
Sargassum thunbergii Extract Attenuates High-Fat Diet-Induced Obesity in Mice by Modulating AMPK Activation and the Gut Microbiota


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Extracts and HPLC Analysis
2.2. Cell Culture and Differentiation
2.3. Cell Viability Assay
2.4. Oil Red O Staining
2.5. Animal Studies
2.6. Histological Observations of Adipose Tissue and Liver
2.7. Blood Parameter Analysis
2.8. RT-PCR, Bacterial DNA Extraction, and 16S rRNA Gene Sequencing
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
3.1. Identification of Bioactive Compounds in S. thunbergii Extract
3.2. Effects of S. thunbergii Extract and Indole-2-Carboxaldehyde on the Adipogenic Differentiation of 3T3-L1 Pre-Adipocytes
3.3. Effects of S. thunbergii Extract on the Expression of Adipogenic and Thermogenic Genes and Proteins in Differentiated 3T3-L1 Cells
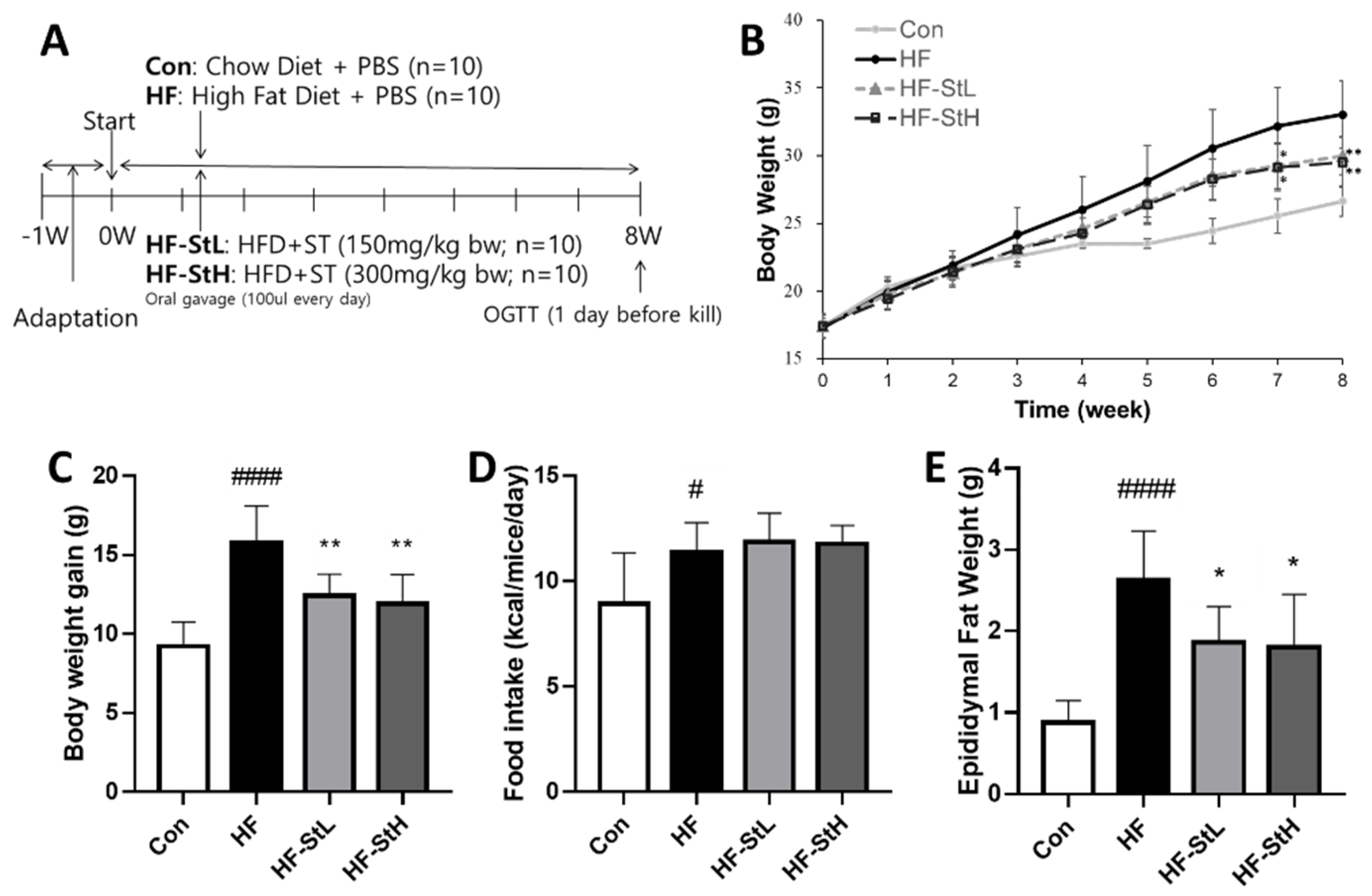
3.4. Effects of S. thunbergii Extract on Body and Organ Weights in High Fat-Fed C57BL/6 Mice
3.5. Effects of S. thunbergii Extract on Histology of Liver and Adipose Tissue in High-Fat-Fed C57BL/6 Mice
3.6. Effects of S. thunbergii Extract on the Serum Profiles in High-Fat-Fed C57BL/6 Mice
3.7. Effects of S. thunbergii Extract on AMPK Activation in White Adipose Tissue and Liver in High-Fat-Fed C57BL/6 Mice
3.8. Effects of S. thunbergii Extract on the Gut Microbiota Composition in High-Fat-Fed C57BL/6 Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, G.; Kim, K.; Wilding, J.; Federation, W.O. Obesity: A chronic relapsing progressive disease process. A position statement of the World Obesity Federation. Obes. Rev. 2017, 18, 715–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity: Implications for metabolic syndrome, diabetes, hypertension, dyslipidemia, atherosclerosis, and cancer. Obes. Res. Clin. Pract. 2013, 7, e330–e341. [Google Scholar] [CrossRef] [PubMed]
- Torres-Fuentes, C.; Schellekens, H.; Dinan, T.G.; Cryan, J.F. A natural solution for obesity: Bioactives for the prevention and treatment of weight gain. A review. Nutr. Neurosci. 2015, 18, 49–65. [Google Scholar] [CrossRef]
- Liu, S.; Chang, X.; Yu, J.; Xu, W. Cerasus humilis cherry polyphenol reduces high-fat diet-induced obesity in C57BL/6 mice by mitigating fat deposition, inflammation, and oxidation. J. Agric. Food Chem. 2020, 68, 4424–4436. [Google Scholar] [CrossRef]
- Jakab, J.; Miškić, B.; Mikšić, Š.; Juranić, B.; Ćosić, V.; Schwarz, D.; Včev, A. Adipogenesis as a potential anti-obesity target: A review of pharmacological treatment and natural products. Diabetes Metab. Syndr. Obes. 2021, 14, 67. [Google Scholar] [CrossRef] [PubMed]
- Batchuluun, B.; Pinkosky, S.L.; Steinberg, G.R. Lipogenesis inhibitors: Therapeutic opportunities and challenges. Nat. Rev. Drug Discov. 2022, 21, 283–305. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Cohen, P.; Spiegelman, B.M. Adaptive thermogenesis in adipocytes: Is beige the new brown? Genes Dev. 2013, 27, 234–250. [Google Scholar] [CrossRef] [Green Version]
- Cohen, P.; Kajimura, S. The cellular and functional complexity of thermogenic fat. Nat. Rev. Mol. Cell Biol. 2021, 22, 393–409. [Google Scholar] [CrossRef]
- Cantó, C.; Auwerx, J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 2009, 20, 98. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Liu, B.; Wei, H.; Cheng, K.W.; Chen, F. Extract of the Microalga Nitzschia laevis prevents high-fat-diet-induced obesity in mice by modulating the composition of gut microbiota. Mol. Nutr. Food Res. 2019, 63, 1800808. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The human intestinal microbiome in health and disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaiserman, A.M.; Koliada, A.K.; Marotta, F. Gut microbiota: A player in aging and a target for anti-aging intervention. Ageing Res. Rev. 2017, 35, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of diet on the gut microbiota: Rethinking intervention duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Reynés, B.; Palou, M.; Rodriguez, A.M.; Palou, A. Regulation of adaptive thermogenesis and browning by prebiotics and postbiotics. Front. Physiol. 2019, 9, 1908. [Google Scholar] [CrossRef]
- Carrera-Quintanar, L.; Lopez Roa, R.I.; Quintero-Fabián, S.; Sánchez-Sánchez, M.A.; Vizmanos, B.; Ortuño-Sahagún, D. Phytochemicals that influence gut microbiota as prophylactics and for the treatment of obesity and inflammatory diseases. Mediat. Inflamm. 2018, 2018, 9734845. [Google Scholar] [CrossRef]
- Azhar, Y.; Parmar, A.; Miller, C.N.; Samuels, J.S.; Rayalam, S. Phytochemicals as novel agents for the induction of browning in white adipose tissue. Nutr. Metab. 2016, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef] [Green Version]
- Forbes-Hernández, T.Y.; Cianciosi, D.; Ansary, J.; Mezzetti, B.; Bompadre, S.; Quiles, J.L.; Giampieri, F.; Battino, M. Strawberry (Fragaria× ananassa cv. Romina) methanolic extract promotes browning in 3T3-L1 cells. Food Funct. 2020, 11, 297–304. [Google Scholar] [CrossRef]
- Lee, H.-G.; Lu, Y.A.; Li, X.; Hyun, J.-M.; Kim, H.-S.; Lee, J.J.; Kim, T.H.; Kim, H.M.; Kang, M.-C.; Jeon, Y.-J. Anti-Obesity effects of Grateloupia elliptica, a red seaweed, in mice with high-fat diet-induced obesity via suppression of adipogenic factors in white adipose tissue and increased thermogenic factors in brown adipose tissue. Nutrients 2020, 12, 308. [Google Scholar] [CrossRef] [Green Version]
- Wan-Loy, C.; Siew-Moi, P. Marine algae as a potential source for anti-obesity agents. Mar. Drugs 2016, 14, 222. [Google Scholar] [CrossRef] [Green Version]
- Park, P.-J.; Heo, S.-J.; Park, E.-J.; Kim, S.-K.; Byun, H.-G.; Jeon, B.-T.; Jeon, Y.-J. Reactive oxygen scavenging effect of enzymatic extracts from Sargassum thunbergii. J. Agric. Food Chem. 2005, 53, 6666–6672. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-G.; Kang, H. Neuroprotective effect of Sargassum thunbergii (Mertens ex Roth) Kuntze in activated murine microglial cells. Trop. J. Pharm. Res. 2015, 14, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-C.; Ding, Y.; Kim, E.-A.; Choi, Y.K.; De Araujo, T.; Heo, S.-J.; Lee, S.-H. Indole derivatives isolated from brown alga Sargassum thunbergii inhibit adipogenesis through AMPK activation in 3T3-L1 preadipocytes. Mar. Drugs 2017, 15, 119. [Google Scholar] [CrossRef]
- Kang, M.-C.; Lee, H.-G.; Kim, H.-S.; Song, K.-M.; Chun, Y.-G.; Lee, M.H.; Kim, B.-K.; Jeon, Y.-J. Anti-obesity effects of Sargassum thunbergii via downregulation of adipogenesis gene and upregulation of thermogenic genes in high-fat diet-induced obese mice. Nutrients 2020, 12, 3325. [Google Scholar] [CrossRef] [PubMed]
- Swain, T.; Hillis, W. The phenolic constituents of Prunus domestica. I.—The quantitative analysis of phenolic constituents. J. Sci. Food Agric. 1959, 10, 63–68. [Google Scholar] [CrossRef]
- Cassano, A.; Conidi, C.; Ruby-Figueroa, R. Recovery of flavonoids from orange press liquor by an integrated membrane process. Membranes 2014, 4, 509–524. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.-J.; Kim, K.-J.; Choi, J.; Koh, E.-J.; Lee, B.-Y. Spirulina maxima extract reduces obesity through suppression of adipogenesis and activation of browning in 3T3-L1 cells and high-fat diet-induced obese mice. Nutrients 2018, 10, 712. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Pan, Y.; Yu, N.; Bai, Y.; Ma, R.; Mo, F.; Zuo, J.; Chen, B.; Jia, Q.; Zhang, D. Curcumin improves adipocytes browning and mitochondrial function in 3T3-L1 cells and obese rodent model. R. Soc. Open Sci. 2021, 8, 200974. [Google Scholar] [CrossRef]
- Liao, W.; Yin, X.; Li, Q.; Zhang, H.; Liu, Z.; Zheng, X.; Zheng, L.; Feng, X. Resveratrol-induced white adipose tissue browning in obese mice by remodeling fecal microbiota. Molecules 2018, 23, 3356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, S.-M.; Chen, S.; Park, K.W.; Yoon, J.-H. Isolation of lactic acid bacteria from kimchi and screening of Lactobacillus sakei ADM14 with anti-adipogenic effect and potential probiotic properties. LWT 2020, 126, 109296. [Google Scholar] [CrossRef]
- Zhu, Y.; Guo, J.; Hu, X.; Liu, J.; Li, S.; Wang, J. Eckol protects against acute experimental colitis in mice: Possible involvement of Reg3g. J. Funct. Foods 2020, 73, 104088. [Google Scholar] [CrossRef]
- Li, K.K.; Liu, C.L.; Shiu, H.T.; Wong, H.L.; Siu, W.S.; Zhang, C.; Han, X.Q.; Ye, C.X.; Leung, P.C.; Ko, C.H. Cocoa tea (Camellia ptilophylla) water extract inhibits adipocyte differentiation in mouse 3T3-L1 preadipocytes. Sci. Rep. 2016, 6, 20172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qureshi, K.; Abrams, G.A. Metabolic liver disease of obesity and role of adipose tissue in the pathogenesis of nonalcoholic fatty liver disease. World J. Gastroenterol. 2007, 13, 3540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delzenne, N.M.; Cani, P.D. Interaction between obesity and the gut microbiota: Relevance in nutrition. Annu. Rev. Nutr. 2011, 31, 15–31. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-S.; Han, J.-M.; Surendhiran, D.; Chun, B.-S. Physicochemical and biofunctional properties of Sargassum thunbergii extracts obtained from subcritical water extraction and conventional solvent extraction. J. Supercrit. Fluids 2022, 182, 105535. [Google Scholar] [CrossRef]
- Rosen, E.D.; Hsu, C.-H.; Wang, X.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBPα induces adipogenesis through PPARγ: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Zhang, L.; Li, B.; Jiang, H.; Duan, Y.; Xie, Z.; Shuai, L.; Li, J.; Li, J. AMP-activated protein kinase (AMPK) regulates energy metabolism through modulating thermogenesis in adipose tissue. Front. Physiol. 2018, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Klop, B.; Elte, J.W.F.; Castro Cabezas, M. Dyslipidemia in obesity: Mechanisms and potential targets. Nutrients 2013, 5, 1218–1240. [Google Scholar] [CrossRef] [Green Version]
- Landecho, M.F.; Tuero, C.; Valentí, V.; Bilbao, I.; de la Higuera, M.; Frühbeck, G. Relevance of leptin and other adipokines in obesity-associated cardiovascular risk. Nutrients 2019, 11, 2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, G.R.; Carling, D. AMP-activated protein kinase: The current landscape for drug development. Nat. Rev. Drug Discov. 2019, 18, 527–551. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.; Pan, D. Regulation of fatty acid synthesis and oxidation by the AMP-activated protein kinase. Biochem. Soc. Trans. 2002, 30, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Schug, T.T.; Li, X. Sirtuin 1 in lipid metabolism and obesity. Ann. Med. 2011, 43, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Yang, G.; Kim, Y.; Kim, J.; Ha, J. AMPK activators: Mechanisms of action and physiological activities. Exp. Mol. Med. 2016, 48, e224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, C.D. The gut microbiome and its role in obesity. Nutr. Today 2016, 51, 167. [Google Scholar] [CrossRef] [Green Version]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The firmicutes/bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Yoshida, N.; Yamashita, T.; Osone, T.; Hosooka, T.; Shinohara, M.; Kitahama, S.; Sasaki, K.; Sasaki, D.; Yoneshiro, T.; Suzuki, T. Bacteroides spp. promotes branched-chain amino acid catabolism in brown fat and inhibits obesity. Iscience 2021, 24, 103342. [Google Scholar] [CrossRef]
- Chatel, J.-M.; Maioli, T.U.; Borras-Nogues, E.; Barbosa, S.C.; Martins, V.D.; Torres, L.; Azevedo, V.A.D.C. Possible benefits of Faecalibacterium prausnitzii for obesity-associated gut disorders. Front. Pharmacol. 2021, 21, 740636. [Google Scholar]
- Bojović, K.; Ignjatović, Ð.; Soković Bajić, S.; Vojnović Milutinović, D.; Tomić, M.; Golić, N.; Tolinački, M. Gut microbiota dysbiosis associated with altered production of short chain fatty acids in children with neurodevelopmental disorders. Front. Cell. Infect. Microbiol. 2020, 10, 223. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Jin, Y.; Hofseth, A.B.; Pena, E.; Habiger, J.; Chumanevich, A.; Poudyal, D.; Nagarkatti, M.; Nagarkatti, P.S.; Singh, U.P.; et al. Resveratrol suppresses colitis and colon cancer associated with colitis. Cancer Prev. Res. 2010, 3, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mottillo, E.P.; Desjardins, E.M.; Crane, J.D.; Smith, B.K.; Green, A.E.; Ducommun, S.; Henriksen, T.I.; Rebalka, I.A.; Razi, A.; Sakamoto, K. Lack of adipocyte AMPK exacerbates insulin resistance and hepatic steatosis through brown and beige adipose tissue function. Cell Metab. 2016, 24, 118–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| β-actin | CCCTACAGTGCTGTGGGTTT | GAGACATGCAAGGAGTGCAA |
| Ampk | TGTTCCAGCAGATCCTTTCC | ATAATTGGGTGAGCCACAGC |
| C/ebpα | ATCAGCGCCTACATTGATCC | TTGCTTGGCTGTCGTAGATG |
| Pparγ | CCCTGGCAAAGCATTTGTAT | GAAACTGGCACCCTTGAAAA |
| Pgc1α | AATGCAGCGGTCTTAGCACT | GTGTGAGGAGGGTCATCGTT |
| Pparα | TCTTCACGATGCTGTCCTCCT | CTATGTTTAGAAGGCCAGGC |
| Sirt1 | AGTTCCAGCCGTCTCTGTGT | CTCCACGAACAGCTTCACAA |
| Srebp-1 | GAGCCTTCAGACACGTCCTC | ACTCTTCTGGTGTGGCTGCT |
| Ucp1 | CTGCCAGGACAGTACCCAAG | GCCACAAACCCTTTGAAAAA |
| Target | Forward Primer | Reverse Primer | Reference |
|---|---|---|---|
| Uni (F341/R518) | CCTACGGGAGGCAGCAGT | ATTACCGCGGCTGCTGG | Lubbs (2009) |
| Akkermansia muciniphila | CTGAACCAGCCAAGTAGCG | CCGCAAACTTTCACAACTGACTTA | Collado (2007) |
| Bacteroides vulgatus | GCATCATGAGTCCGCATGTTC | TCCATACCCGACTTTATTCCTT | Wang (1996) |
| Faecalibacterium prausnitzii | AGATGGCCTCGCGTCCGA | CCGAAGACCTTCTTCCTCC | Wang (1996) |
| Lactobacillus reuteri | GCCGCCTAAGGTGGGACAGAT | AACACTCAAGGATTGTCTGA | Walter (2000) |
| Lactococcus lactis | TGAAGAATTGATGGAACTCG | CATTGTGGTTCACCGTTC | Bachmann (2015) |
| Romboutsia ilealis | GGGGCTAGCGTTATTCCGAA | CACCTGTCACTTCTGTCCCC | Designed in this study |
| Peak | Compound Name | Molecular Formula | Molecular Weight | Retention Time (min) | Calcd. m/z [M + H]+ | Content * |
|---|---|---|---|---|---|---|
| 1 | L-rhamnulose | C6H12O5 | 164.16 | 0.391 | 165.0546 | 1.53 |
| 2 | Dulcitol | C6H14O6 | 182.17 | 0.391 | 183.0863 | 1.53 |
| 3 | p-Coumaric acid | C9H18O3 | 164.16 | 0.857 | 165.0757 | 0.64 |
| 4 | Indole-2-carboxaldehyde | C9H7NO | 145.16 | 2.614 | 146.0600 | 0.77 |
| 5 | Isovalerycanitine | C12H23NO4 | 245.31 | 3.085 | 246.1700 | 0.85 |
| 6 | Loliolide | C11H16O3 | 196.24 | 3.689 | 197.1172 | 0.49 |
| 7 | Cinnamic acid | C9H8O2 | 148.16 | 9.232 | 149.0597 | 0.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.; Yan, J.; Bak, J.; Park, J.; Lee, H.; Kim, H. Sargassum thunbergii Extract Attenuates High-Fat Diet-Induced Obesity in Mice by Modulating AMPK Activation and the Gut Microbiota. Foods 2022, 11, 2529. https://doi.org/10.3390/foods11162529
Kim D, Yan J, Bak J, Park J, Lee H, Kim H. Sargassum thunbergii Extract Attenuates High-Fat Diet-Induced Obesity in Mice by Modulating AMPK Activation and the Gut Microbiota. Foods. 2022; 11(16):2529. https://doi.org/10.3390/foods11162529
Chicago/Turabian StyleKim, Dahee, Jing Yan, Jinwoo Bak, Jumin Park, Heeseob Lee, and Hyemee Kim. 2022. "Sargassum thunbergii Extract Attenuates High-Fat Diet-Induced Obesity in Mice by Modulating AMPK Activation and the Gut Microbiota" Foods 11, no. 16: 2529. https://doi.org/10.3390/foods11162529