
The Fate of IgE Epitopes and Coeliac Toxic Motifs during Simulated Gastrointestinal Digestion of Pizza Base

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Profiling of Wheat and Soybean Allergens’ Pfam Domain
2.2.2. Batch Digestion Model
2.2.3. Gel Electrophoresis
2.2.4. Protein Mass Spectrometry and Data Analysis
2.2.5. In Silico Digestion and Peptide Mapping
2.2.6. IgE-Binding Epitopes and Coeliac Toxic Motif Analysis
3. Results
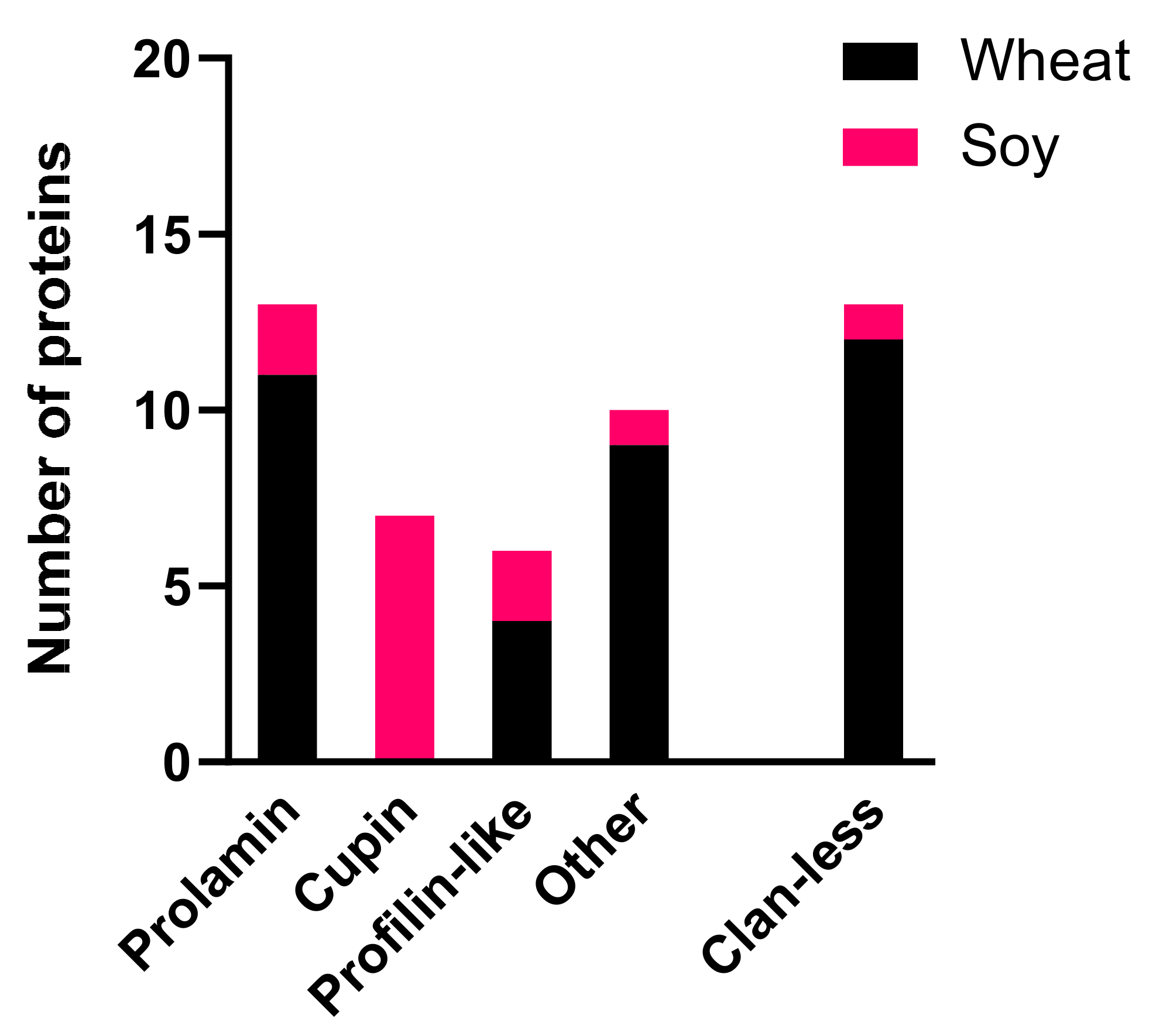
3.1. Pfam Domain Analysis of Curated Wheat and Soy Allergen Sequences
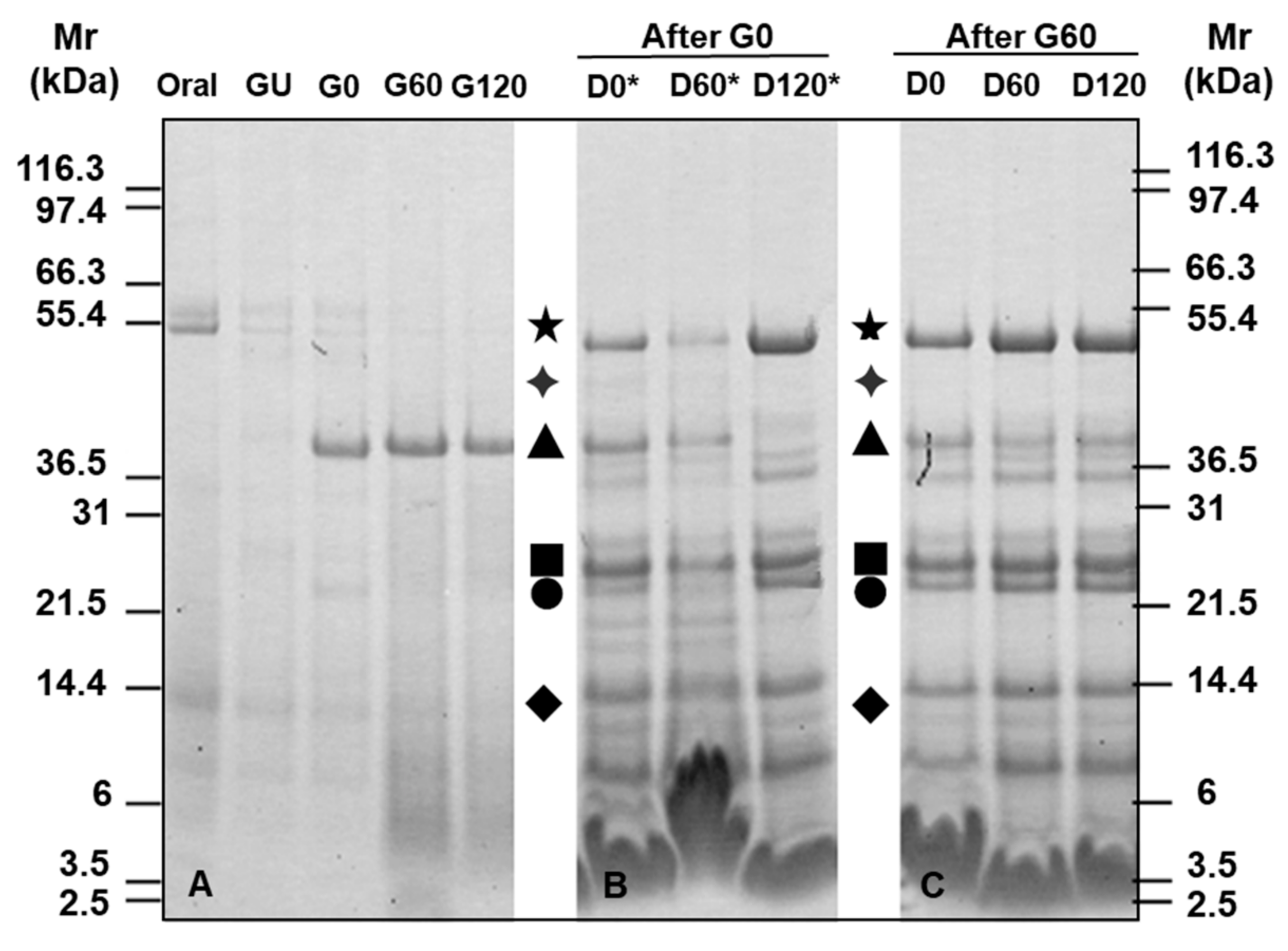
3.2. Gel Electrophoresis of Simulated Digests of Pizza Base
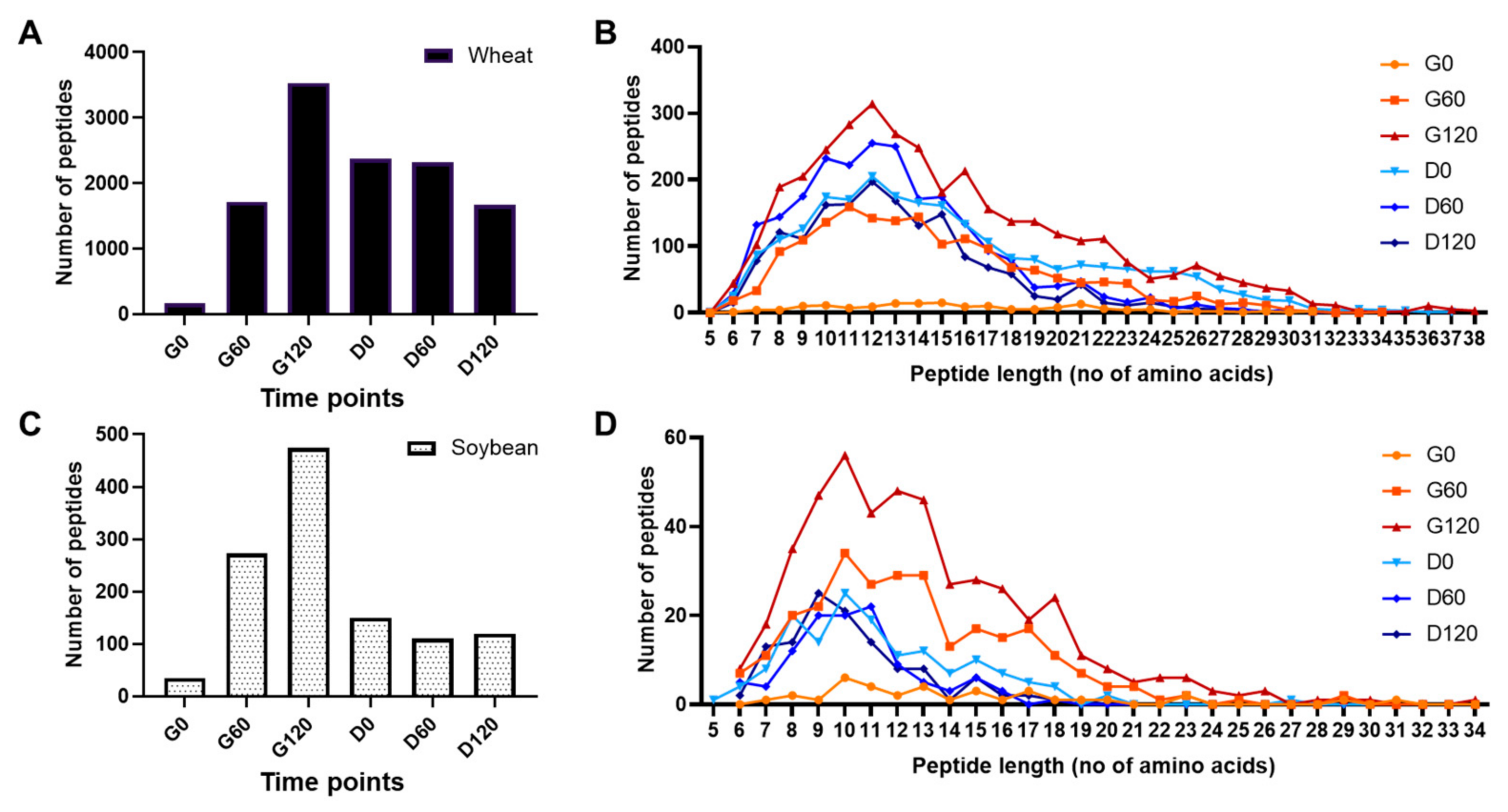
3.3. Peptide Profiling of Simulated Digests of Pizza Base by Mass Spectrometry
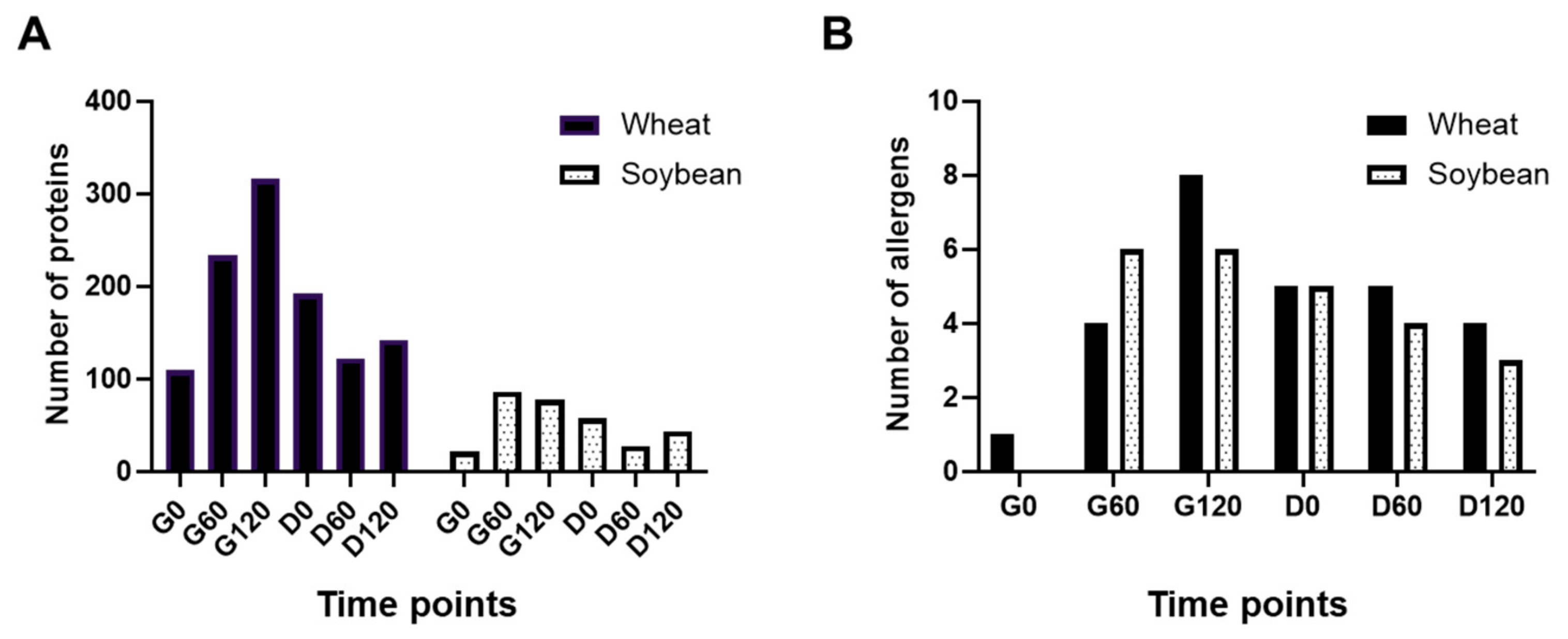
3.4. Protein Profiling of Simulated Digests of Pizza Base by Mass Spectrometry
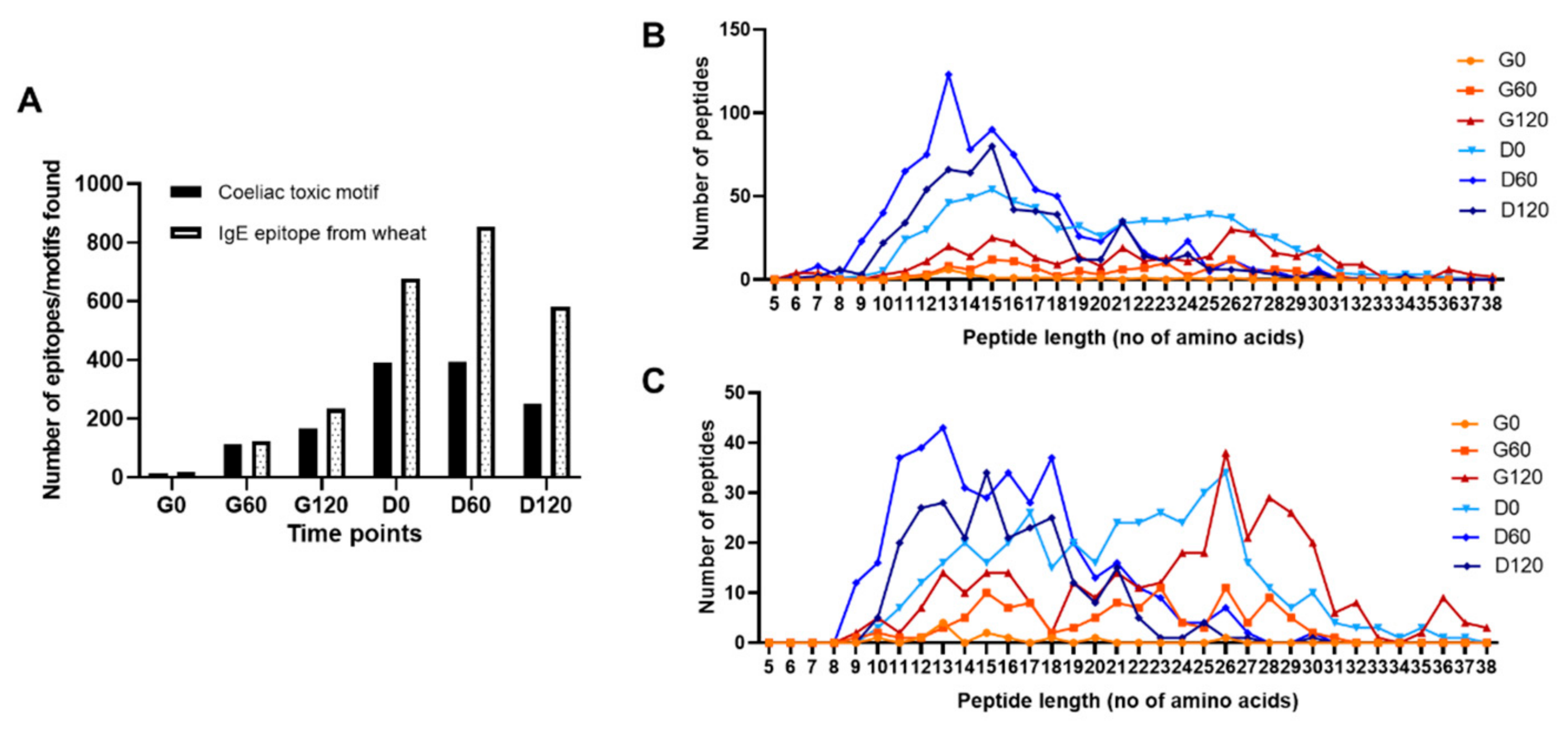
3.5. Bioaccessible Epitopes and Coeliac Toxic Motifs
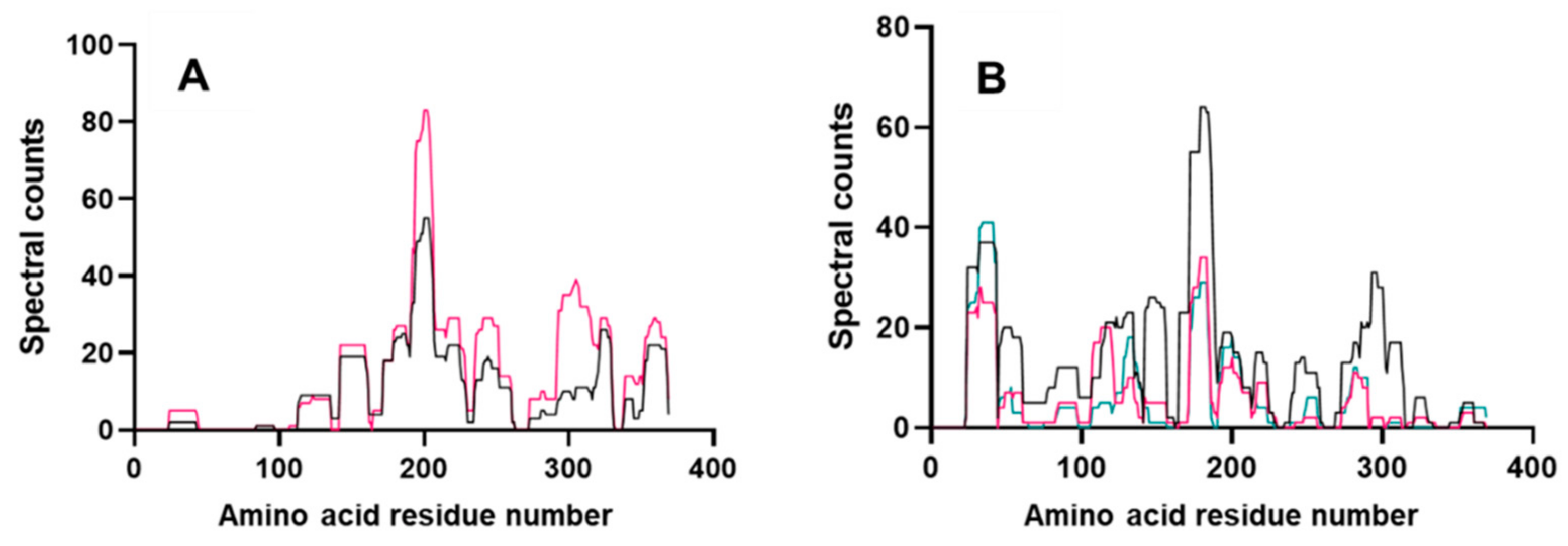
3.6. Peptide Mapping of a LMW-GS Allergen Tri a 36
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lyons, S.A.; Clausen, M.; Knulst, A.C.; Ballmer-Weber, B.K.; Fernandez-Rivas, M.; Barreales, L.; Bieli, C.; Dubakiene, R.; Fernandez-Perez, C.; Jedrzejczak-Czechowicz, M.; et al. Prevalence of Food Sensitization and Food Allergy in Children Across Europe. J. Allergy Clin. Immunol. Pract. 2020, 8, 2736–2746.e9. [Google Scholar] [CrossRef] [PubMed]
- Lyons, S.A.; Burney, P.G.J.; Ballmer-Weber, B.K.; Fernandez-Rivas, M.; Barreales, L.; Clausen, M.; Dubakiene, R.; Fernandez-Perez, C.; Fritsche, P.; Jedrzejczak-Czechowicz, M.; et al. Food Allergy in Adults: Substantial Variation in Prevalence and Causative Foods Across Europe. J. Allergy Clin. Immunol. Pract. 2019, 7, 1920–1928.e11. [Google Scholar] [CrossRef] [PubMed]
- Hefle, S.L.; Taylor, S.L. Food allergy and the food industry. Curr. Allergy Asthma Rep. 2004, 4, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.J.; Eller, E.; Mortz, C.G.; Brockow, K.; Bindslev-Jensen, C. Wheat-Dependent Cofactor-Augmented Anaphylaxis: A Prospective Study of Exercise, Aspirin, and Alcohol Efficacy as Cofactors. J. Allergy Clin. Immunol. Pract. 2019, 7, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Caio, G.; Volta, U.; Sapone, A.; Leffler, D.A.; de Giorgio, R.; Catassi, C.; Fasano, A. Celiac disease: A comprehensive current review. BMC Med. 2019, 17, 142. [Google Scholar] [CrossRef] [Green Version]
- Tye-Din, J.A.; Galipeau, H.J.; Agardh, D. Celiac Disease: A Review of Current Concepts in Pathogenesis, Prevention, and Novel Therapies. Front. Pediatr. 2018, 6, 350. [Google Scholar] [CrossRef]
- Baar, A.; Pahr, S.; Constantin, C.; Scheiblhofer, S.; Thalhamer, J.; Giavi, S.; Papadopoulos, N.G.; Ebner, C.; Mari, A.; Vrtala, S.; et al. Molecular and immunological characterization of Tri a 36, a low molecular weight glutenin, as a novel major wheat food allergen. J. Immunol. 2012, 189, 3018–3025. [Google Scholar] [CrossRef] [Green Version]
- Yokooji, T.; Kurihara, S.; Murakami, T.; Chinuki, Y.; Takahashi, H.; Morita, E.; Harada, S.; Ishii, K.; Hiragun, M.; Hide, M.; et al. Characterization of causative allergens for wheat-dependent exercise-induced anaphylaxis sensitized with hydrolyzed wheat proteins in facial soap. Allergol. Int. 2013, 62, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, H.; Kohno, K.; Morita, E. Molecular cloning, recombinant expression and IgE-binding epitope of omega-5 gliadin, a major allergen in wheat-dependent exercise-induced anaphylaxis. FEBS J. 2005, 272, 4431–4438. [Google Scholar] [CrossRef]
- Holzhauser, T.; Wackermann, O.; Ballmer-Weber, B.K.; Bindslev-Jensen, C.; Scibilia, J.; Perono-Garoffo, L.; Utsumi, S.; Poulsen, L.K.; Vieths, S. Soybean (Glycine max) allergy in Europe: Gly m 5 (beta-conglycinin) and Gly m 6 (glycinin) are potential diagnostic markers for severe allergic reactions to soy. J. Allergy Clin. Immunol. 2009, 123, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Klemans, R.J.; Knol, E.F.; Michelsen-Huisman, A.; Pasmans, S.G.; de Kruijf-Broekman, W.; Bruijnzeel-Koomen, C.A.; van Hoffen, E.; Knulst, A.C. Components in soy allergy diagnostics: Gly m 2S albumin has the best diagnostic value in adults. Allergy 2013, 68, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Mittag, D.; Vieths, S.; Vogel, L.; Becker, W.M.; Rihs, H.P.; Helbling, A.; Wuthrich, B.; Ballmer-Weber, B.K. Soybean allergy in patients allergic to birch pollen: Clinical investigation and molecular characterization of allergens. J. Allergy Clin. Immunol. 2004, 113, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Sollid, L.M.; Tye-Din, J.A.; Qiao, S.W.; Anderson, R.P.; Gianfrani, C.; Koning, F. Update 2020: Nomenclature and listing of celiac disease-relevant gluten epitopes recognized by CD4(+) T cells. Immunogenetics 2020, 72, 85–88. [Google Scholar] [CrossRef]
- Daly, M.; Bromilow, S.N.; Nitride, C.; Shewry, P.R.; Gethings, L.A.; Mills, E.N.C. Mapping Coeliac Toxic Motifs in the Prolamin Seed Storage Proteins of Barley, Rye, and Oats Using a Curated Sequence Database. Front. Nutr. 2020, 7, 87. [Google Scholar] [CrossRef] [PubMed]
- Palacin, A.; Varela, J.; Quirce, S.; del Pozo, V.; Tordesillas, L.; Barranco, P.; Fernandez-Nieto, M.; Sastre, J.; Diaz-Perales, A.; Salcedo, G. Recombinant lipid transfer protein Tri a 14: A novel heat and proteolytic resistant tool for the diagnosis of baker’s asthma. Clin. Exp. Allergy 2009, 39, 1267–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Angelis, E.; Pilolli, R.; Bavaro, S.L.; Monaci, L. Insight into the gastro-duodenal digestion resistance of soybean proteins and potential implications for residual immunogenicity. Food Funct. 2017, 8, 1599–1610. [Google Scholar] [CrossRef]
- Palosuo, K.; Varjonen, E.; Nurkkala, J.; Kalkkinen, N.; Harvima, R.; Reunala, T.; Alenius, H. Transglutaminase-mediated cross-linking of a peptic fraction of omega-5 gliadin enhances IgE reactivity in wheat-dependent, exercise-induced anaphylaxis. J. Allergy Clin. Immunol. 2003, 111, 1386–1392. [Google Scholar] [CrossRef]
- Shan, L.; Molberg, Ø.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.M.; Sollid, L.M.; Khosla, C. Structural basis for gluten intolerance in celiac sprue. Science 2002, 297, 2275–2279. [Google Scholar] [CrossRef] [Green Version]
- Mills, C.; Johnson, P.E.; Zuidmeer-Jongejan, L.; Critenden, R.; Wal, J.-M.; Asero, R. Effect of Processing on the Allergenicity of Foods. In Risk Management for Food Allergy; Academic Press: Cambridge, MA, USA, 2014; pp. 227–251. [Google Scholar]
- Pasini, G.; Simonato, B.; Giannattasio, M.; Peruffo, A.D.; Curioni, A. Modifications of wheat flour proteins during in vitro digestion of bread dough, crumb, and crust: An electrophoretic and immunological study. J. Agric. Food Chem. 2001, 49, 2254–2261. [Google Scholar] [CrossRef]
- Simonato, B.; Pasini, G.; Giannattasio, M.; Peruffo, A.D.; de Lazzari, F.; Curioni, A. Food allergy to wheat products: The effect of bread baking and in vitro digestion on wheat allergenic proteins. A study with bread dough, crumb, and crust. J. Agric. Food Chem. 2001, 49, 5668–5673. [Google Scholar] [CrossRef]
- Smith, F.; Pan, X.Y.; Bellido, V.; Toole, G.A.; Gates, F.K.; Wickham, M.S.J.; Shewry, P.R.; Bakalis, S.; Padfield, P.; Mills, E.N.C. Digestibility of gluten proteins is reduced by baking and enhanced by starch digestion. Mol. Nutr. Food Res. 2015, 59, 2034–2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattar, H.; Padfield, P.; Simpson, A.; Mills, E.N.C. The impact of a baked muffin matrix on the bioaccessibility and IgE reactivity of egg and peanut allergens. Food Chem. 2021, 362, 129879. [Google Scholar] [CrossRef] [PubMed]
- Rao, H.; Baricevic, I.; Bernard, H.; Smith, F.; Sayers, R.; Balasundaram, A.; Costello, C.A.; Padfield, P.; Semic-Jusufagic, A.; Simpson, A.; et al. The Effect of the Food Matrix on the In Vitro Bio-Accessibility and IgE Reactivity of Peanut Allergens. Mol. Nutr. Food Res. 2020, 64, e1901093. [Google Scholar] [CrossRef] [PubMed]
- Prandi, B.; Faccini, A.; Tedeschi, T.; Cammerata, A.; Sgrulletta, D.; D’Egidio, M.G.; Galaverna, G.; Sforza, S. Qualitative and quantitative determination of peptides related to celiac disease in mixtures derived from different methods of simulated gastrointestinal digestion of wheat products. Anal. Bioanal. Chem. 2014, 406, 4765–4775. [Google Scholar] [CrossRef] [PubMed]
- Mamone, G.; Nitride, C.; Picariello, G.; Addeo, F.; Ferranti, P.; Mackie, A. Tracking the fate of pasta (T. Durum semolina) immunogenic proteins by in vitro simulated digestion. J. Agric. Food Chem. 2015, 63, 2660–2667. [Google Scholar] [CrossRef]
- Pan, X.; Smith, F.; Cliff, M.; Capozzi, F.; Mills, E.N.C. The Application of Nutrimetabolomics to Investigating the Bioaccessibility of Nutrients in Ham Using a Batch in Vitro Digestion Model. Food Nutr. Sci. 2014, 5, 17–26. [Google Scholar]
- Glicerina, V.; Balestra, F.; Capozzi, F.; Rosa, M.D.; Romani, S. Influence of the addition of soy product and wheat fiber on rheological, textural, and other quality characteristics of pizza. J. Texture Stud. 2017, 49, 415–423. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [Green Version]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; Garcia-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- Matsuo, H.; Yokooji, T.; Taogoshi, T. Common food allergens and their IgE-binding epitopes. Allergol. Int. 2015, 64, 332–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, H.; Gagnon, C.; Cober, E.; Gleddie, S. Using patient serum to epitope map soybean glycinins reveals common epitopes shared with many legumes and tree nuts. Mol. Immunol. 2016, 70, 125–133. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Xi, J. Identification of an IgE epitope of soybean allergen Gly m Bd 60K. LWT 2020, 133, 110131. [Google Scholar] [CrossRef]
- Kern, K.; Havenith, H.; Delaroque, N.; Rautenberger, P.; Lehmann, J.; Fischer, M.; Spiegel, H.; Schillberg, S.; Ehrentreich-Foerster, E.; Aurich, S.; et al. The immunome of soy bean allergy: Comprehensive identification and characterization of epitopes. Clin. Exp. Allergy 2019, 49, 239–251. [Google Scholar] [CrossRef]
- Goodman, R.E.; Ebisawa, M.; Ferreira, F.; Sampson, H.A.; van Ree, R.; Vieths, S.; Baumert, J.L.; Bohle, B.; Lalithambika, S.; Wise, J.; et al. AllergenOnline: A peer-reviewed, curated allergen database to assess novel food proteins for potential cross-reactivity. Mol. Nutr. Food Res. 2016, 60, 1183–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomes, A.; Davies, J.M.; Gadermaier, G.; Hilger, C.; Holzhauser, T.; Lidholm, J.; Lopata, A.L.; Mueller, G.A.; Nandy, A.; Radauer, C.; et al. WHO/IUIS Allergen Nomenclature: Providing a common language. Mol. Immunol. 2018, 100, 3–13. [Google Scholar] [CrossRef]
- Bittner, C.; Peters, U.; Frenzel, K.; Musken, H.; Brettschneider, R. New wheat allergens related to baker’s asthma. J. Allergy Clin. Immunol. 2015, 136, 1416–14188.e2. [Google Scholar] [CrossRef] [Green Version]
- Jensen-Jarolim, E.; Schmid, B.; Bernier, F.; Berna, A.; Kinaciyan, T.; Focke, M.; Ebner, C.; Scheiner, O.; Boltz-Nitulescu, G. Allergologic exploration of germins and germin-like proteins, a new class of plant allergens. Allergy 2002, 57, 805–810. [Google Scholar] [CrossRef]
- Shewry, P.R.; D’Ovidio, R.; Lafiandra, D.; Jenkins, J.A.; Mills, E.N.C.; Békés, F. Wheat grain proteins. In Wheat: Chemistry and Technology; Khan, K., Shewry, R.P., Eds.; AACC International: Saint Paul, MN, USA, 2009; pp. 223–298. [Google Scholar]
- Bacala, R.; Fu, B.X.; Perreault, H.; Hatcher, D.W. C-terminal tyrosine removal from wheat low-molecular weight glutenin subunits (LMW-GS); biologically relevant or mistaken substrate? J. Cereal Sci. 2020, 95, 103060. [Google Scholar] [CrossRef]
- Liu, H.; Sadygov, R.G.; Yates, J.R., 3rd. A model for random sampling and estimation of relative protein abundance in shotgun proteomics. Anal. Chem. 2004, 76, 4193–4201. [Google Scholar] [CrossRef]
- Keil, B. Specificity of Proteolysis; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar]
- Ogilvie, O.; Roberts, S.; Sutton, K.; Domigan, L.; Larsen, N.; Gerrard, J.; Demarais, N. Proteomic modelling of gluten digestion from a physiologically relevant food system: A focus on the digestion of immunogenic peptides from wheat implicated in celiac disease. Food Chem. 2020, 333, 127466. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Genetically Modified Organisms; Naegeli, H.; Birch, A.N.; Casacuberta, J.; de Schrijver, A.; Gralak, M.A.; Guerche, P.; Jones, H.; Manachini, B.; Messean, A.; et al. Guidance on allergenicity assessment of genetically modified plants. EFSA J. 2017, 15, e04862. [Google Scholar]
- Dørum, S.; Arntzen, M.; Qiao, S.W.; Holm, A.; Koehler, C.J.; Thiede, B.; Sollid, L.M.; Fleckenstein, B. The preferred substrates for transglutaminase 2 in a complex wheat gluten digest are Peptide fragments harboring celiac disease T-cell epitopes. PLoS ONE 2010, 5, e14056. [Google Scholar] [CrossRef] [Green Version]
- Potocnakova, L.; Bhide, M.; Pulzova, L.B. An Introduction to B-Cell Epitope Mapping and In Silico Epitope Prediction. J. Immunol. Res. 2016, 2016, 6760830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, A.; Mills, E.N.C.; Koning, F.; Moreno, F.J. Safety Assessment of Immune-Mediated Adverse Reactions to Novel Food Proteins. Trends Biotechnol. 2019, 37, 796–800. [Google Scholar] [CrossRef]
- Fernandez, A.; Mills, E.N.C.; Koning, F.; Moreno, F.J. Allergenicity Assessment of Novel Food Proteins: What Should Be Improved? Trends Biotechnol. 2021, 39, 4–8. [Google Scholar] [CrossRef]
- Selb, R.; Wal, J.M.; Moreno, F.J.; Lovik, M.; Mills, C.; Hoffmann-Sommergruber, K.; Fernandez, A. Assessment of endogenous allergenicity of genetically modified plants exemplified by soybean—Where do we stand? Food Chem. Toxicol. 2017, 101, 139–148. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Cleavage Sites | ||||||
|---|---|---|---|---|---|---|
| Pepsin | Chymotrypsin | Pepsin/Chymotrypsin | Trypsin | Alternative | Total | |
| Predicted | 47 | 49 | 21 | 7 | n.a. | 82 |
| Experimentally Determined | ||||||
| G60 | 32 | n.a. | n.a. | n.a. | 53 | 85 |
| G120 | 38 | n.a. | n.a. | n.a. | 70 | 108 |
| D0 | 28 | 35 | 15 | 2 | 79 | 144 |
| D60 | 23 | 31 | 14 | 2 | 66 | 122 |
| D120 | 17 | 25 | 10 | 0 | 70 | 112 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daly, M.E.; Wang, K.; Pan, X.; Depau, R.L.; Marsh, J.; Capozzi, F.; Johnson, P.; Gethings, L.A.; Mills, E.N.C. The Fate of IgE Epitopes and Coeliac Toxic Motifs during Simulated Gastrointestinal Digestion of Pizza Base. Foods 2022, 11, 2000. https://doi.org/10.3390/foods11142000
Daly ME, Wang K, Pan X, Depau RL, Marsh J, Capozzi F, Johnson P, Gethings LA, Mills ENC. The Fate of IgE Epitopes and Coeliac Toxic Motifs during Simulated Gastrointestinal Digestion of Pizza Base. Foods. 2022; 11(14):2000. https://doi.org/10.3390/foods11142000
Chicago/Turabian StyleDaly, Matthew E., Kai Wang, Xiaoyan Pan, Rosa L. Depau, Justin Marsh, Francesco Capozzi, Phil Johnson, Lee A. Gethings, and E. N. Clare Mills. 2022. "The Fate of IgE Epitopes and Coeliac Toxic Motifs during Simulated Gastrointestinal Digestion of Pizza Base" Foods 11, no. 14: 2000. https://doi.org/10.3390/foods11142000