
Effect of Coffee on Lipopolysaccharide-Induced Immortalized Human Oral Keratinocytes

 , and
, and 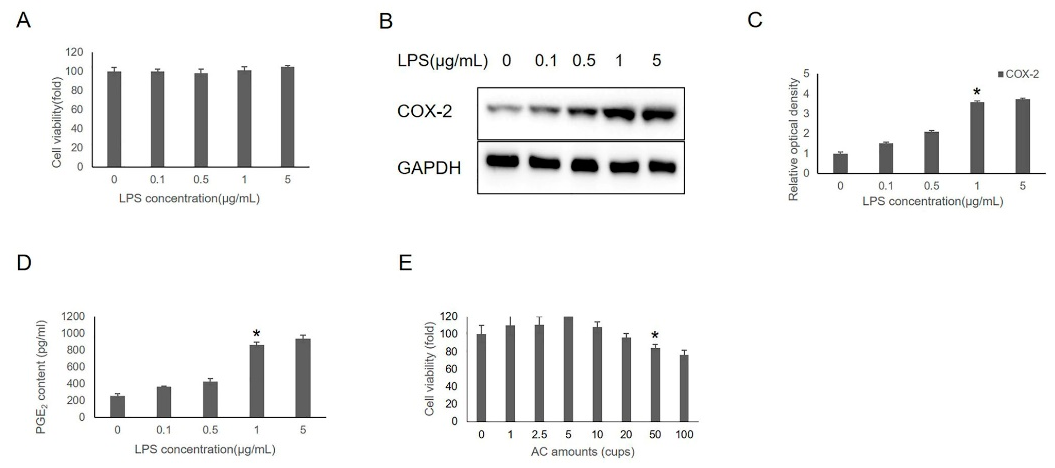{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstracts
1. Introduction
2. Material and Methods
2.1. Chemicals and Antibodies
2.2. Preparation of AC
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. PGE2 Release Assay
2.6. Cellular Reactive Oxygen Species (ROS) Determination
2.7. Immunofluorescence Staining
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Effects of AC on IHOK Viability
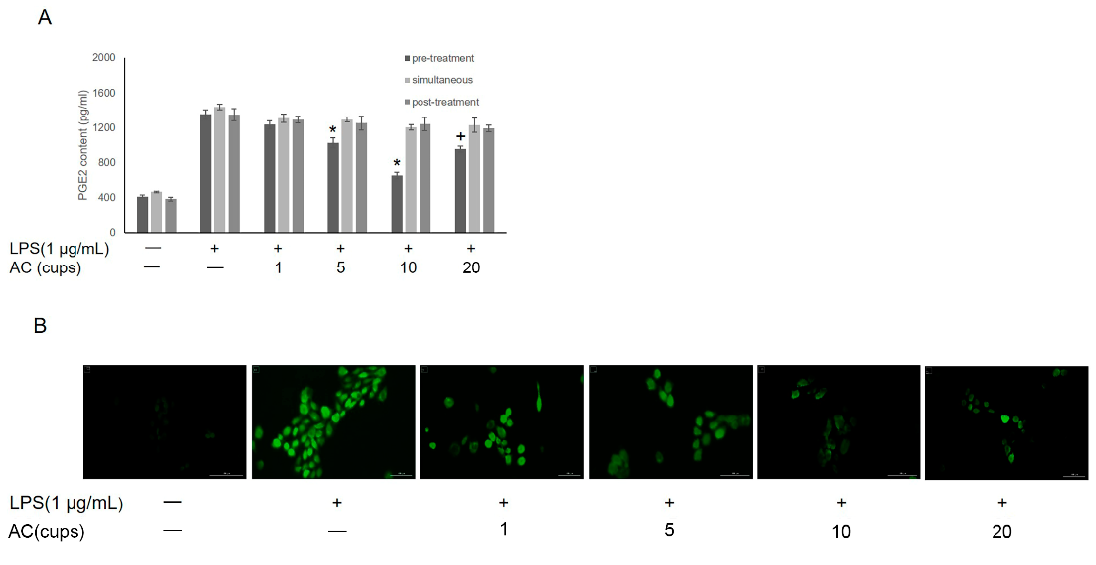
3.2. Effects of AC on LPS-Induced Inflammation in IHOKs
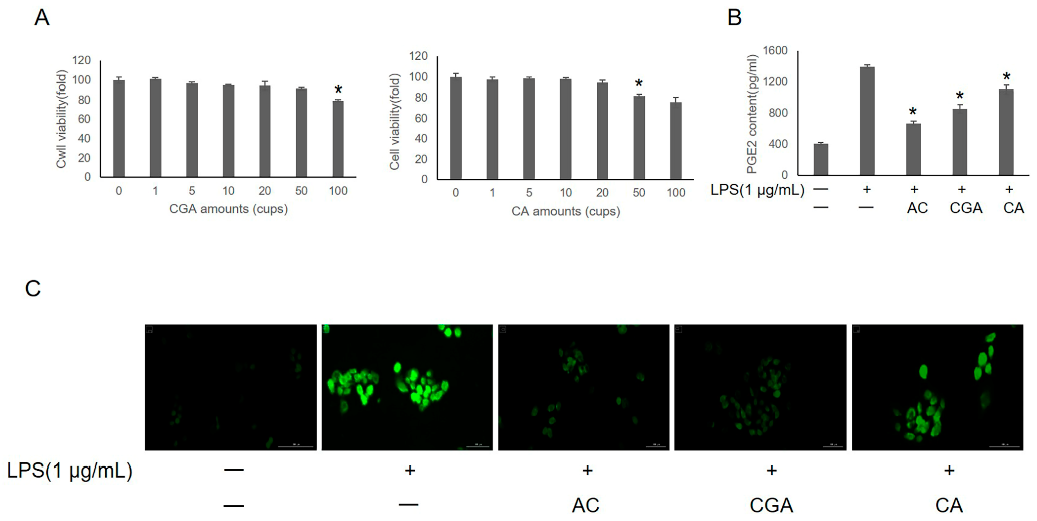
3.3. Effects of CGA and CA on LPS-Induced IHOKs
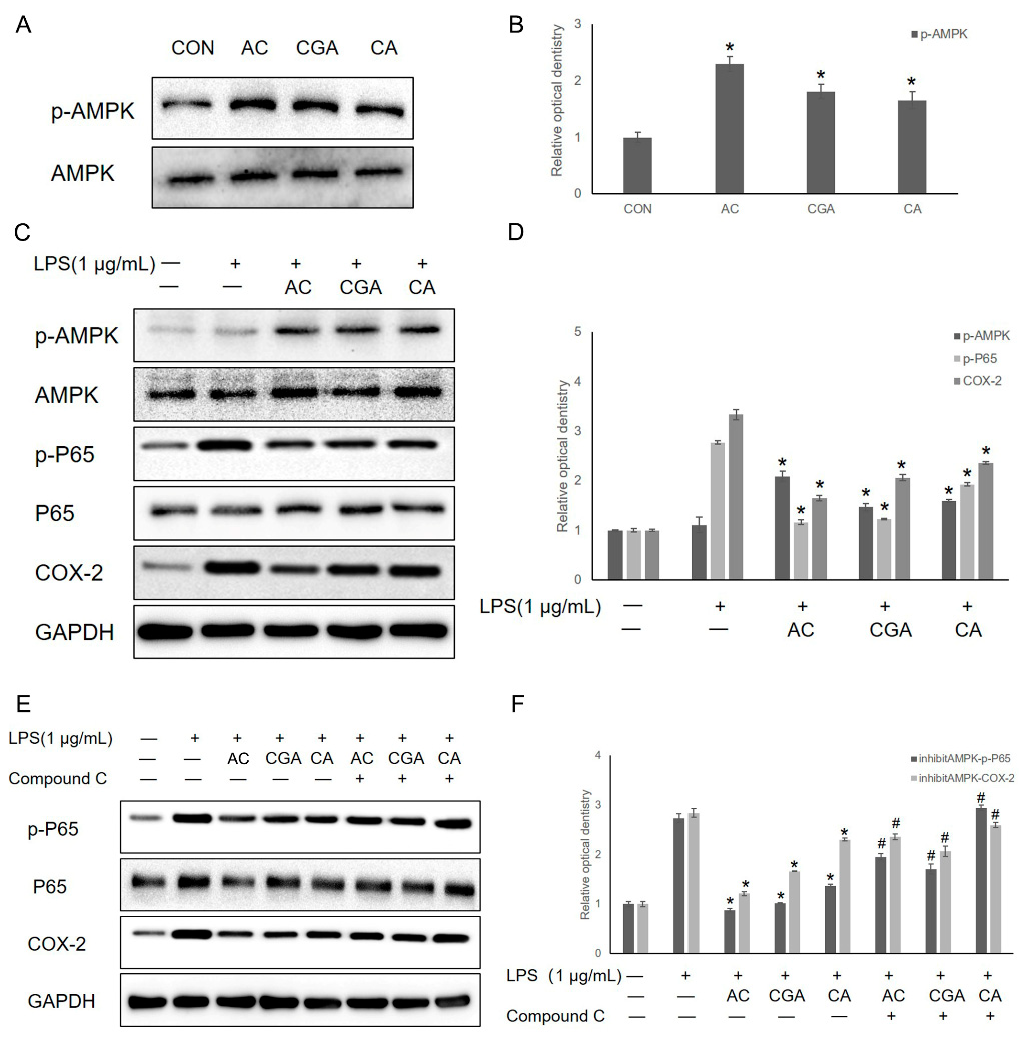
3.4. AC, CGA, and CA Inhibit LPS-Induced Inflammation by Activating the AMPK Pathway
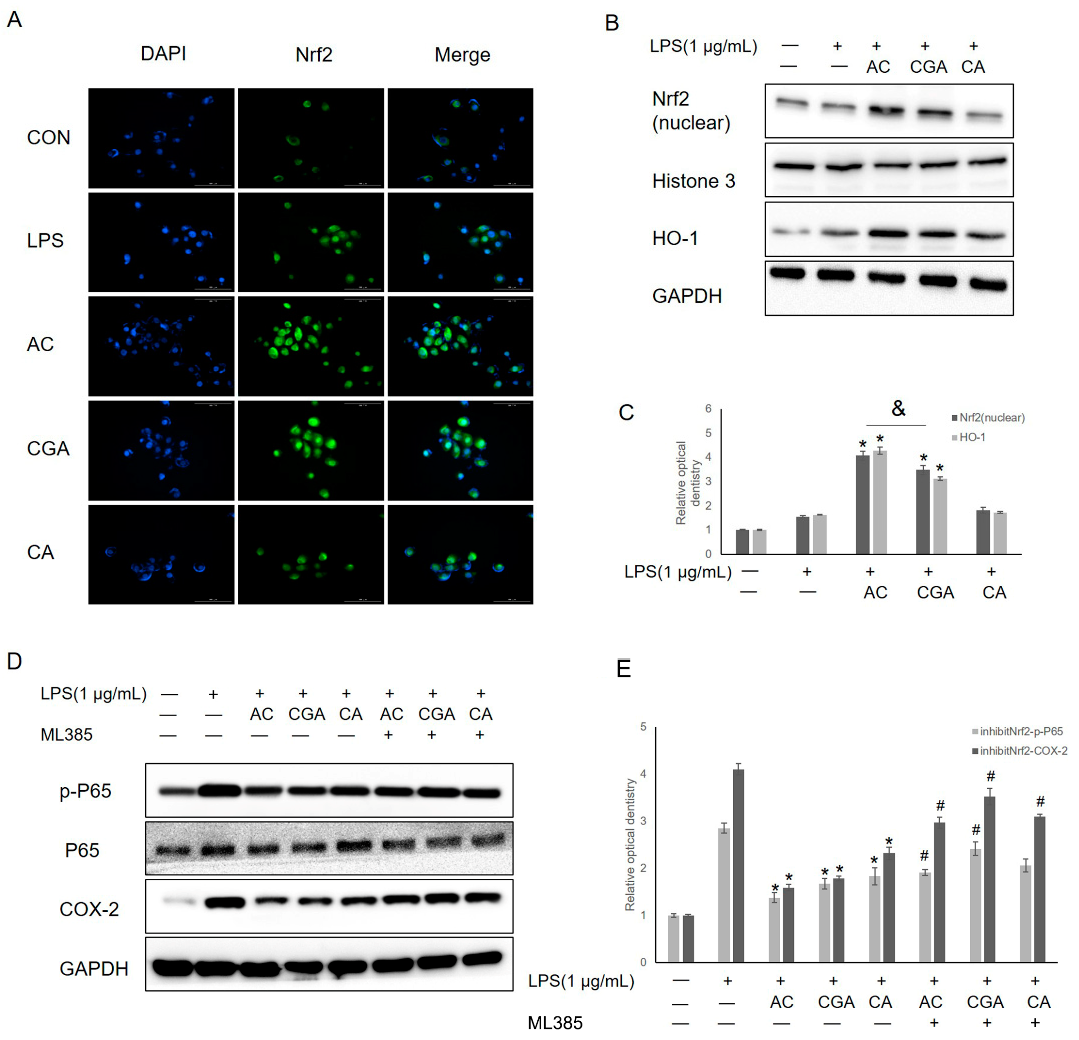
3.5. AC and CGA Inhibit LPS-Induced Inflammation by Activating the Nrf2/HO-1 Pathway
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ceuta Group. The Coffee Market in 2019; CG: Bournemouth, UK, 2019; Available online: https://www.ceutagroup.com/the-coffee-market-in-2019 (accessed on 9 May 2019).
- Ludwig, I.A.; Clifford, M.N.; Lean, M.E.J.; Ashihara, H.; Crozier, A. Coffee: Biochemistry and potential impact on health. Food Funct. 2014, 5, 1695–1717. [Google Scholar] [CrossRef] [PubMed]
- Wierzejska, R. Coffee consumption vs. cancer risk—A review of scientific data. Rocz. Panstw. Zakl. Hig. 2015, 66, 293–298. [Google Scholar] [PubMed]
- Ding, M.; Bhupathiraju, S.N.; Satija, A.; van Dam, R.; Hu, F.B. Long-Term Coffee Consumption and Risk of Cardiovascular Disease: A systematic review and a dose-response meta-analysis of prospective cohort studies. Circulation 2014, 129, 643–659. [Google Scholar] [CrossRef] [Green Version]
- Navarro, A.M.; Martinez-Gonzalez, M.Á.; Gea, A.; Grosso, G.; Martín-Moreno, J.M.; Lopez-Garcia, E.; Martin-Calvo, N.; Toledo, E. Coffee consumption and total mortality in a Mediterranean prospective cohort. Am. J. Clin. Nutr. 2018, 108, 1113–1120. [Google Scholar] [CrossRef]
- Torres, D.M.; Harrison, S.A. Is It Time to Write a Prescription for Coffee? Coffee and Liver Disease. Gastroenterology 2013, 144, 670–672. [Google Scholar] [CrossRef]
- Huxley, R.; Lee, C.; Barzi, F.; Timmermeister, L.; Czernichow, S.; Perkovic, V.; Grobbee, D.E.; Batty, D.; Woodward, M. Coffee, Decaffeinated Coffee, and Tea Consumption in Relation to Incident Type 2 Diabetes Mellitus: A Systematic Review With Meta-analysis. Arch. Intern. Med. 2009, 169, 2053–2063. [Google Scholar] [CrossRef]
- Liu, R.; Guo, X.; Park, Y.; Huang, X.; Sinha, R.; Freedman, N.D.; Hollenbeck, A.R.; Blair, A.; Chen, H. Caffeine Intake, Smoking, and Risk of Parkinson Disease in Men and Women. Am. J. Epidemiol. 2012, 175, 1200–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Discacciati, A.; Orsini, N.; Wolk, A. Coffee consumption and risk of nonaggressive, aggressive and fatal prostate cancer—A dose–response meta-analysis. Ann. Oncol. 2014, 25, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Galeone, C.; Turati, F.; La Vecchia, C.; Tavani, A. Coffee consumption and risk of colorectal cancer: A meta-analysis of case–control studies. Cancer Causes Control 2010, 21, 1949–1959. [Google Scholar] [CrossRef]
- Je, Y.; Giovannucci, E. Coffee consumption and risk of endometrial cancer: Findings from a large up-to-date meta-analysis. Int. J. Cancer 2012, 131, 1700–1710. [Google Scholar] [CrossRef]
- Lee, W.J.; Zhu, B.T. Inhibition of DNA methylation by caffeic acid and chlorogenic acid, two common catechol-containing coffee polyphenols. Carcinogenesis 2005, 27, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higdon, J.V.; Frei, B. Coffee and Health: A Review of Recent Human Research. Crit. Rev. Food Sci. Nutr. 2006, 46, 101–123. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, P.; Jordan, S.; Eastwood, J.; Rotstein, J.; Hugenholtz, A.; Feeley, M. Effects of caffeine on human health. Food Addit. Contam. 2003, 20, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Franks, A.M.; Schmidt, J.M.; McCain, K.R.; Fraer, M. Comparison of the Effects of Energy Drink Versus Caffeine Supplementation on Indices of 24-Hour Ambulatory Blood Pressure. Ann. Pharmacother. 2012, 46, 192–199. [Google Scholar] [CrossRef]
- Eke, P.; Dye, B.; Wei, L.; Thornton-Evans, G.; Genco, R. Prevalence of Periodontitis in Adults in the United States: 2009 and 2010. J. Dent. Res. 2012, 91, 914–920. [Google Scholar] [CrossRef]
- Han, K.; Hwang, E.; Park, J.-B. Association between Consumption of Coffee and the Prevalence of Periodontitis: The 2008–2010 Korea National Health and Nutrition Examination Survey. PLoS ONE 2016, 11, e0158845. [Google Scholar] [CrossRef]
- Choi, H.M.; Han, K.; Park, Y.-G.; Park, J.-B. Associations Among Oral Hygiene Behavior and Hypertension Prevalence and Control: The 2008 to 2010 Korea National Health and Nutrition Examination Survey. J. Periodontol. 2015, 86, 866–873. [Google Scholar] [CrossRef]
- Javid, A.Z.; Seal, C.J.; Heasman, P.; Moynihan, P.J. Impact of a customised dietary intervention on antioxidant status, dietary intakes and periodontal indices in patients with adult periodontitis. J. Hum. Nutr. Diet. 2013, 27, 523–532. [Google Scholar] [CrossRef]
- Lacerda, S.A.; Matuoka, R.I.; Macedo, R.M.; Petenusci, S.O.; Campos, A.A.; Brentegani, L.G. Bone quality associated with daily intake of coffee: A biochemical, radiographic and histometric study. Braz. Dent. J. 2010, 21, 199–204. [Google Scholar] [CrossRef]
- Macedo, R.M.; Brentegani, L.G.; De Lacerda, S.A. Effects of Coffee Intake and Intraperitoneal Caffeine on Bone Repair Process—A Histologic and Histometric Study. Braz. Dent. J. 2015, 26, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Miyake, Y.; Sasaki, S.; Ohya, Y.; Matsunaga, I.; Yoshida, T.; Hirota, Y.; Oda, H.; Group, C.H.S. Beverage Consumption andthe Prevalence of Tooth Loss in Pregnant Japanese Women: The Osaka Maternal and Child Health Study. Hukuoka Acta Med. 2008, 99, 80–89. [Google Scholar] [PubMed]
- Machida, T.; Tomofuji, T.; Ekuni, D.; Azuma, T.; Takeuchi, N.; Maruyama, T.; Mizutani, S.; Kataoka, K.; Kawabata, Y.; Morita, M. Severe Periodontitis Is Inversely Associated with Coffee Consumption in the Maintenance Phase of Periodontal Treatment. Nutrients 2014, 6, 4476–4490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamagata, K. Do Coffee Polyphenols Have a Preventive Action on Metabolic Syndrome Associated Endothelial Dysfunctions? An Assessment of the Current Evidence. Antioxidants 2018, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janda, K.; Jakubczyk, K.; Baranowska-Bosiacka, I.; Kapczuk, P.; Kochman, J.; Rębacz-Maron, E.; Gutowska, I. Mineral Composition and Antioxidant Potential of Coffee Beverages Depending on the Brewing Method. Foods 2020, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Yoon, C.S.; Kim, M.K.; Kim, Y.S.; Lee, S.K. In vitro protein expression changes in RAW 264.7 cells and HUVECs treated with dialyzed coffee extract by immunoprecipitation high performance liquid chromatography. Sci. Rep. 2018, 8, 13841. [Google Scholar] [CrossRef]
- Olagnier, D.; Brandtoft, A.M.; Gunderstofte, C.; Villadsen, N.L.; Krapp, C.; Thielke, A.L.; Laustsen, A.; Peri, S.; Hansen, A.L.; Bonefeld, L.; et al. Nrf2 negatively regulates STING indicating a link between antiviral sensing and metabolic reprogramming. Nat. Commun. 2018, 9, 3506. [Google Scholar] [CrossRef] [Green Version]
- Hutachok, N.; Angkasith, P.; Chumpun, C.; Fucharoen, S.; Mackie, I.; Porter, J.; Srichairatanakool, S. Anti-Platelet Aggregation and Anti-Cyclooxygenase Activities for a Range of Coffee Extracts (Coffea arabica). Molecules 2020, 26, 10. [Google Scholar] [CrossRef]
- Hang, D.; Kværner, A.S.; Ma, W.; Hu, Y.; Tabung, F.K.; Nan, H.; Hu, Z.; Shen, H.; Mucci, L.A.; Chan, A.T.; et al. Coffee consumption and plasma biomarkers of metabolic and inflammatory pathways in US health professionals. Am. J. Clin. Nutr. 2019, 109, 635–647. [Google Scholar] [CrossRef]
- Socała, K.; Szopa, A.; Serefko, A.; Poleszak, E.; Wlaź, P. Neuroprotective Effects of Coffee Bioactive Compounds: A Review. Int. J. Mol. Sci. 2020, 22, 107. [Google Scholar] [CrossRef]
- Trevitt, J.; Kawa, K.; Jalali, A.; Larsen, C. Differential effects of adenosine antagonists in two models of parkinsonian tremor. Pharmacol. Biochem. Behav. 2009, 94, 24–29. [Google Scholar] [CrossRef]
- Arendash, G.W.; Cao, C. Caffeine and coffee as therapeutics against Alzheimer’s disease. J. Alzheimer’s Dis. 2010, 20, S117–S126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurminen, M.-L.; Niittynen, L.; Korpela, R.; Vapaatalo, H. Coffee, caffeine and blood pressure: A critical review. Eur. J. Clin. Nutr. 1999, 53, 831–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redondo, B.; Vera, J.; Molina, R.; Jiménez, R. Short-term effects of caffeine intake on anterior chamber angle and intraocular pressure in low caffeine consumers. Graefe’s Arch. Clin. Exp. Ophthalmol. 2020, 258, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Struppek, J.; Walther, C.; Bunte, K.; Zyriax, B.-C.; Wenzel, J.-P.; Senftinger, J.; Nikorowitsch, J.; Heydecke, G.; Seedorf, U.; Beikler, T.; et al. The association between coffee consumption and periodontitis: A cross-sectional study of a northern German population. Clin. Oral Investig. 2021, 26, 2421–2427. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Maruyama, T.; Yoneda, T.; Miyai, H.; Azuma, T.; Tomofuji, T.; Ekuni, D.; Morita, M. Effects of Coffee Intake on Oxidative Stress during Aging-related Alterations in Periodontal Tissue. In Vivo 2020, 34, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Ng, N.; Kaye, E.K.; Garcia, R.I. Coffee Consumption and Periodontal Disease in Males. J. Periodontol. 2014, 85, 1042–1049. [Google Scholar] [CrossRef]
- Zupo, R.; Castellana, F.; De Nucci, S.; Dibello, V.; Lozupone, M.; Giannelli, G.; De Pergola, G.; Panza, F.; Sardone, R.; Boeing, H. Beverages Consumption and Oral Health in the Aging Population: A Systematic Review. Front. Nutr. 2021, 8, 762383. [Google Scholar] [CrossRef]
- Tsou, S.-H.; Hu, S.-W.; Yang, J.-J.; Yan, M.; Lin, Y.-Y. Potential Oral Health Care Agent from Coffee Against Virulence Factor of Periodontitis. Nutrients 2019, 11, 2235. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Xu, M.; Qiu, Y.; Liao, M.; Zhang, Q.; Yang, L.; Zheng, G. Chlorogenic acid and caffeine combination attenuates adipogenesis by regulating fat metabolism and inhibiting adipocyte differentiation in 3T3-L1 cells. J. Food Biochem. 2021, 45, e13795. [Google Scholar] [CrossRef]
- Delitto, A.E.; Rocha, F.; Decker, A.M.; Amador, B.; Sorenson, H.L.; Wallet, S.M. MyD88-mediated innate sensing by oral epithelial cells controls periodontal inflammation. Arch. Oral Biol. 2018, 87, 125–130. [Google Scholar] [CrossRef]
- Hans, M.; Hans, V.M. Toll-like receptors and their dual role in periodontitis: A review. J. Oral Sci. 2011, 53, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorillo, L.; Cervino, G.; Laino, L.; D’Amico, C.; Mauceri, R.; Tozum, T.F.; Gaeta, M.; Cicciù, M. Porphyromonas gingivalis, Periodontal and Systemic Implications: A Systematic Review. Dent. J. 2019, 7, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, C.L.; Roche, H.M. Nutritional Modulation of AMPK-Impact upon Metabolic-Inflammation. Int. J. Mol. Sci. 2018, 19, 3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi-Tago, M.; Nonaka, Y.; Tago, K.; Takeda, M.; Ishihara, Y.; Sakai, A.; Matsutaka, M.; Kobata, K.; Tamura, H. Pyrocatechol, a component of coffee, suppresses LPS-induced inflammatory responses by inhibiting NF-κB and activating Nrf2. Sci. Rep. 2020, 10, 2584. [Google Scholar] [CrossRef]
- Hayakawa, S.; Ohishi, T.; Miyoshi, N.; Oishi, Y.; Nakamura, Y.; Isemura, M. Anti-Cancer Effects of Green Tea Epigallocatchin-3-Gallate and Coffee Chlorogenic Acid. Molecules 2020, 25, 4553. [Google Scholar] [CrossRef]
- Tamura, H. Molecular Basis of Preventive Effects of Habitual Coffee Intake against Chronic Diseases. Yakugaku Zasshi J. Pharm. Soc. Jpn. 2020, 140, 1351–1363. [Google Scholar] [CrossRef]
- Mansour, A.; Mohajeri-Tehrani, M.R.; Samadi, M.; Qorbani, M.; Merat, S.; Adibi, H.; Poustchi, H.; Hekmatdoost, A. Effects of supplementation with main coffee components including caffeine and/or chlorogenic acid on hepatic, metabolic, and inflammatory indices in patients with non-alcoholic fatty liver disease and type 2 diabetes: A randomized, double-blind, placebo-controlled, clinical trial. Nutr. J. 2021, 20, 35. [Google Scholar] [CrossRef]
- Watanabe, S.; Takahashi, T.; Ogawa, H.; Uehara, H.; Tsunematsu, T.; Baba, H.; Morimoto, Y.; Tsuneyama, K. Daily Coffee Intake Inhibits Pancreatic Beta Cell Damage and Nonalcoholic Steatohepatitis in a Mouse Model of Spontaneous Metabolic Syndrome, Tsumura-Suzuki Obese Diabetic Mice. Metab. Syndr. Relat. Disord. 2017, 15, 170–177. [Google Scholar] [CrossRef]
- Passos, C.; Costa, R.; Ferreira, S.; Lopes, G.; Cruz, M.; Coimbra, M. Role of Coffee Caffeine and Chlorogenic Acids Adsorption to Polysaccharides with Impact on Brew Immunomodulation Effects. Foods 2021, 10, 378. [Google Scholar] [CrossRef]
- Derry, C.J.; Derry, S.; Moore, R.A. Caffeine as an analgesic adjuvant for acute pain in adults. Cochrane Database Syst. Rev. 2014, 2014, CD009281. [Google Scholar] [CrossRef] [PubMed]
- Agotegaray, M.A.; Dennehy, M.; Boeris, M.A.; Grela, M.A.; Burrow, R.A.; Quinzani, O.V. Therapeutic properties, SOD and catecholase mimetic activities of novel ternary copper(II) complexes of the anti-inflammatory drug Fenoprofen with imidazole and caffeine. Polyhedron 2012, 34, 74–83. [Google Scholar] [CrossRef]
- Granados-Soto, V.; Castañeda-Hernández, G. A review of the pharmacokinetic and pharmacodynamic factors in the potentiation of the antinociceptive effect of nonsteroidal anti-inflammatory drugs by caffeine. J. Pharmacol. Toxicol. Methods 1999, 42, 67–72. [Google Scholar] [CrossRef]
- Poole, R.; Kennedy, O.J.; Roderick, P.; Fallowfield, J.A.; Hayes, P.C.; Parkes, J. Coffee consumption and health: Umbrella review of meta-analyses of multiple health outcomes. BMJ 2017, 359, j5024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, E.M.; Asamenew, G.; Kim, H.W.; Lee, S.H.; Yoo, S.-M.; Cho, S.-M.; Cha, Y.-S.; Kang, M.-S. Anti-Obesity Effects of Petasites japonicus (Meowi) Ethanol Extract on RAW 264.7 Macrophages and 3T3-L1 Adipocytes and Its Characterization of Polyphenolic Compounds. Nutrients 2020, 12, 1261. [Google Scholar] [CrossRef] [PubMed]
- Vamanu, E.; Gatea, F. Correlations between Microbiota Bioactivity and Bioavailability of Functional Compounds: A Mini-Review. Biomedicines 2020, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Castaldo, L.; Toriello, M.; Sessa, R.; Izzo, L.; Lombardi, S.; Narváez, A.; Ritieni, A.; Grosso, M. Antioxidant and Anti-Inflammatory Activity of Coffee Brew Evaluated after Simulated Gastrointestinal Digestion. Nutrients 2021, 13, 4368. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Kim, B.; Kim, O.; Yang, Y.; Liu, D.; Fu, W.; Ma, G.; Kim, Y.; Kim, O. Effect of Coffee on Lipopolysaccharide-Induced Immortalized Human Oral Keratinocytes. Foods 2022, 11, 2199. https://doi.org/10.3390/foods11152199
Song J, Kim B, Kim O, Yang Y, Liu D, Fu W, Ma G, Kim Y, Kim O. Effect of Coffee on Lipopolysaccharide-Induced Immortalized Human Oral Keratinocytes. Foods. 2022; 11(15):2199. https://doi.org/10.3390/foods11152199
Chicago/Turabian StyleSong, Jianan, Byunggook Kim, Oksu Kim, Ying Yang, Danyang Liu, Wenqi Fu, Guowu Ma, Young Kim, and Okjoon Kim. 2022. "Effect of Coffee on Lipopolysaccharide-Induced Immortalized Human Oral Keratinocytes" Foods 11, no. 15: 2199. https://doi.org/10.3390/foods11152199