2016, 19, 837–846. [Google Scholar] [CrossRef]
- Spohn, R.; Daruka, L.; Lázár, V.; Martins, A.; Vidovics, F.; Grézal, G.; Méhi, O.; Kintses, B.; Számel, M.; Jangir, P.; et al. Integrated evolutionary analysis reveals antimicrobial peptides with limited resistance. Nat. Commun. 2019, 10, 4538. [Google Scholar] [CrossRef]
- Battersby, A.J.; Khara, J.; Wright, V.J.; Levy, O.; Kampmann, B. Antimicrobial proteins and peptides in early life: Ontogeny and translational opportunities. Front. Immunol. 2016, 7, 309. [Google Scholar] [CrossRef]
- Bechinger, B.; Gorr, S.-U. Antimicrobial peptides: Mechanisms of action and resistance. J. Dent. Res. 2017, 96, 254–260. [Google Scholar] [CrossRef]
- Berthelot, K.; Peruch, F.; Lecomte, S. Highlights on Hevea brasiliensis (pro) hevein proteins. Biochimie 2016, 127, 258–270. [Google Scholar] [CrossRef]
- Kim, C.-H.; Go, H.-J.; Oh, H.Y.; Park, J.B.; Lee, T.K.; Seo, J.-K.; Elphick, M.R.; Park, N.G. Identification of a novel antimicrobial peptide from the sea star Patiria pectinifera. Dev. Comp. Immunol. 2018, 86, 203–213. [Google Scholar] [CrossRef]
- Koehbach, J.; Craik, D.J. The vast structural diversity of antimicrobial peptides. Trends Pharmacol. Sci. 2019, 40, 517–528. [Google Scholar] [CrossRef]
- Bogdanova, L.; Valiullina, Y.; Faizullin, D.; Kurbanov, R.K.; Ermakova, E. Spectroscopic, zeta potential and molecular dynamics studies of the interaction of antimicrobial peptides with model bacterial membrane. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 242, 118785. [Google Scholar] [CrossRef] [PubMed]
- Barreto-Santamaría, A.; Curtidor, H.; Arévalo-Pinzón, G.; Herrera, C.; Suárez, D.; Pérez, W.H.; Patarroyo, M.E. A new synthetic peptide having two target of antibacterial action in E. coli ML35. Front. Microbiol. 2016, 7, 2006. [Google Scholar] [CrossRef] [PubMed]
- Di Somma, A.; Avitabile, C.; Cirillo, A.; Moretta, A.; Merlino, A.; Paduano, L.; Duilio, A.; Romanelli, A. The antimicrobial peptide Temporin L impairs E. coli cell division by interacting with FtsZ and the divisome complex. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2020, 1864, 129606. [Google Scholar] [CrossRef] [PubMed]
- Graf, M.; Wilson, D.N. Intracellular antimicrobial peptides targeting the protein synthesis machinery. Antimicrob. Pept. 2019, 1117, 73–89. [Google Scholar]
- Pradhan, B.; Guha, D.; Murmu, K.C.; Sur, A.; Ray, P.; Das, D.; Aich, P. Comparative efficacy analysis of anti-microbial peptides, LL-37 and indolicidin upon conjugation with CNT, in human monocytes. J. Nanobiotechnol. 2017, 15, 44. [Google Scholar] [CrossRef]
- Soundrarajan, N.; Park, S.; Le Van Chanh, Q.; Cho, H.-S.; Raghunathan, G.; Ahn, B.; Song, H.; Kim, J.-H.; Park, C. Protegrin-1 cytotoxicity towards mammalian cells positively correlates with the magnitude of conformational changes of the unfolded form upon cell interaction. Sci. Rep. 2019, 9, 11569. [Google Scholar] [CrossRef]
- Taniguchi, M.; Ochiai, A.; Kondo, H.; Fukuda, S.; Ishiyama, Y.; Saitoh, E.; Kato, T.; Tanaka, T. Pyrrhocoricin, a proline-rich antimicrobial peptide derived from insect, inhibits the translation process in the cell-free Escherichia coli protein synthesis system. J. Biosci. Bioeng. 2016, 121, 591–598. [Google Scholar] [CrossRef]
- Lorenzon, E.N.; Nobre, T.M.; Caseli, L.; Cilli, E.M.; da Hora, G.C.; Soares, T.A.; Oliveira, O.N., Jr. The “pre-assembled state” of magainin 2 lysine-linked dimer determines its enhanced antimicrobial activity. Colloids Surf. B Biointerfaces 2018, 167, 432–440. [Google Scholar] [CrossRef]
- Parducho, K.R.; Beadell, B.; Ybarra, T.K.; Bush, M.; Escalera, E.; Trejos, A.T.; Chieng, A.; Mendez, M.; Anderson, C.; Park, H.; et al. The antimicrobial peptide human beta-defensin 2 inhibits biofilm production of Pseudomonas aeruginosa without compromising metabolic activity. Front. Immunol. 2020, 11, 805. [Google Scholar] [CrossRef]
- Moreno-Angarita, A.; Aragón, C.C.; Tobón, G.J. Cathelicidin LL-37: A new important molecule in the pathophysiology of systemic lupus erythematosus. J. Transl. Autoimmun. 2020, 3, 100029. [Google Scholar] [CrossRef]
- Anumudu, C.; Hart, A.; Miri, T.; Onyeaka, H. Recent advances in the application of the antimicrobial peptide nisin in the inactivation of spore-forming bacteria in foods. Molecules 2021, 26, 5552. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, F.; Leitch, J.J.; Su, Z.; Szymanski, G.; Lipkowski, J. Direct visualization of alamethicin ion pores formed in a floating phospholipid membrane supported on a gold electrode surface. Electrochim. Acta 2018, 267, 195–205. [Google Scholar] [CrossRef]
- Migoń, D.; Jaśkiewicz, M.; Neubauer, D.; Bauer, M.; Sikorska, E.; Kamysz, E.; Kamysz, W. Alanine scanning studies of the antimicrobial peptide aurein 1.2. Probiotics Antimicrob. Proteins 2019, 11, 1042–1054. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Qu, Y.; Liu, J.; Mai, S.; Gu, L. A universal adhesive incorporating antimicrobial peptide nisin: Effects on Streptococcus mutans and saliva-derived multispecies biofilms. Odontology 2020, 108, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, M.; Kolodkin, N.; Kolobov, A.; Afonin, V.; Afonina, I.; Stefanenko, L.; Shpen’, V.; Shamova, O. Indolicidin analogs with broad-spectrum antimicrobial activity and low hemolytic activity. Peptides 2020, 132, 170356. [Google Scholar] [CrossRef] [PubMed]
- André, S.; Raja, Z.; Humblot, V.; Piesse, C.; Foulon, T.; Sereno, D.; Oury, B.; Ladram, A. Functional characterization of temporin-SHe, a new broad-spectrum antibacterial and Leishmanicidal temporin-SH paralog from the sahara frog (Pelophylax saharicus). Int. J. Mol. Sci. 2020, 21, 6713. [Google Scholar] [CrossRef]
- Zhong, K.; Wang, Y.; Wang, Z.; Zhang, Z.; Zhao, S.; Li, H.; Huang, J.; Guo, W.; Zheng, X.; Guo, G.; et al. AP-64, Encoded by C5orf46, Exhibits Antimicrobial Activity against Gram-Negative Bacteria. Biomolecules 2021, 11, 485. [Google Scholar] [CrossRef]
- Pandit, G.; Biswas, K.; Ghosh, S.; Debnath, S.; Bidkar, A.P.; Satpati, P.; Bhunia, A.; Chatterjee, S. Rationally designed antimicrobial peptides: Insight into the mechanism of eleven residue peptides against microbial infections. Biochim. Biophys. Acta (BBA)—Biomembr. 2020, 1862, 183177. [Google Scholar] [CrossRef]
- Yang, S.; Yuan, Z.; Aweya, J.J.; Huang, S.; Deng, S.; Shi, L.; Zheng, M.; Zhang, Y.; Liu, G. Low-intensity ultrasound enhances the antimicrobial activity of neutral peptide TGH2 against Escherichia coli. Ultrason. Sonochemistry 2021, 77, 105676. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Sun, Y.; Sun, L. A Crustin from Hydrothermal Vent Shrimp: Antimicrobial Activity and Mechanism. Mar. Drugs 2021, 19, 176. [Google Scholar] [CrossRef]
- Yin, Q.; Wu, S.; Wu, L.; Wang, Z.; Mu, Y.; Zhang, R.; Dong, C.; Zhou, B.; Zhao, B.; Zheng, J.; et al. A novel in silico antimicrobial peptide DP7 combats MDR Pseudomonas aeruginosa and related biofilm infections. J. Antimicrob. Chemother. 2020, 75, 3248–3259. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, H.; Gao, C.; Chen, R.; Li, C. Antimicrobial Mechanism of pBD2 against Staphylococcus aureus. Molecules 2020, 25, 3513. [Google Scholar] [CrossRef] [PubMed]
- Maturana, P.; Gonçalves, S.; Martinez, M.; Espeche, J.C.; Santos, N.C.; Semorile, L.; Maffia, P.C.; Hollmann, A. Interactions of “de novo” designed peptides with bacterial membranes: Implications in the antimicrobial activity. Biochim. Biophys. Acta (BBA)—Biomembr. 2020, 1862, 183443. [Google Scholar] [CrossRef]
- Memariani, H.; Memariani, M.; Robati, R.M.; Nasiri, S.; Abdollahimajd, F.; Baseri, Z.; Moravvej, H. Anti-Staphylococcal and cytotoxic activities of the short anti-microbial peptide PVP. World J. Microbiol. Biotechnol. 2020, 36, 174. [Google Scholar]
- Boix-Lemonche, G.; Lekka, M.; Skerlavaj, B. A Rapid Fluorescence-Based Microplate Assay to Investigate the Interaction of Membrane Active Antimicrobial Peptides with Whole Gram-Positive Bacteria. Antibiotics 2020, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Di Somma, A.; Recupido, F.; Cirillo, A.; Romano, A.; Romanelli, A.; Caserta, S.; Guido, S.; Duilio, A. Antibiofilm Properties of Temporin-L on Pseudomonas fluorescens in Static and In-Flow Conditions. Int. J. Mol. Sci. 2020, 21, 8526. [Google Scholar] [CrossRef] [PubMed]
- Kalsy, M.; Tonk, M.; Hardt, M.; Dobrindt, U.; Zdybicka-Barabas, A.; Cytrynska, M.; Vilcinskas, A.; Mukherjee, K. The insect antimicrobial peptide cecropin A disrupts uropathogenic Escherichia coli biofilms. NPJ Biofilms Microbiomes 2020, 6, 6. [Google Scholar] [CrossRef]
- Sharma, A.; Gaur, A.; Kumar, V.; Sharma, N.; Patil, S.A.; Verma, R.K.; Singh, A.K. Antimicrobial activity of synthetic antimicrobial peptides loaded in poly-ε-caprolactone nanoparticles against mycobacteria and their functional synergy with rifampicin. Int. J. Pharm. 2021, 608, 121097. [Google Scholar] [CrossRef]
- Felício, M.R.; Silveira, G.G.O.S.; Oshiro, K.G.N.; Meneguetti, B.T.; Franco, O.L.; Santos, N.C.; Gonçalves, S. Polyalanine peptide variations may have different mechanisms of action against multidrug-resistant bacterial pathogens. J. Antimicrob. Chemother. 2021, 76, 1174–1186. [Google Scholar] [CrossRef]
- Wieland, T.; Assmann, J.; Bethe, A.; Fidelak, C.; Gmoser, H.; Janßen, T.; Kotthaus, K.; Lübke-Becker, A.; Wieler, L.; Urban, G. A Real-Time Thermal Sensor System for Quantifying the Inhibitory Effect of Antimicrobial Peptides on Bacterial Adhesion and Biofilm Formation. Sensors 2021, 21, 2771. [Google Scholar] [CrossRef]
- Pandit, G.; Chowdhury, N.; Mohid, S.A.; Bidkar, A.P.; Bhunia, A.; Chatterjee, S. Effect of Secondary Structure and Side Chain Length of Hydrophobic Amino Acid Residues on the Antimicrobial Activity and Toxicity of 14-Residue-Long de novo AMPs. ChemMedChem 2020, 16, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Landon, C.; Zhu, Y.; Mustafi, M.; Madinier, J.-B.; Lelièvre, D.; Aucagne, V.; Delmas, A.F.; Weisshaar, J.C. Real-Time Fluorescence Microscopy on Living E. coli Sheds New Light on the Antibacterial Effects of the King Penguin β-Defensin AvBD103b. Int. J. Mol. Sci. 2022, 23, 2057. [Google Scholar] [CrossRef] [PubMed]
- Junior, A.G.D.S.; Frias, I.A.; Lima-Neto, R.G.; Franco, O.L.; Oliveira, M.D.; Andrade, C.A. Electrochemical detection of gram-negative bacteria through mastoparan-capped magnetic nanoparticle. Enzym. Microb. Technol. 2022, 160, 110088. [Google Scholar] [CrossRef] [PubMed]
- Barzan, G.; Kokalari, I.; Gariglio, G.; Ghibaudi, E.; Devocelle, M.; Monopoli, M.P.; Sacco, A.; Greco, A.; Giovannozzi, A.M.; Rossi, A.M.; et al. Molecular Aspects of the Interaction with Gram-Negative and Gram-Positive Bacteria of Hydrothermal Carbon Nanoparticles Associated with Bac8c2, 5Leu Antimicrobial Peptide. ACS Omega 2022, 7, 16402–16413. [Google Scholar] [CrossRef]
- Piller, P.; Wolinski, H.; Cordfunke, R.A.; Drijfhout, J.W.; Keller, S.; Lohner, K.; Malanovic, N. Membrane Activity of LL-37 Derived Antimicrobial Peptides against Enterococcus hirae: Superiority of SAAP-148 over OP-145. Biomolecules 2022, 12, 523. [Google Scholar] [CrossRef]
- Jiang, S.; Xiao, X.; Zheng, J.; Lai, S.; Yang, L.; Li, J.; Liu, C.; Yang, Y.; Mu, Y. Antibacterial and Antibiofilm Activities of Novel Antimicrobial Peptide DP7 against the Periodontal Pathogen Porphyromonas gingivalis. J. Appl. Microbiol. 2022, 133, 1052–1062. [Google Scholar] [CrossRef]
- Lin, B.; Hung, A.; Li, R.; Barlow, A.; Singleton, W.; Matthyssen, T.; Sani, M.-A.; Hossain, M.A.; Wade, J.D.; O’Brien-Simpson, N.M.; et al. Systematic comparison of activity and mechanism of antimicrobial peptides against nosocomial pathogens. Eur. J. Med. Chem. 2022, 231, 114135. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, N.; Mao, R.; Hao, Y.; Teng, D.; Wang, J. In Vitro Pharmacodynamics and Bactericidal Mechanism of Fungal Defensin-Derived Peptides NZX and P2 against Streptococcus agalactiae. Microorganisms 2022, 10, 881. [Google Scholar] [CrossRef]
- Saidumohamed, B.E.; Johny, T.K.; Raveendran, A.T.; Sheela, U.B.; Sreeranganathan, M.; Sasidharan, R.S.; Bhat, S.G. 3D Structure Elucidation and Appraisal of Mode of Action of a Bacteriocin BaCf3 with Anticancer Potential Produced by Marine Bacillus amyloliquefaciens BTSS3. Re GEN Open 2022, 2, 45–56. [Google Scholar] [CrossRef]
- Zhang, H.; Cao, Z.; Diao, Q.; Zhou, Y.; Ao, J.; Liu, C.; Sun, Y. Antimicrobial activity and mechanisms of a derived antimicrobial peptide TroNKL-27 from golden pompano (Trachinotus ovatus) NK-lysin. Fish Shellfish Immunol. 2022, 126, 357–369. [Google Scholar] [CrossRef]
- Valdivieso-Rivera, F.; Bermúdez-Puga, S.; Proaño-Bolaños, C.; Almeida, J.R. Deciphering the Limitations and Antibacterial Mechanism of Cruzioseptins. Int. J. Pept. Res. Ther. 2022, 28, 73. [Google Scholar] [CrossRef]
- Dewangan, R.P.; Verma, D.P.; Verma, N.K.; Gupta, A.; Pant, G.; Mitra, K.; Habib, S.; Ghosh, J.K. Spermine-Conjugated Short Proline-Rich Lipopeptides as Broad-Spectrum Intracellular Targeting Antibacterial Agents. J. Med. Chem. 2022, 65, 5433–5448. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Aweya, J.J.; Li, J.; Wang, Z.; Huang, S.; Zheng, M.; Shi, L.; Deng, S.; Yang, S. Synergistic antibacterial effects of low-intensity ultrasound and peptide LCMHC against Staphylococcus aureus. Int. J. Food Microbiol. 2022, 373, 109713. [Google Scholar] [CrossRef]
- Theansungnoen, T.; Phosri, S.; Bumrungthai, S.; Daduang, J.; Klaynongsruang, S.; Daduang, S. Novel non-cytotoxic antimicrobial peptides WSKK11 and WSRR11 with potent activity against Cutibacterium acnes. J. Antimicrob. Chemother. 2022, 77, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Sawin, E.A.; De Wolfe, T.J.; Aktas, B.; Stroup, B.M.; Murali, S.G.; Steele, J.L.; Ney, D.M. Glycomacropeptide is a prebiotic that reduces Desulfovibrio bacteria, increases cecal short-chain fatty acids, and is anti-inflammatory in mice. Am. J. Physiol.—Gastrointest. Liver Physiol. 2015, 309, G590–G601. [Google Scholar] [CrossRef]
- Ney, D.M.; Etzel, M.R. Designing medical foods for inherited metabolic disorders: Why intact protein is superior to amino acids. Curr. Opin. Biotechnol. 2017, 44, 39–45. [Google Scholar] [CrossRef]
- Ballatore, M.B.; Bettiol, M.D.R.; Braber, N.L.V.; Aminahuel, C.A.; Rossi, Y.E.; Petroselli, G.; Erra-Balsells, R.; Cavaglieri, L.R.; Montenegro, M.A. Antioxidant and cytoprotective effect of peptides produced by hydrolysis of whey protein concentrate with trypsin. Food Chem. 2020, 319, 126472. [Google Scholar] [CrossRef]
- Ali, E.; Nielsen, S.D.; Aal, S.A.-E.; El-Leboudy, A.; Saleh, E.; LaPointe, G. Use of Mass Spectrometry to Profile Peptides in Whey Protein Isolate Medium Fermented by Lactobacillus helveticus LH-2 and Lactobacillus acidophilus La-5. Front. Nutr. 2019, 6, 152. [Google Scholar] [CrossRef]
- Dinika, I.; Verma, D.K.; Balia, R.; Utama, G.L.; Patel, A.R. Potential of cheese whey bioactive proteins and peptides in the development of antimicrobial edible film composite: A review of recent trends. Trends Food Sci. Technol. 2020, 103, 57–67. [Google Scholar] [CrossRef]
- Elbarbary, H.A.; Ejima, A.; Sato, K. Generation of antibacterial peptides from crude cheese whey using pepsin and rennet enzymes at various pH conditions. J. Sci. Food Agric. 2018, 99, 555–563. [Google Scholar] [CrossRef]
- Osman, A.; Goda, H.A.; Abdel-Hamid, M.; Badran, S.M.; Otte, J. Antibacterial peptides generated by Alcalase hydrolysis of goat whey. LWT 2016, 65, 480–486. [Google Scholar] [CrossRef]
- Théolier, J.; Hammami, R.; Labelle, P.; Fliss, I.; Jean, J. Isolation and identification of antimicrobial peptides derived by peptic cleavage of whey protein isolate. J. Funct. Foods 2013, 5, 706–714. [Google Scholar] [CrossRef]
- Mazorra-Manzano, M.A.; Robles-Porchas, G.R.; González-Velázquez, D.A.; Torres-Llanez, M.J.; Martínez-Porchas, M.; García-Sifuentes, C.O.; González-Córdova, A.F.; Vallejo-Córdoba, B. Cheese Whey Fermentation by Its Native Microbiota: Proteolysis and Bioactive Peptides Release with ACE-Inhibitory Activity. Fermentation 2020, 6, 19. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslam, M.Z.; Firdos, S.; Li, Z.; Wang, X.; Liu, Y.; Qin, X.; Yang, S.; Ma, Y.; Xia, X.; Zhang, B.; et al. Detecting the Mechanism of Action of Antimicrobial Peptides by Using Microscopic Detection Techniques. Foods 2022, 11, 2809. https://doi.org/10.3390/foods11182809
Aslam MZ, Firdos S, Li Z, Wang X, Liu Y, Qin X, Yang S, Ma Y, Xia X, Zhang B, et al. Detecting the Mechanism of Action of Antimicrobial Peptides by Using Microscopic Detection Techniques. Foods. 2022; 11(18):2809. https://doi.org/10.3390/foods11182809
Chicago/Turabian StyleAslam, Muhammad Zohaib, Shumaila Firdos, Zhousi Li, Xiang Wang, Yangtai Liu, Xiaojie Qin, Shuo Yang, Yue Ma, Xuejuan Xia, Bolin Zhang, and et al. 2022. "Detecting the Mechanism of Action of Antimicrobial Peptides by Using Microscopic Detection Techniques" Foods 11, no. 18: 2809. https://doi.org/10.3390/foods11182809