
Myristicin and Elemicin: Potentially Toxic Alkenylbenzenes in Food


Abstract
:1. Introduction
2. Occurrence of Myristicin and Elemicin
2.1. Myristicin and Elemicin in Nutmeg and Mace

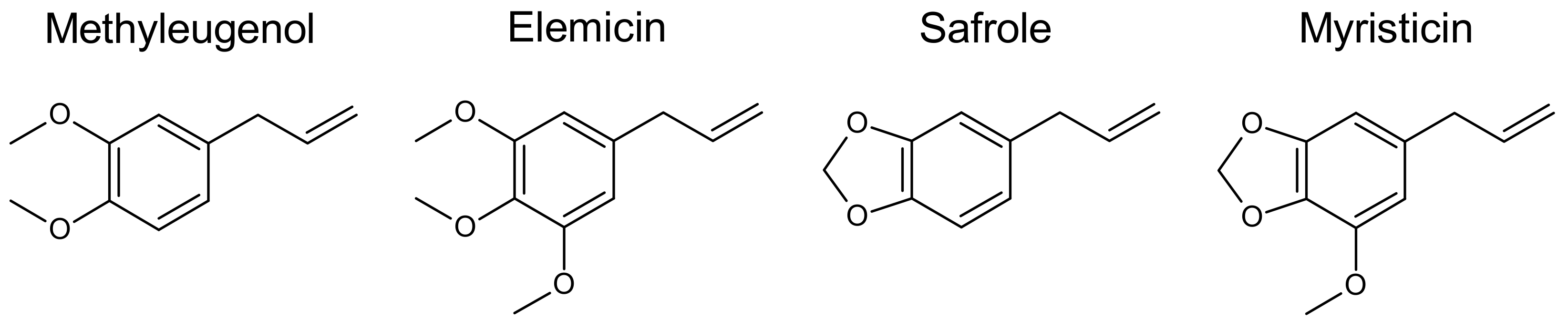{kind=link}
{kind=link}
| Source | Safrole | Myristicin | Methyleugenol | Elemicin |
|---|---|---|---|---|
| Nutmeg | 0.1–3.2% [19,20,22,23] | 0.5–12.4% [19,20,22,23] (16.9 ± 0.6 mg [26]) | 0.1–16.7% [19,20,22] | 0.3–4.6% [19,20,22] |
| Parsley | 20.3–94.1% (seed) [27,28]; 3.1–91.9% (leaf) [27]; 6.6–30.1% (root) [27] (1435 ppm [29]; 3.6–526 ppm (leaf) [30]) | |||
| Sweet fennel | 2.5–10% (root) [31,32] | |||
| Dill | 0.21% (seed) [33]; 4.38% (root) [32] | |||
| Parsnip | 18.3–66.2% (root) [34,35] (200 ppm (root) [36]) | |||
| Sweet basil | 9.24–87.04% [15]; 0.03% (flower) [37,38]; 0.06% (stem) [37,38]; 0.18–76% (leaf) [37,38,39] | 0.30% (stem) [37,38] | ||
| Carrot | 34.4% (leaf) [40]; 43.9% (fruit) [40]; 0.4–29.7% (root) [41,42]; (0.5–15 ppm (root) [43]) | 2.51% (fruit) [44] | 1.4–35.3% [45]; 32.89% (fruit) [44] | |
| Pepper (Piper) | 0.2–3.0 mg/kg (fruit) [46]; <3% (fruit) [47]; 4.81% (fruit) [48]; 24% [49]; 49% [50]; 64–98% [51] | <1% (fruit) [47]; 16.55% (fruit) [48]; 0.3–7.6% (leaf) [52] | <3% (fruit) [47]; 1.53% (fruit) [48] | <1% (fruit) [47]; 3.91% (fruit) [48]; 0.2–1.6% (leaf) [52] |
| Japanese star anise | 6.6% [53] | 3.5% [53] | 9.8% [53] | |
| Tarragon | 9.59–28.40% (seeds) [54] | 21.45–38.90% (seeds) [54] | ||
| Sweet bay | 3.1% (flower) [55]; 4.7% (bark) [55]; 16.0% (stem) [55]; 11.8–21.3% (leaf) [55,56] | 0.8% (stem) [55]; 5% (leaf) [56] |
2.2. Myristicin and Elemicin in Food Flavorings
2.3. Myristicin and Elemicin in Foods
3. Toxicity of Myristicin and Elemicin: Lessons Learned from Safrole and Methyleugenol
3.1. Metabolism of Myristicin and Elemicin vs. Safrole and Methyleugenol
3.1.1. Common Structural Features
3.1.2. Metabolism of Methyleugenol
3.1.3. Metabolism of Elemicin
3.1.4. Metabolism of Safrole
3.1.5. Metabolism of Myristicin
3.2. Genotoxicity
3.2.1. Genotoxicity of Methyleugenol vs. Elemicin
3.2.2. Genotoxicity of Safrole vs. Myristicin
3.2.3. Genotoxic Effects in Pregnant Mice and in Offsprings
3.3. Carcinogenicity of Safrole, Methyleugenol, Myristicin and Elemicin
3.4. Other Toxicological Endpoints
3.4.1. Acute Toxicity of Methyleugenol vs. Elemicin and Safrole vs. Myristicin
3.4.2. Subchronic Toxic Effects of Methyleugenol vs. Elemicin and Safrole vs. Myristicin
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IARC. Overall evaluations of carcinogenicity: An updating of IARC Monographs Volumes 1 to 42—Eugenol. IARC Monogr. Eval. Carcinog. Risk Chem. Hum. 1987, 36, 1–63. [Google Scholar]
- IARC. Some chemicals present in industrial and consumer products, food and drinking-water/Methyleugenol. IARC Monogr. Eval. Carcinog. Risks Hum. 2013, 101, 407–433. [Google Scholar]
- SCF. Opinion of the Scientific Committee on Food on Methyleugenol (4-Allyl-1,2-dimethoxybenzene). In SCF/CS/FLAV/FLAVOUR/4 ADD1 FINAL; European Commission: Brussels, Belgium, 2001; pp. 1–10. [Google Scholar]
- SCF. Opinion of the Scientific Committee on Food on the safety of the presence of safrole (1-allyl-3,4-methylene dioxy benzene) in flavourings and other food ingredients with flavouring properties. In SCF/CS/FLAV/FLAVOUR/6 ADD3 FINAL; European Commission: Brussels, Belgium, 2002; pp. 1–15. [Google Scholar]
- Eisenreich, A.; Götz, M.E.; Sachse, B.; Monien, B.H.; Herrmann, K.; Schäfer, B. Alkenylbenzenes in Foods: Aspects Impeding the Evaluation of Adverse Health Effects. Foods 2021, 10, 2139. [Google Scholar] [CrossRef] [PubMed]
- Solheim, E.; Scheline, R.R. Metabolism of alkenebenzene derivatives in the rat III. Elemicin and isoelemicin. Xenobiotica 1980, 10, 371–380. [Google Scholar] [CrossRef]
- Lee, H.S.; Jeong, T.C.; Kim, J.H. In vitro and in vivo metabolism of myristicin in the rat. J. Chromatogr. B Biomed. Sci. Appl. 1998, 705, 367–372. [Google Scholar] [CrossRef]
- Beyer, J.; Ehlers, D.; Maurer, H.H. Abuse of nutmeg (Myristica fragrans Houtt.): Studies on the metabolism and the toxicologic detection of its ingredients elemicin, myristicin, and safrole in rat and human urine using gas chromatography/mass spectrometry. Ther. Drug Monit. 2006, 28, 568–575. [Google Scholar] [CrossRef]
- Kristanc, L.; Kreft, S. European medicinal and edible plants associated with subacute and chronic toxicity part I: Plants with carcinogenic, teratogenic and endocrine-disrupting effects. Food Chem. Toxicol. 2016, 92, 150–164. [Google Scholar] [CrossRef]
- Martins, C.; Rueff, J.; Rodrigues, A.S. Genotoxic alkenylbenzene flavourings, a contribution to risk assessment. Food Chem. Toxicol. 2018, 118, 861–879. [Google Scholar] [CrossRef]
- Al-Malahmeh, A.J.; Alajlouni, A.M.; Ning, J.; Wesseling, S.; Vervoort, J.; Rietjens, I. Determination and risk assessment of naturally occurring genotoxic and carcinogenic alkenylbenzenes in nutmeg-based plant food supplements. J. Appl. Toxicol. 2017, 37, 1254–1264. [Google Scholar] [CrossRef]
- Raffo, A.; D’Aloise, A.; Magri, A.L.; Leclercq, C. Quantitation of tr-cinnamaldehyde, safrole and myristicin in cola-flavoured soft drinks to improve the assessment of their dietary exposure. Food Chem. Toxicol. 2013, 59, 626–635. [Google Scholar] [CrossRef]
- Siano, F.; Ghizzoni, C.; Gionfriddo, F.; Colombo, E.; Servillo, L.; Castaldo, D. Determination of estragole, safrole and eugenol methyl ether in food products. Food Chem. 2003, 81, 469–475. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Yoo, J.H.; Yu, C.Y.; Chung, I.M. GC-MS analysis of volatile compounds of Perilla frutescens Britton var. Japonica accessions: Morphological and seasonal variability. Asian Pac. J. Trop. Med. 2017, 10, 643–651. [Google Scholar] [CrossRef]
- Muráriková, A.; Ťažký, A.; Neugebauerová, J.; Planková, A.; Jampílek, J.; Mučaji, P.; Mikuš, P. Characterization of essential oil composition in different basil species and pot cultures by a GC-MS method. Molecules 2017, 22, 1221. [Google Scholar] [CrossRef] [Green Version]
- Simándi, B.; Deák, A.; Rónyai, E.; Yanxiang, G.; Veress, T.; Lemberkovics, É.; Then, M.; Sass-Kiss, Á.; Vámos-Falusi, Z. Supercritical carbon dioxide extraction and fractionation of fennel oil. J. Agric. Food Chem. 1999, 47, 1635–1640. [Google Scholar] [CrossRef]
- Shulgin, A.T. Possible implication of myristicin as a psychotropic substance. Nature 1966, 210, 380–384. [Google Scholar] [CrossRef]
- Shulgin, A.T. Composition of the myristicin fraction from oil of nutmeg. Nature 1963, 197, 379. [Google Scholar] [CrossRef]
- Du, S.S.; Yang, K.; Wang, C.F.; You, C.X.; Geng, Z.F.; Guo, S.S.; Deng, Z.W.; Liu, Z.L. Chemical constituents and activities of the essential oil from myristica fragrans against cigarette beetle lasioderma serricorne. Chem. Biodivers. 2014, 11, 1449–1456. [Google Scholar] [CrossRef]
- Analytical Methods Committee. Application of gas-liquid chromatography to the analysis of essential oils. Part XIV. Monographs for five essential oils. Analyst 1988, 113, 1125–1136. [Google Scholar] [CrossRef]
- Sanford, K.J.; Heinz, D. Effects of storage on the volatile composition of nutmeg. Phytochemistry 1971, 10, 1245–1250. [Google Scholar] [CrossRef]
- Baldry, J.; Dougan, J.; Matthews, W.S.; Nabney, J.; Pickering, G.R.; Robinson, F.V. Composition and Flavour of Nutmeg oils. Int. Flavours Food Addit. 1976, 7, 28–30. [Google Scholar]
- Archer, A.W. Determination of safrole and myristicin in nutmeg and mace by high-performance liquid chromatography. J. Chromatogr. A 1988, 438, 117–121. [Google Scholar] [CrossRef]
- Hallstrom, H.; Thuvander, A. Toxicological evaluation of myristicin. Nat. Toxins 1997, 5, 186–192. [Google Scholar] [CrossRef]
- Lopez, P.; van Sisseren, M.; De Marco, S.; Jekel, A.; de Nijs, M.; Mol, H.G. A straightforward method to determine flavouring substances in food by GC-MS. Food Chem. 2015, 174, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Ávila, M.; Zougagh, M.; Escarpa, A.; Rios, A. Determination of alkenylbenzenes and related flavour compounds in food samples by on-column preconcentration-capillary liquid chromatography. J. Chromatogr. A 2009, 1216, 7179–7185. [Google Scholar] [CrossRef] [PubMed]
- Franz, C.; Glasl, H. Zur Kenntnis der Ätherischen Öle von Petersilie II. Vergleichende Untersuchung des Frucht-, Blatt- und Wurzelöles einiger Petersiliensorten. Qual. Plant. 1976, 25, 253–262. [Google Scholar] [CrossRef]
- Simon, J.E.; Quinn, J. Characterization of essential oil of parsley. J. Agric. Food Chem. 1988, 36, 467–472. [Google Scholar] [CrossRef]
- Macleod, A.J.; Snyder, C.H.; Subramanian, G. Volatile aroma constituents of parsley leaves. Phytochemistry 1985, 24, 2623–2627. [Google Scholar] [CrossRef]
- Masanetz, C.; Grosch, W. Key odorants of parsley leaves (Petroselinum crispum [Mill.] Nym. ssp. crispum) by Odour–activity values. Flavour. Frag. J. 1998, 13, 115–124. [Google Scholar] [CrossRef]
- Trenkle, K. Recent studies on fennel (Foeniculum vulgare M.) 2. The volatile oil of the fruit, herbs and roots of fruit-bearing plants. Die Pharm. 1972, 27, 319–324. [Google Scholar]
- Hao, Y.; Kang, J.; Guo, X.; Yang, R.; Chen, Y.; Li, J.; Shi, L. Comparison of Nutritional Compositions and Essential Oil Profiles of Different Parts of a Dill and Two Fennel Cultivars. Foods 2021, 10, 1784. [Google Scholar] [CrossRef]
- Al-Sheddi, E.S.; Al-Zaid, N.A.; Al-Oqail, M.M.; Al-Massarani, S.M.; El-Gamal, A.A.; Farshori, N.N. Evaluation of cytotoxicity, cell cycle arrest and apoptosis induced by Anethum graveolens L. essential oil in human hepatocellular carcinoma cell line. Saudi Pharm. J. 2019, 27, 1053–1060. [Google Scholar] [CrossRef]
- Kubeczka, K.H.; Stahl, E. [On the essential oils from the Apiaceae (Umbelliferae). I. The oil of roots from Pastinaca sativa (author’s transl)]. Planta Med. 1975, 27, 235–241. [Google Scholar] [CrossRef]
- Stahl, E. Variation of Myristicin Content in Cultivated Parsnip Roots (Pastinaca sativa ssp. sativa var. hortensis). J. Agric. Food Chem. 1981, 29, 890–892. [Google Scholar] [CrossRef]
- Lichtenstein, E.P.; Casida, J.E. Naturally occurring insecticides: Myristicin, an Insecticide and Synergist Occurring Naturally in the Edible Parts of Parsnips. J. Agric. Food Chem. 1963, 11, 410–415. [Google Scholar] [CrossRef]
- Fischer, R.; Nitzan, N.; Chaimovitsh, D.; Rubin, B.; Dudai, N. Variation in essential oil composition within individual leaves of sweet basil (Ocimum basilicum L.) is more affected by leaf position than by leaf age. J. Agric. Food Chem. 2011, 59, 4913–4922. [Google Scholar] [CrossRef]
- Chalchat, J.-C.; Özcan, M.M. Comparative essential oil composition of flowers, leaves and stems of basil (Ocimum basilicum L.) used as herb. Food Chem. 2008, 110, 501–503. [Google Scholar] [CrossRef]
- Vani, S.R.; Cheng, S.; Chuah, C. Comparative study of volatile compounds from genus Ocimum. Am. J. Appl. Sci. 2009, 6, 523. [Google Scholar] [CrossRef]
- Smaili, T.; Zellagui, A.; Cioni, P.L.; Flamini, G. A myristicin-rich essential oil from Daucus sahariensis growing in Algeria. Nat. Prod. Commun. 2011, 6, 883–886. [Google Scholar] [CrossRef] [Green Version]
- Jabrane, A.; Jannet, H.B.; Harzallah-Skhiri, F.; Mastouri, M.; Casanova, J.; Mighri, Z. Flower and root oils of the tunisian Daucus carota L. ssp.maritimus (Apiaceae): Integrated analyses by GC, GC/MS, and 13C-NMR spectroscopy, and in vitro antibacterial activity. Chem. Biodivers. 2009, 6, 881–889. [Google Scholar] [CrossRef]
- Buttery, R.G.; Seifert, R.M.; Guadagni, D.G.; Black, D.R.; Ling, L. Characterization of some volatile constituents of carrots. J. Agric. Food Chem. 1968, 16, 1009–1015. [Google Scholar] [CrossRef]
- Wulf, L.W.; Nagel, C.W.; Branen, A.L. Analysis of Myristicin and Falcarinol in Carrots by High-Pressure Liquid Chromatography. J. Agric. Food Chem. 1978, 26, 1390–1393. [Google Scholar] [CrossRef]
- Mansour, E.S.S.; Maatooq, G.T.; Khalil, A.T.; Marwan, E.S.M.; Sallam, A.A. Essential oil of Daucus glaber Forssk. Z. Naturforsch. Sect. C J. Biosci. 2004, 59, 373–378. [Google Scholar] [CrossRef]
- Marzouki, H.; Khaldi, A.; Falconieri, D.; Piras, A.; Marongiu, B.; Molicotti, P.; Zanetti, S. Essential oils of Daucus carota subsp. carota of Tunisia obtained by supercritical carbon dioxide extraction. Nat. Prod. Commun. 2010, 5, 1955–1958. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Pérez, A.; López-Ruiz, R.; Romero-González, R.; Frenich, A.G. A new strategy based on gas chromatography–high resolution mass spectrometry (GC–HRMS-Q-Orbitrap) for the determination of alkenylbenzenes in pepper and its varieties. Food Chem. 2020, 321, 126727. [Google Scholar] [CrossRef]
- Jirovetz, L.; Buchbauer, G.; Ngassoum, M.B.; Geissler, M. Aroma compound analysis of Piper nigrum and Piper guineense essential oils from Cameroon using solid-phase microextraction–gas chromatography, solid-phase microextraction–gas chromatography–mass spectrometry and olfactometry. J. Chromatogr. A 2002, 976, 265–275. [Google Scholar] [CrossRef]
- Ekundayo, O.; Laakso, I.; Hiltunen, R.; Adegbola, R.M.; Oguntimein, B.; Sofowora, A. Essential oil constituents of ashanti pepper (piper guineense) fruits (Berries). J. Agric. Food Chem. 1988, 36, 880–882. [Google Scholar] [CrossRef]
- Bessiere, J.; Menut, C.; Lamaty, G.; Joseph, H. Variations in the volatile constituents of peperomia rotundifolia schlecht. & cham. grown on different host-trees in guadeloupe. Flavour. Frag. J. 1994, 9, 131–133. [Google Scholar]
- De Diaz, A.M.; Diaz, P.P.; Cardoso, H. Volatile Constituents of Peperomia subespatulata. Planta Med. 1988, 54, 92–93. [Google Scholar] [CrossRef]
- Andrade, E.H.A.; Carreira, L.M.M.; da Silva, M.H.L.; da Silva, J.D.; Bastos, C.N.; Sousa, P.J.C.; Guimarães, E.F.; Maia, J.G.S. Variability in Essential-Oil Composition of Piper marginatum sensu lato. Chem. Biodivers. 2008, 5, 197–208. [Google Scholar] [CrossRef]
- De Lira, P.N.B.; Da Silva, J.K.R.; Andrade, E.H.A.; Sousa, P.J.C.; Silva, N.N.S.; Maia, J.G.S. Essential oil composition of three Peperomia species from the Amazon, Brazil. Nat. Pro. Comm. 2009, 4, 427–430. [Google Scholar] [CrossRef] [Green Version]
- Cook, W.; Howard, A. The essential oil of Illicium anisatum Linn. Can. J. Chem. 1966, 44, 2461–2464. [Google Scholar] [CrossRef]
- Lawrence, B.M. Progress in essential oils/Progrès dans le domaine des huiles essentielles. Perfum. Flavor. 1988, 13, 44–50. [Google Scholar]
- Fiorini, C.; Fouraste, I.; David, B.; Bessiere, J. Composition of the Flower, Leaf and Stem Essential Oils from Laurus nobilis. Flavour. Frag. J. 1997, 12, 91–93. [Google Scholar] [CrossRef]
- Conforti, F.; Statti, G.; Uzunov, D.; Menichini, F. Comparative chemical composition and antioxidant activities of wild and cultivated Laurus nobilis L. leaves and Foeniculum vulgare subsp. piperitum (Ucria) coutinho seeds. Biol. Pharm. Bull. 2006, 29, 2056–2064. [Google Scholar] [CrossRef] [Green Version]
- Krishnamoorty, B.; Rema, J. Nutmeg and mace. In Handbook of Herbs and Spices; Peter, K.V., Ed.; CRC Press-Woodhead Publishing Ltd.: Boca Raton, FL, USA, 2001; Volume 1, pp. 238–248. [Google Scholar]
- Al-Malahmeh, A.J.; Alajlouni, A.M.; Wesseling, S.; Vervoort, J.; Rietjens, I.M.C.M. Determination and risk assessment of naturally occurring genotoxic and carcinogenic alkenylbenzenes in basil-containing sauce of pesto. Toxicol. Rep. 2017, 4, 1–8. [Google Scholar] [CrossRef]
- Alajlouni, A.M.; Al-Malahmeh, A.J.; Isnaeni, F.N.; Wesseling, S.; Vervoort, J.; Rietjens, I.M.C.M. Level of Alkenylbenzenes in Parsley and Dill Based Teas and Associated Risk Assessment Using the Margin of Exposure Approach. J. Agric. Food Chem. 2016, 64, 8640–8646. [Google Scholar] [CrossRef]
- Alajlouni, A.M.; Al-Malahmeh, A.J.; Wesseling, S.; Kalli, M.; Vervoort, J.; Rietjens, I.M.C.M. Risk assessment of combined exposure to alkenylbenzenes through consumption of plant food supplements containing parsley and dill. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2017, 34, 2201–2211. [Google Scholar] [CrossRef]
- De Vincenzi, M.; De Vincenzi, A.; Silano, M. Constituents of aromatic plants: Elemicin. Fitoterapia 2004, 75, 615–618. [Google Scholar] [CrossRef]
- De Vincenzi, M.; Silano, M.; Maialetti, F.; Scazzocchio, B. Constituents of aromatic plants: II. Estragole. Fitoterapia 2000, 71, 725–729. [Google Scholar] [CrossRef]
- De Vincenzi, M.; Silano, M.; Stacchini, P.; Scazzocchio, B. Constituents of aromatic plants: I. Methyleugenol. Fitoterapia 2000, 71, 216–221. [Google Scholar] [CrossRef]
- Dawidowicz, A.L.; Dybowski, M.P. Determination of myristicin in commonly spices applying SPE/GC. Food Chem. Toxicol. 2012, 50, 2362–2367. [Google Scholar] [CrossRef]
- Dawidowicz, A.L.; Dybowski, M.P. Simple and rapid determination of myristicin in human serum. Forensic Toxicol. 2013, 31, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Xie, Q.; Zhang, S.; Xu, H.; Su, J.; Zhang, J.; Deng, C.; Song, G. Fabrication of functionalized magnetic microspheres based on monodispersed polystyrene for quantitation of allyl-benzodioxoles coupled with gas chromatography and mass spectrometry. J. Chromatogr. A 2019, 1607, 460402. [Google Scholar] [CrossRef]
- Guenthner, T.M.; Luo, G. Investigation of the role of the 2′,3′-epoxidation pathway in the bioactivation and genotoxicity of dietary allylbenzene analogs. Toxicology 2001, 160, 47–58. [Google Scholar] [CrossRef]
- Solheim, E.; Scheline, R.R. Metabolism of alkenebenzene derivatives in the rat I. P-methoxyallylbenzene (estragole) and p-methoxypropenylbenzene (anethole). Xenobiotica 1973, 3, 493–510. [Google Scholar] [CrossRef]
- Solheim, E.; Scheline, R.R. Metabolism of alkenebenzene derivatives in the rat. II. Eugenol and isoeugenol methyl ethers. Xenobiotica 1976, 6, 137–150. [Google Scholar] [CrossRef]
- Stillwell, W.; Carman, J.K.; Bell, L.; Horning, M.G. The metabolism of safrole and 2′,3′-epoxysafrole in the rat and guinea pig. Drug Metab. Dispos. 1974, 2, 489–498. [Google Scholar]
- Wislocki, P.; Borchert, P.; Miller, J.; Miller, E. The metabolic activation of the carcinogen 1′-hydroxysafrole in vivo and in vitro and the electrophilic reactivities of possible ultimate carcinogens. Cancer Res. 1976, 36, 1686–1695. [Google Scholar]
- Delaforge, M.; Janiaud, P.; Levi, P.; Morizot, J.P. Biotransformation of allylbenzene analogues in vivo and in vitro through the epoxide-diol pathway. Xenobiotica 1980, 10, 737–744. [Google Scholar] [CrossRef]
- Fischer, I.U.; Von Unruh, G.E.; Dengler, H.J. The metabolism of eugenol in man. Xenobiotica 1990, 20, 209–222. [Google Scholar] [CrossRef]
- Boberg, E.W.; Miller, E.C.; Miller, J.A. The metabolic sulfonation and side-chain oxidation of 3′-hydroxyisosafrole in the mouse and its inactivity as a hepatocarcinogen relative to 1′-hydroxysafrole. Chem. Biol. Interact. 1986, 59, 73–97. [Google Scholar] [CrossRef]
- Boberg, E.W.; Miller, E.C.; Miller, J.A.; Poland, A.; Liem, A. Strong evidence from studies with brachymorphic mice and pentachlorophenol that 1′-sulfoöxysafrole is the major ultimate electrophilic and carcinogenic metabolite of 1′-hydroxysafrole in mouse liver. Cancer Res. 1983, 43, 5163–5173. [Google Scholar]
- Van den Berg, S.J.P.L.; Punt, A.; Soffers, A.E.; Vervoort, J.; Ngeleja, S.; Spenkelink, B.; Rietjens, I.M.C.M. Physiologically based kinetic models for the alkenylbenzene elemicin in rat and human and possible implications for risk assessment. Chem. Res. Toxicol. 2012, 25, 2352–2367. [Google Scholar] [CrossRef]
- Monien, B.H.; Sachse, B.; Niederwieser, B.; Abraham, K. Detection of N-Acetyl-S-[3′-(4-methoxyphenyl)allyl]-l-Cys (AMPAC) in Human Urine Samples after Controlled Exposure to Fennel Tea: A New Metabolite of Estragole and trans-Anethole. Chem. Res. Toxicol. 2019, 32, 2260–2267. [Google Scholar] [CrossRef]
- Herrmann, K.; Engst, W.; Appel, K.E.; Monien, B.H.; Glatt, H. Identification of human and murine sulfotransferases able to activate hydroxylated metabolites of methyleugenol to mutagens in Salmonella typhimurium and detection of associated DNA adducts using UPLC-MS/MS methods. Mutagenesis 2012, 27, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Klungsøyr, J.; Scheline, R.R. Metabolism of Safrole in the Rat. Acta Pharmacol. Toxicol. 1983, 52, 211–216. [Google Scholar] [CrossRef]
- Anders, M.W.; Sunram, J.M.; Wilkinson, C.F. Mechanism of the metabolism of 1,3-benzodioxoles to carbon monoxide. Biochem Pharm. 1984, 33, 577–580. [Google Scholar] [CrossRef]
- Martati, E.; Boersma, M.G.; Spenkelink, A.; Khadka, D.B.; van Bladeren, P.J.; Rietjens, I.M.; Punt, A. Physiologically based biokinetic (PBBK) modeling of safrole bioactivation and detoxification in humans as compared with rats. Toxicol. Sci. 2012, 128, 301–316. [Google Scholar] [CrossRef] [Green Version]
- Bolton, J.L.; Acay, N.M.; Vukomanovic, V. Evidence That 4-Allyl-o-quinones Spontaneously Rearrange to Their More Electrophilic Quinone Methides: Potential Bioactivation Mechanism for the Hepatocarcinogen Safrole. Chem. Res. Toxicol. 1994, 7, 443–450. [Google Scholar] [CrossRef]
- Dietz, B.M.; Bolton, J.L. Biological reactive intermediates (BRIs) formed from botanical dietary supplements. Chem. Biol. Interact. 2011, 192, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Bolton, J.L.; Dunlap, T.L.; Dietz, B.M. Formation and biological targets of botanical o-quinones. Food Chem. Toxicol. 2018, 120, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Tsai, R.S.; Carrupt, P.A.; Testa, B.; Caldwell, J. Structure-genotoxicity relationships of allylbenzenes and propenylbenzenes: A quantum chemical study. Chem. Res. Toxicol. 1994, 7, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.; Constantin-Teodosiu, D.; Egestad, B.; Mickos, H.; Moldeus, P. Formation of glutathione conjugates during oxidation of eugenol by microsomal fractions of rat liver and lung. Biochem. Pharm. 1990, 39, 1587–1595. [Google Scholar] [CrossRef]
- Thompson, D.C.; Constantin-Teodosiu, D.; Moldeus, P. Metabolism and cytotoxicity of eugenol in isolated rat hepatocytes. Chem. Biol. Interact. 1991, 77, 137–147. [Google Scholar] [CrossRef]
- Wang, Y.K.; Yang, X.N.; Zhu, X.; Xiao, X.R.; Yang, X.W.; Qin, H.B.; Gonzalez, F.J.; Li, F. Role of Metabolic Activation in Elemicin-Induced Cellular Toxicity. J. Agric. Food Chem. 2019, 67, 8243–8252. [Google Scholar] [CrossRef] [PubMed]
- Jeurissen, S.M.F.; Bogaards, J.J.P.; Awad, H.M.; Boersma, M.G.; Brand, W.; Fiamegos, Y.C.; Van Beek, T.A.; Alink, G.M.; Sudhölter, E.J.R.; Cnubben, N.H.P.; et al. Human cytochrome P450 enzyme specificity for bioactivation of safrole to the proximate carcinogen 1′-hydroxysafrole. Chem. Res. Toxicol. 2004, 17, 1245–1250. [Google Scholar] [CrossRef] [PubMed]
- Jeurissen, S.M.F.; Bogaards, J.J.P.; Boersma, M.G.; Ter Horst, J.P.F.; Awad, H.M.; Fiamegos, Y.C.; Van Beek, T.A.; Alink, G.M.; Sudhölter, E.J.R.; Cnubben, N.H.P.; et al. Human cytochrome P450 enzymes of importance for the bioactivation of methyleugenol to the proximate carcinogen 1′-hydroxymethyleugenol. Chem. Res. Toxicol. 2006, 19, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Jeurissen, S.M.F.; Punt, A.; Boersma, M.G.; Bogaards, J.J.P.; Fiamegos, Y.C.; Schilter, B.; Van Bladeren, P.J.; Cnubben, N.H.P.; Rietjens, I.M.C.M. Human cytochrome P450 enzyme specificity for the bioactivation of estragole and related alkenylbenzenes. Chem. Res. Toxicol. 2007, 20, 798–806. [Google Scholar] [CrossRef]
- NTP. NTP Technical Report on the Toxicology and Carcinogenesis Studies of Methyleugenol (CAS NO. 93-15-2) in F344/N Rats and B6C3F1 Mice (Gavage Studies). Natl. Toxicol. Program Tech. Rep. Ser. 2000, 491, 1–412. [Google Scholar]
- Yun, C.H.; Lee, H.S.; Lee, H.Y.; Yim, S.K.; Kim, K.H.; Kim, E.; Yea, S.S.; Guengerich, F.P. Roles of human liver cytochrome P450 3A4 and 1A2 enzymes in the oxidation of myristicin. Toxicol. Lett. 2003, 137, 143–150. [Google Scholar] [CrossRef]
- Tremmel, R.; Herrmann, K.; Engst, W.; Meinl, W.; Klein, K.; Glatt, H.; Zanger, U.M. Methyleugenol DNA adducts in human liver are associated with SULT1A1 copy number variations and expression levels. Arch. Toxicol. 2017, 91, 3329–3339. [Google Scholar] [CrossRef]
- Zangouras, A.; Caldwell, J.; Hutt, A.J.; Smith, R.L. Dose dependent conversion of estragole in the rat and mouse to the carcinogenic metabolite, 1 -hydroxyestragole. Biochem. Pharmacol. 1981, 30, 1383–1386. [Google Scholar] [CrossRef]
- Yao, H.; Peng, Y.; Zheng, J. Identification of glutathione and related cysteine conjugates derived from reactive metabolites of methyleugenol in rats. Chem. Biol. Interact. 2016, 253, 143–152. [Google Scholar] [CrossRef]
- Gardner, I.; Wakazono, H.; Bergin, P.; De Waziers, I.; Beaune, P.; Kenna, J.G.; Caldwell, J. Cytochrome P450 mediated bioactivation of methyleugenol to 1′-hydroxymethyleugenol in Fischer 344 rat and human liver microsomes. Carcinogenesis 1997, 18, 1775–1783. [Google Scholar] [CrossRef] [Green Version]
- Casida, J.E.; Engel, J.L.; Essac, E.G.; Kamienski, F.X.; Kuwatsuka, S. Methylene-C14-dioxyphenyl compounds: Metabolism in relation to their synergistic action. Science 1966, 153, 1130–1133. [Google Scholar] [CrossRef]
- Kamienski, F.X.; Casida, J.E. Importance of demethylenation in the metabolism in vivo and in vitro of methylenedioxyphenyl synergists and related compounds in mammals. Biochem. Pharmacol. 1970, 19, 91–112. [Google Scholar] [CrossRef]
- Randerath, K.; Haglund, R.E.; Phillips, D.H.; Reddy, M.V. 32P-post-labelling analysis of DNA adducts formed in the livers of animals treated with safrole, estragole and other naturally-occurring alkenylbenzenes. I. adult female CD-1 mice. Carcinogenesis 1984, 5, 1613–1622. [Google Scholar] [CrossRef]
- Randerath, K.; Putman, K.L.; Randerath, E. Flavor constituents in cola drinks induce hepatic DNA adducts in adult and fetal mice. Biochem. Biophys. Res. Commun. 1993, 192, 61–68. [Google Scholar] [CrossRef]
- Randerath, K.; Randerath, E.; Danna, T.F.; van Golen, L.; Putman, K.L. A new sensitive 32P-postlabeling assay based on the specific enzymatic conversion of bulky DNA lesions to radiolabeled dinucleotides and nucleoside 5′-monophosphates. Carcinogenesis 1989, 10, 1231–1239. [Google Scholar] [CrossRef]
- Randerath, K.; Reddy, M.V.; Gupta, R.C. 32P-labeling test for DNA damage. Proc. Natl. Acad. Sci. USA 1981, 78, 6126–6129. [Google Scholar] [CrossRef] [Green Version]
- Reddy, M.V.; Randerath, K. Nuclease p1-mediated enhancement of sensitivity of 32P-postlabeling test for structurally diverse DNA adducts. Carcinogenesis 1986, 7, 1543–1551. [Google Scholar] [CrossRef]
- Bergau, N.; Herfurth, U.M.; Sachse, B.; Abraham, K.; Monien, B.H. Bioactivation of estragole and anethole leads to common adducts in DNA and hemoglobin. Food Chem. Toxicol. 2021, 153, 112253. [Google Scholar] [CrossRef]
- Phillips, D.H.; Miller, J.A.; Miller, E.C.; Adams, B. Structures of the DNA adducts formed in mouse liver after administration of the proximate hepatocarcinogen 1′-hydroxyestragole. Cancer Res. 1981, 41, 176–186. [Google Scholar]
- Phillips, D.H.; Miller, J.A.; Miller, E.C.; Adams, B. N2 atom of guanine and N6 atom of adenine residues as sites for covalent binding of metabolically activated 1′-hydroxysafrole to mouse liver DNA in vivo. Cancer Res. 1981, 41, 2664–2671. [Google Scholar]
- Rietjens, I.M.; Cohen, S.M.; Fukushima, S.; Gooderham, N.J.; Hecht, S.; Marnett, L.J.; Smith, R.L.; Adams, T.B.; Bastaki, M.; Harman, C.G.; et al. Impact of structural and metabolic variations on the toxicity and carcinogenicity of hydroxy- and alkoxy-substituted allyl- and propenylbenzenes. Chem. Res. Toxicol. 2014, 27, 1092–1103. [Google Scholar] [CrossRef] [Green Version]
- Groh, I.A.M.; Cartus, A.T.; Vallicotti, S.; Kajzar, J.; Merz, K.H.; Schrenk, D.; Esselen, M. Genotoxic potential of methyleugenol and selected methyleugenol metabolites in cultured Chinese hamster V79 cells. Food Funct. 2012, 3, 428–436. [Google Scholar] [CrossRef]
- Groh, I.A.M.; Rudakovski, O.; Gründken, M.; Schroeter, A.; Marko, D.; Esselen, M. Methyleugenol and oxidative metabolites induce DNA damage and interact with human topoisomerases. Arch. Toxicol. 2016, 90, 2809–2823. [Google Scholar] [CrossRef]
- Groh, I.A.M.; Esselen, M. Methyleugenol and selected oxidative metabolites affect DNA-Damage signalling pathways and induce apoptosis in human colon tumour HT29 cells. Food Chem. Toxicol. 2017, 108, 267–275. [Google Scholar] [CrossRef]
- Chan, V.S.; Caldwell, J. Comparative induction of unscheduled DNA synthesis in cultured rat hepatocytes by allylbenzenes and their 1′-hydroxy metabolites. Food Chem. Toxicol. 1992, 30, 831–836. [Google Scholar] [CrossRef]
- Ellis, J.K.; Carmichael, P.L.; Gooderham, N.J. DNA adduct levels in the liver of the F344 rat treated with the natural flavour methyleugenol. Toxicology 2006, 1, 73–74. [Google Scholar] [CrossRef]
- Herrmann, K.; Schumacher, F.; Engst, W.; Appel, K.E.; Klein, K.; Zanger, U.M.; Glatt, H. Abundance of DNA adducts of methyleugenol, a rodent hepatocarcinogen, in human liver samples. Carcinogenesis 2013, 34, 1025–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, K.; Engst, W.; Florian, S.; Lampen, A.; Meinl, W.; Glatt, H.R. The influence of the SULT1A status-wild-type, knockout or humanized-on the DNA adduct formation by methyleugenol in extrahepatic tissues of mice. Toxicol. Res. 2016, 5, 808–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, K.; Engst, W.; Meinl, W.; Florian, S.; Cartus, A.T.; Schrenk, D.; Appel, K.E.; Nolden, T.; Himmelbauer, H.; Glatt, H. Formation of hepatic DNA adducts by methyleugenol in mouse models: Drastic decrease by Sult1a1 knockout and strong increase by transgenic human SULT1A1/2. Carcinogenesis 2014, 35, 935–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Wesseling, S.; Rietjens, I. Estragole DNA adduct accumulation in human liver HepaRG cells upon repeated in vitro exposure. Toxicol. Lett. 2021, 337, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Feng, Y.; Zhang, Z.; Wang, H.; Li, W.; Wang, D.O.; Peng, Y.; Zheng, J. In Vitro and In Vivo Evidence for RNA Adduction Resulting from Metabolic Activation of Methyleugenol. J. Agric. Food Chem. 2020, 68, 15134–15141. [Google Scholar] [CrossRef]
- Van den Berg, S.; Restani, P.; Boersma, M.; Delmulle, L.; Rietjens, I. Levels of Genotoxic and Carcinogenic Compounds in Plant Food Supplements and Associated Risk Assessment. Food Nutr. Sci. 2011, 2, 989–1010. [Google Scholar] [CrossRef] [Green Version]
- Hasheminejad, G.; Caldwell, J. Genotoxicity of the alkenylbenzenes α- and β-asarone, myristicin and elemicin as determined by the UDS assay in cultured rat hepatocytes. Food Chem. Toxicol. 1994, 32, 223–231. [Google Scholar] [CrossRef]
- Sekizawa, J.; Shibamoto, T. Genotoxicity of safrole-related chemicals in microbial test systems. Mutat. Res. 1982, 101, 127–140. [Google Scholar] [CrossRef]
- NTP. Methyleugenol CAS No. 93-15-2. Report on Carcinogens. 2016; pp. 1–2. Available online: https://ntp.niehs.nih.gov/ntp/roc/content/profiles/methyleugenol.pdf (accessed on 13 May 2022).
- Devereux, T.R.; Anna, C.H.; Foley, J.F.; White, C.M.; Sills, R.C.; Barrett, J.C. Mutation of beta-catenin is an early event in chemically induced mouse hepatocellular carcinogenesis. Oncogene 1999, 18, 4726–4733. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Kijima, A.; Hibi, D.; Ishii, Y.; Takasu, S.; Matsushita, K.; Kuroda, K.; Nohmi, T.; Nishikawa, A.; Umemura, T. In vivo genotoxicity of methyleugenol in gpt delta transgenic rats following medium-term exposure. Toxicol. Sci. 2013, 131, 387–394. [Google Scholar] [CrossRef]
- Daimon, H.; Sawada, S.; Asakura, S.; Sagami, F. Inhibition of sulfotransferase affecting in vivo genotoxicity and DNA adducts induced by safrole in rat liver. Teratog. Carcinog. Mutag. 1997, 17, 327–337. [Google Scholar] [CrossRef]
- Wiseman, R.W.; Miller, E.C.; Miller, J.A.; Liem, A. Structure-Activity Studies of the Hepatocarcinogenicities of Alkenylbenzene Derivatives Related to Estragole and Safrole on Administration to Preweanling Male C57BL/6JΧC3H/HeJ F1Mice. Cancer Res. 1987, 47, 2275–2283. [Google Scholar] [CrossRef]
- Daimon, H.; Sawada, S.; Asakura, S.; Sagami, F. In vivo genotoxicity and DNA adduct levels in the liver of rats treated with safrole. Carcinogenesis 1998, 19, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Howes, A.J.; Chan, V.S.; Caldwell, J. Structure-specificity of the genotoxicity of some naturally occurring alkenylbenzenes determined by the unscheduled DNA synthesis assay in rat hepatocytes. Food Chem. Toxicol. 1990, 28, 537–542. [Google Scholar] [CrossRef]
- Martin, C.N.; McDermid, A.C.; Garner, R.C. Testing of known carcinogens and noncarcinogens for their ability to induce unscheduled DNA synthesis in HeLa cells. Cancer Res. 1978, 38, 2621–2627. [Google Scholar]
- Zhou, G.D.; Moorthy, B.; Bi, J.; Donnelly, K.C.; Randerath, K. DNA adducts from alkoxyallylbenzene herb and spice constituents in cultured human (HepG2) cells. Environ. Mol. Mutagen. 2007, 48, 715–721. [Google Scholar] [CrossRef]
- Kobets, T.; Cartus, A.T.; Fuhlbrueck, J.A.; Brengel, A.; Stegmuller, S.; Duan, J.D.; Brunnemann, K.D.; Williams, G.M. Assessment and characterization of DNA adducts produced by alkenylbenzenes in fetal turkey and chicken livers. Food Chem. Toxicol. 2019, 129, 424–433. [Google Scholar] [CrossRef]
- Kobets, T.; Duan, J.D.; Brunnemann, K.D.; Etter, S.; Smith, B.; Williams, G.M. Structure-activity relationships for DNA damage by alkenylbenzenes in Turkey egg fetal liver. Toxicol. Sci. 2016, 150, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Kobets, T.; Iatropoulos, M.J.; Williams, G.M. Mechanisms of DNA-reactive and epigenetic chemical carcinogens: Applications to carcinogenicity testing and risk assessment. Toxicol. Res. 2019, 8, 123–145. [Google Scholar] [CrossRef] [Green Version]
- Martins, C.; Doran, C.; Silva, I.C.; Miranda, C.; Rueff, J.; Rodrigues, A.S. Myristicin from nutmeg induces apoptosis via the mitochondrial pathway and down regulates genes of the DNA damage response pathways in human leukaemia K562 cells. Chem. Biol. Interact. 2014, 218, 1–9. [Google Scholar] [CrossRef]
- Martins, C.M.d.S. Study of the Natural Alkenylbenzenes Compounds: Mechanisms of DNA Lesions and Implications for Humana Health; Universidade Nova de Lisboa: Lisboa, Portugal, 2016; pp. 1–247. [Google Scholar]
- Martins, C.; Doran, C.; Laires, A.; Rueff, J.; Rodrigues, A.S. Genotoxic and apoptotic activities of the food flavourings myristicin and eugenol in AA8 and XRCC1 deficient EM9 cells. Food Chem. Toxicol. 2011, 49, 385–392. [Google Scholar] [CrossRef]
- Baker, R.S.U.; Bonin, A.M. Tests with the Salmonella plate-incorporation assay. Prog. Mutat. Res. 1985, 5, 177–180. [Google Scholar]
- Green, N.R.; Savage, J.R. Screening of safrole, eugenol, their ninhydrin positive metabolites and selected secondary amines for potential mutagenicity. Mutat. Res. 1978, 57, 115–121. [Google Scholar] [CrossRef]
- Swanson, A.B.; Chambliss, D.D.; Blomquist, J.C.; Miller, E.C.; Miller, J.A. The mutagenicities of safrole, estragole, eugenol, trans-anethole, and some of their known or possible metabolites for Salmonella typhimurium mutants. Mutat. Res. Fundam. Mol. Mech. Mutagen. 1979, 60, 143–153. [Google Scholar] [CrossRef]
- Wislocki, P.G.; Miller, E.C.; Miller, J.A.; Mccoy, E.C.; Rosenkranz, H.S. Carcinogenic and mutagenic activities of safrole, 1′-hydroxysafrole, and some known or possible metabolites. Cancer Res. 1977, 37, 1883–1891. [Google Scholar] [PubMed]
- Poirier, L.A.; de Serres, F.J. Initial National Cancer Institute studies on mutagenesis as a prescreen for chemical carcinogens: An appraisal. J. Natl. Cancer Inst. 1979, 62, 919–926. [Google Scholar] [PubMed]
- Purchase, I.F.; Longstaff, E.; Ashby, J.; Styles, J.A.; Anderson, D.; Lefevre, P.A.; Westwood, F.R. An evaluation of 6 short-term tests for detecting organic chemical carcinogens. Br. J. Cancer 1978, 37, 873–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishidate, M., Jr. The in vitro chromosomal aberration test using Chinese hamster lung (CHL) fibroblast cells in culture. Prog. Mutat. Res. 1985, 5, 427–432. [Google Scholar]
- Myhr, B.; Bowers, L.; Caspary, W. Assays for the induction of gene mutations at the thymidine kinase locus in L5178Y mouse lymphoma cells in culture. Prog. Mutat. Res. 1985, 5, 555–568. [Google Scholar]
- Bradley, M. Measurement of DNA single-strand breaks by alkaline elution in rat hepatocytes. Prog. Mutat. Res. 1985, 5, 353–357. [Google Scholar]
- Epstein, S.S.; Arnold, E.; Andrea, J.; Bass, W.; Bishop, Y. Detection of chemical mutagens by the dominant lethal assay in the mouse. Toxicol Appl Pharm. 1972, 23, 288–325. [Google Scholar] [CrossRef]
- Gocke, E.; King, M.T.; Eckhardt, K.; Wild, D. Mutagenicity of cosmetics ingredients licensed by the European Communities. Mutat. Res. 1981, 90, 91–109. [Google Scholar] [CrossRef]
- Mirsalis, J.C.; Tyson, C.K.; Butterworth, B.E. Detection of genotoxic carcinogens in the in vivo-in vitro hepatocyte DNA repair assay. Env. Mutagen 1982, 4, 553–562. [Google Scholar] [CrossRef]
- Jin, M.; Kijima, A.; Suzuki, Y.; Hibi, D.; Inoue, T.; Ishii, Y.; Nohmi, T.; Nishikawa, A.; Ogawa, K.; Umemura, T. Comprehensive toxicity study of safrole using a medium-term animal model with gpt delta rats. Toxicology 2011, 290, 312–321. [Google Scholar] [CrossRef]
- Matsushita, K.; Kijima, A.; Ishii, Y.; Takasu, S.; Jin, M.; Kuroda, K.; Kawaguchi, H.; Miyoshi, N.; Nohmi, T.; Ogawa, K.; et al. Development of a medium-term animal model using gpt delta rats to evaluate chemical carcinogenicity and genotoxicity. J. Toxicol. Pathol. 2013, 26, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Damhoeri, A.; Hosono, A.; Itoh, T.; Matsuyama, A. In vitro mutagenicity tests on capsicum pepper, shallot and nutmeg oleoresins. Agric. Biol. Chem. 1985, 49, 1519–1520. [Google Scholar] [CrossRef]
- NTP. NTP Technical Report on the Toxicity Studies of Myristicin (CASRN 607-91-0) Administered by Gavage to F344/NTac Rats and B6C3F1/N Mice. Natl. Toxicol. Program Tech. Rep. Ser. 2019, 95, 1–65. [Google Scholar]
- Nohmi, T. Past, Present and Future Directions of gpt delta Rodent Gene Mutation Assays. Food Saf. 2016, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Feuer, G. Action of pregnancy and various progesterones on hepatic microsomal activites. Drug Metab. Rev. 1979, 9, 147–169. [Google Scholar] [CrossRef]
- Vesselinovitch, S.D.; Rao, K.V.; Mihailovich, N. Transplacental and Lactational Carcinogenesis by Safrole. Cancer Res. 1979, 39, 4378–4380. [Google Scholar]
- Rice, J.M. Exposure to chemical carcinogens during pregnancy: Consequences for mother and conceptus. Adv. Exp. Med. Biol. 1984, 176, 13–49. [Google Scholar] [CrossRef]
- Lu, L.J.W.; Disher, R.M.; Randerath, K. Differences in the covalent binding of benzo[a]pyrene, safrole, 1′-hydroxysafrole, and 4-aminobiphenyl to DNA of pregnant and non-pregnant mice. Cancer Lett. 1986, 31, 43–52. [Google Scholar] [CrossRef]
- Vesselinovitch, S.D.; Koka, M.; Rao, K.V.; Mihailovich, N.; Rice, J.M. Prenatal multicarcinogenesis by ethylnitrosourea in mice. Cancer Res. 1977, 37, 1822–1828. [Google Scholar]
- Vesselinovitch, S.D.; Rao, K.V.; Mihailovich, N.; Rice, J.M.; Lombard, L.S. Development of broad spectrum of tumors by ethylnitrosourea in mice and the modifying role of age, sex, and strain. Cancer Res. 1974, 34, 2530–2538. [Google Scholar]
- Giglia-Mari, G.; Zotter, A.; Vermeulen, W. DNA damage response. Cold Spring Harb. Perspect. Biol. 2011, 3, a000745. [Google Scholar] [CrossRef]
- Rodrigues, A.S.; Gomes, B.C.; Martins, C.; Gromicho, M.; Oliveira, N.G.; Guerreiro, P.S.; Rueff, J. DNA repair and resistance to cancer therapy. In New Research Directions in DNA Repair; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef]
- Long, E.; Nelson, A.; Fitzhugh, O.; Hansen, W. Liver tumours produced in rats by feeding safrole. Arch. Pathol. 1963, 75, 595–604. [Google Scholar]
- Borchert, P.; Miller, J.A.; Miller, E.C.; Shires, T.K. 1′-Hydroxysafrole, a Proximate Carcinogenic Metabolite of Safrole in the Rat and Mouse. Cancer Res. 1973, 33, 590–600. [Google Scholar]
- Borchert, P.; Wislocki, P.G.; Miller, J.A.; Miller, E.C. The Metabolism of the Naturally Occurring Hepatocarcinogen Safrole to 1′-Hydroxysafrole and the Electrophilic Reactivity of 1′-Acetoxysafrole. Cancer Res. 1973, 33, 575–589. [Google Scholar] [PubMed]
- Drinkwater, N.R.; Miller, E.C.; Miller, J.A.; Pitot, H.C. Hepatocarcinogenicity of estragole (1-allyl-4-methoxybenzene) and 1′-hydroxyestragole in the mouse and mutagenicity of 1′-acetoxyestragole in bacteria. J. Natl. Cancer Inst. 1976, 57, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.C.; Swanson, A.B.; Phillips, D.H.; Liem, A.; Miller, J.A.; Fletcher, T.L. Structure-activity studies of the carcinogenicities in the mouse and rat of some naturally occurring and synthetic alkenylbenzene derivatives related to safrole and estragole. Cancer Res. 1983, 43, 1124–1134. [Google Scholar] [PubMed]
- Johnson, J.D.; Ryan, M.J.; Toft Ii, J.D.; Graves, S.W.; Hejtmancik, M.R.; Cunningham, M.L.; Herbert, R.; Abdo, K.M. Two-year toxicity and carcinogenicity study of methyleugenol in F344/N rats and B6C3F1 mice. J. Agric. Food Chem. 2000, 48, 3620–3632. [Google Scholar] [CrossRef]
- NTP. Safrole-CAS No. 94-59-7. Report on Carcinogens; 2016; pp. 1–2. Available online: https://ntp.niehs.nih.gov/ntp/roc/content/profiles/safrole.pdf (accessed on 13 May 2022).
- Miller, J.A.; Miller, E.C. The metabolic activation and nucleic acid adducts of naturally-occurring carcinogens: Recent results with ethyl carbamate and the spice flavors safrole and estragole. Br. J. Cancer 1983, 48, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Auerbach, S.S.; Shah, R.R.; Mav, D.; Smith, C.S.; Walker, N.J.; Vallant, M.K.; Boorman, G.A.; Irwin, R.D. Predicting the hepatocarcinogenic potential of alkenylbenzene flavoring agents using toxicogenomics and machine learning. Toxicol. Appl. Pharm. 2010, 243, 300–314. [Google Scholar] [CrossRef]
- Glück, J.; Buhrke, T.; Frenzel, F.; Braeuning, A.; Lampen, A. In silico genotoxicity and carcinogenicity prediction for food-relevant secondary plant metabolites. Food Chem. Toxicol. 2018, 116, 298–306. [Google Scholar] [CrossRef]
- Beroza, M.; Inscoe, M.N.; Schwartz Jr, P.H.; Keplinger, M.L.; Mastri, C.W. Acute toxicity studies with insect attractants. Toxicol. Appl. Pharm. 1975, 31, 421–429. [Google Scholar] [CrossRef]
- Singleton, V.L.; Kratzer, F.H. Toxicity and related physiological activity of phenolic substances of plant origin. J. Agric. Food Chem. 1969, 17, 497–512. [Google Scholar] [CrossRef]
- Liu, G.X.; Xu, F.; Shang, M.Y.; Wang, X.; Cai, S.Q. The Relative Content and Distribution of Absorbed Volatile Organic Compounds in Rats Administered Asari Radix et Rhizoma Are Different between Powder- and Decoction-Treated Groups. Molecules 2020, 25, 4441. [Google Scholar] [CrossRef]
- Taylor, J.; Jones, W.; Hagan, E.; Gross, M.; Davis, D.; Cook, E. Toxicity of oil of calamus (Jammu variety). Toxicol. Appl. Pharm. 1967, 10, 405. [Google Scholar]
- Buchanan, R.L. Toxicity of spices containing methylenedioxybenzene derivatives: A review. J. Food Saf. 1978, 1, 275–293. [Google Scholar] [CrossRef]
- Von Oettingen, W.F. Phenol and Its Derivatives: The Relation between Their Chemical Constitution and Their Effect on the Organism; US Government Printing Office: Washington, DC, USA, 1949. [Google Scholar]
- Truitt, E.B., Jr.; Callaway, E., III; Braude, M.C.; Krantz, J.C., Jr. The pharmacology of myristicin. A contribution to the psychopharmacology of nutmeg. J. Neuropsychiatr. 1961, 2, 205–210. [Google Scholar]
- Abdo, K.M.; Cunningham, M.L.; Snell, M.L.; Herbert, R.A.; Travlos, G.S.; Eldridge, S.R.; Bucher, J.R. 14-Week toxicity and cell proliferation of methyleugenol administered by gavage to F344 rats and B6C3F1 mice. Food Chem. Toxicol. 2001, 39, 303–316. [Google Scholar] [CrossRef]
- Hagan, E.C.; Jenner, P.M.; Jones, W.I.; Fitzhugh, O.G.; Long, E.L.; Brouwer, J.G.; Webb, W.K. Toxic Properties of Compounds Related to Safrole. Toxicol. Appl. Pharm. 1965, 7, 18–24. [Google Scholar] [CrossRef]
- Chiu, S.; Wang, T.; Belski, M.; Abourashed, E.A. HPLC-guided isolation, purification and characterization of phenylpropanoid and phenolic constituents of nutmeg kernel (Myristica fragrans). Nat. Pro. Comm. 2016, 11, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Dang, H.N.P.; Quirino, J.P. High Performance Liquid Chromatography versus Stacking-Micellar Electrokinetic Chromatography for the Determination of Potentially Toxic Alkenylbenzenes in Food Flavouring Ingredients. Molecules 2021, 27, 13. [Google Scholar] [CrossRef]
- Dang, H.N.P.; Quirino, J.P. Analytical Separation of Carcinogenic and Genotoxic Alkenylbenzenes in Foods and Related Products (2010–2020). Toxins 2021, 13, 387. [Google Scholar] [CrossRef]
- Rivera-Pérez, A.; Romero-González, R.; Garrido Frenich, A. Determination and Occurrence of Alkenylbenzenes, Pyrrolizidine and Tropane Alkaloids in Spices, Herbs, Teas, and Other Plant-derived Food Products Using Chromatographic Methods: Review from 2010–2020. Food Rev. Int. 2021, 1–27. [Google Scholar] [CrossRef]
- OECD. Test No. 471: Bacterial Reverse Mutation Test; OECD Publishing: Paris, France, 2020. [Google Scholar] [CrossRef] [Green Version]
- Honda, H.; Minegawa, K.; Fujita, Y.; Yamaguchi, N.; Oguma, Y.; Glatt, H.; Nishiyama, N.; Kasamatsu, T. Modified Ames test using a strain expressing human sulfotransferase 1C2 to assess the mutagenicity of methyleugenol. Genes Env. 2016, 38, 1. [Google Scholar] [CrossRef] [Green Version]
- Monien, B.H.; Herrmann, K.; Florian, S.; Glatt, H. Metabolic activation of furfuryl alcohol: Formation of 2-methylfuranyl DNA adducts in Salmonella typhimurium strains expressing human sulfotransferase 1A1 and in FVB/N mice. Carcinogenesis 2011, 32, 1533–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD. Test No. 476: In Vitro Mammalian Cell Gene Mutation Tests Using the Hprt and Xprt Genes; OECD Publishing: Paris, France, 2016. [Google Scholar] [CrossRef]
- Dobrovolsky, V.N.; Heflich, R.H. Chapter Eleven—Detecting Mutations Vivo. In Mutagenicity: Assays and Applications; Kumar, A., Dobrovolsky, V.N., Dhawan, A., Shanker, R., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 229–249. [Google Scholar] [CrossRef]
- OECD. Test No. 488: Transgenic Rodent Somatic and Germ Cell Gene Mutation Assays; OECD Publishing: Paris, France, 2011. [Google Scholar]
- Lambert, I.B.; Singer, T.M.; Boucher, S.E.; Douglas, G.R. Detailed review of transgenic rodent mutation assays. Mutat. Res. 2005, 590, 1–280. [Google Scholar] [CrossRef] [PubMed]
- Nohmi, T.; Katoh, M.; Suzuki, H.; Matsui, M.; Yamada, M.; Watanabe, M.; Suzuki, M.; Horiya, N.; Ueda, O.; Shibuya, T.; et al. A new transgenic mouse mutagenesis test system using Spi- and 6-thioguanine selections. Env. Mol. Mutagen. 1996, 28, 465–470. [Google Scholar] [CrossRef]
- Nohmi, T.; Masumura, K.; Toyoda-Hokaiwado, N. Transgenic rat models for mutagenesis and carcinogenesis. Genes Env. 2017, 39, 11. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Umemura, T.; Hibi, D.; Inoue, T.; Jin, M.; Ishii, Y.; Sakai, H.; Nohmi, T.; Yanai, T.; Nishikawa, A.; et al. Possible involvement of genotoxic mechanisms in estragole-induced hepatocarcinogenesis in rats. Arch. Toxicol. 2012, 86, 1593–1601. [Google Scholar] [CrossRef]
- SCF. Opinion of the Scientific Committee on Food on Estragole (1-Allyl-4-methoxybenzene). In SCF/CS/FLAV/FLAVOUR/6 ADD2 FINAL; European Commission: Brussels, Belgium, 2001; pp. 1–10. [Google Scholar]


| Substance | Cytochrome P450 Subtype | Reference |
|---|---|---|
| Methyleugenol | CYP1A2 (CYP2C9, 2C19) | [90,92] |
| Elemicin | CYP1A1, CYP1A2, CYP3A4 | [88] |
| Safrole | CYP2A6 (CYP1A2, CYP2C19, CYP2E1) | [89,91] |
| Myristicin | CYP3A4 (CYP1A1) | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Götz, M.E.; Sachse, B.; Schäfer, B.; Eisenreich, A. Myristicin and Elemicin: Potentially Toxic Alkenylbenzenes in Food. Foods 2022, 11, 1988. https://doi.org/10.3390/foods11131988
Götz ME, Sachse B, Schäfer B, Eisenreich A. Myristicin and Elemicin: Potentially Toxic Alkenylbenzenes in Food. Foods. 2022; 11(13):1988. https://doi.org/10.3390/foods11131988
Chicago/Turabian StyleGötz, Mario E., Benjamin Sachse, Bernd Schäfer, and Andreas Eisenreich. 2022. "Myristicin and Elemicin: Potentially Toxic Alkenylbenzenes in Food" Foods 11, no. 13: 1988. https://doi.org/10.3390/foods11131988