
Short vs. Long-Distance Avocado Supply Chains: Life Cycle Assessment Impact Associated to Transport and Effect of Fruit Origin and Supply Conditions Chain on Primary and Secondary Metabolites

 , , , , ,
, , , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Fruit Material and Simulation of Shipping Conditions
2.2. Assessment of the Environmental Impact
2.3. Fruit Quality Evaluations
2.4. Assessment of Primary and Secondary Metabolites
2.4.1. Determination of Fatty Acids
2.4.2. Determination of Polar Metabolites
2.4.3. Determination of Phenolic Compounds
2.4.4. Determination of Abscisic Acid Content by UPLC-PAD
2.4.5. Determination of In Vitro Hydrophilic and Lipophilic Antioxidant Capacity
2.5. Statistical Analysis
3. Results and Discussion
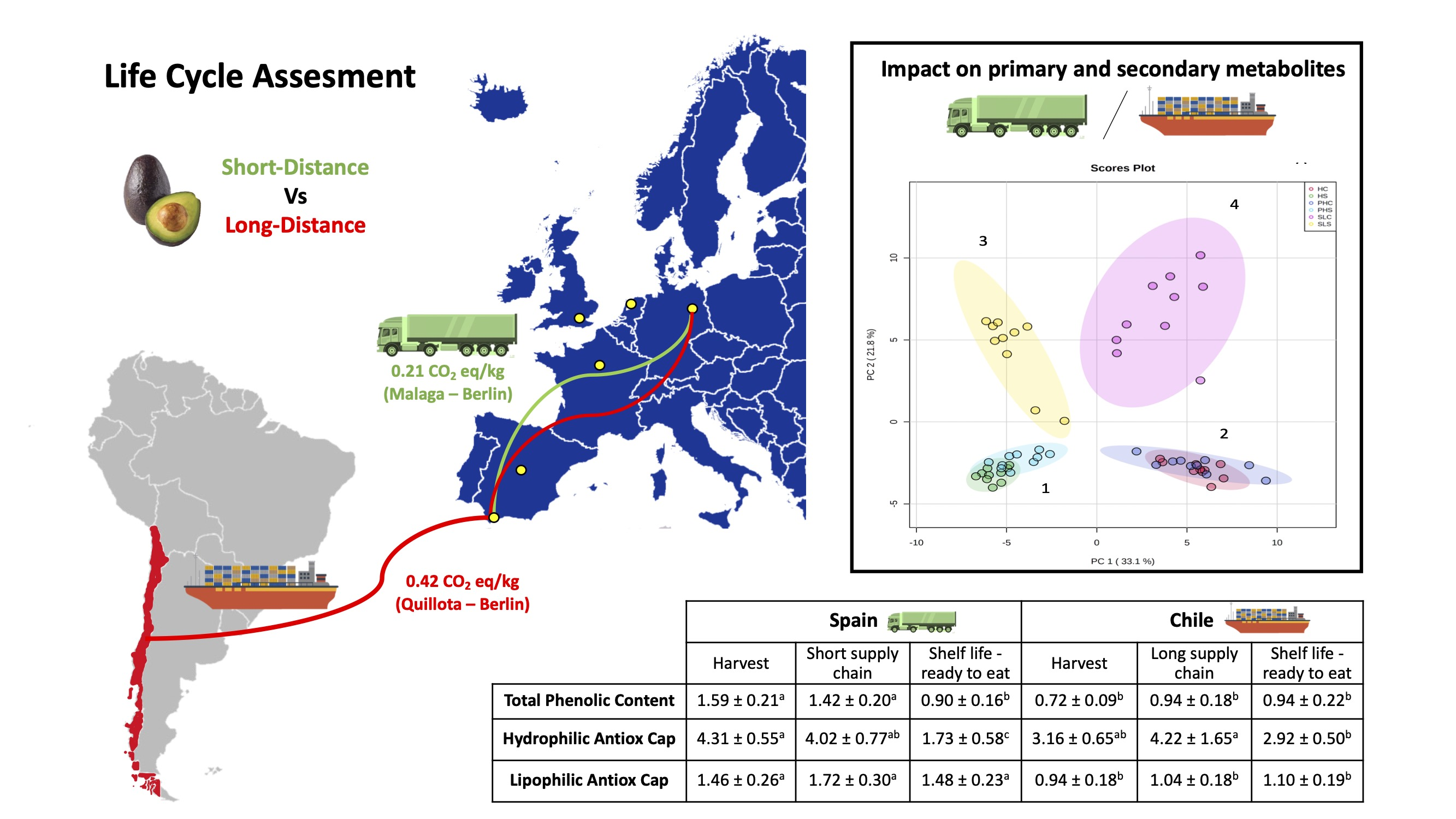
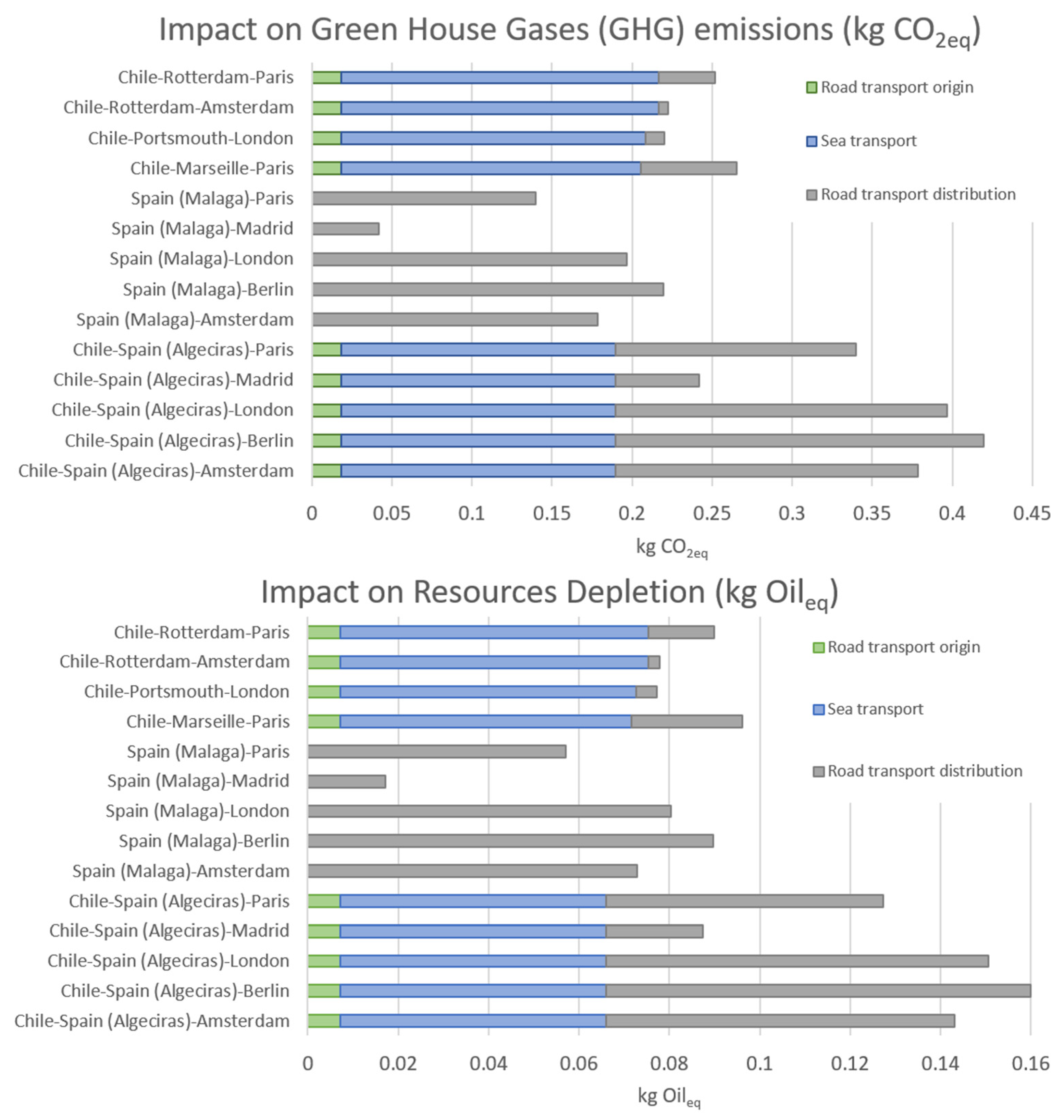
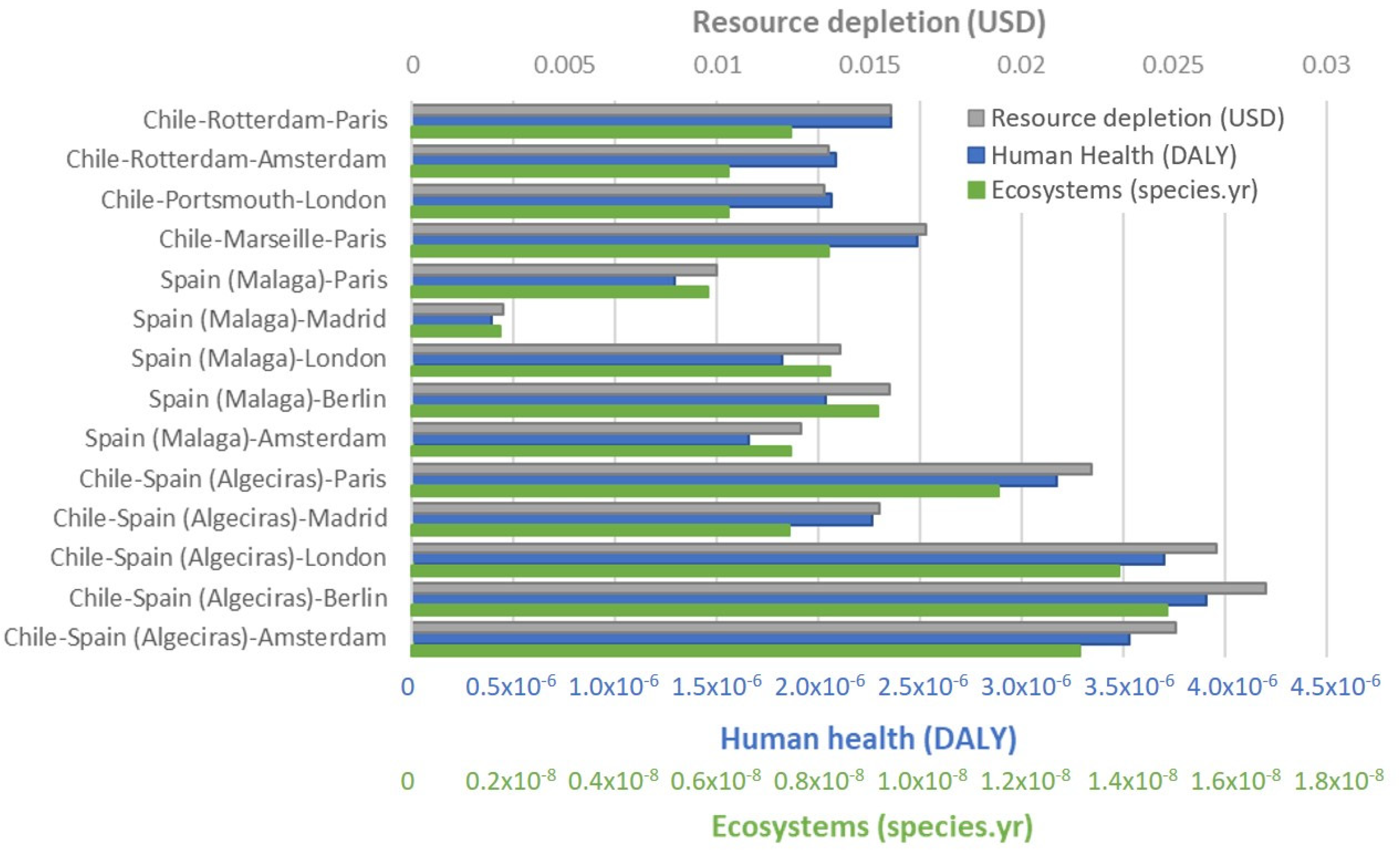
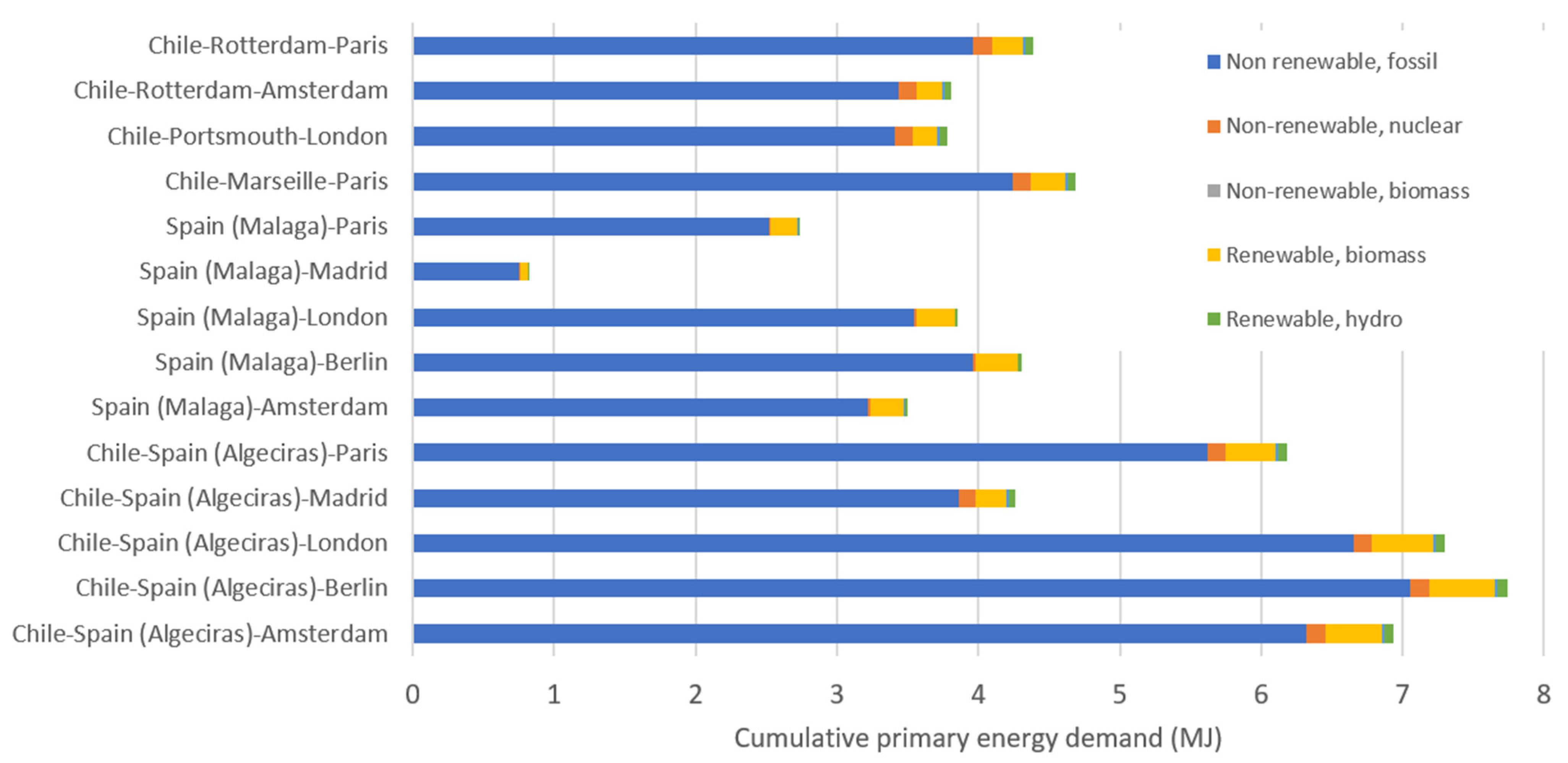
3.1. Differences in Environmental Impacts between Short and Long-Distance Supply Chains
3.2. Differences in Internal and External Quality Attributes in Hass Avocado Fruits from Different Origins and Supply Chain Conditions
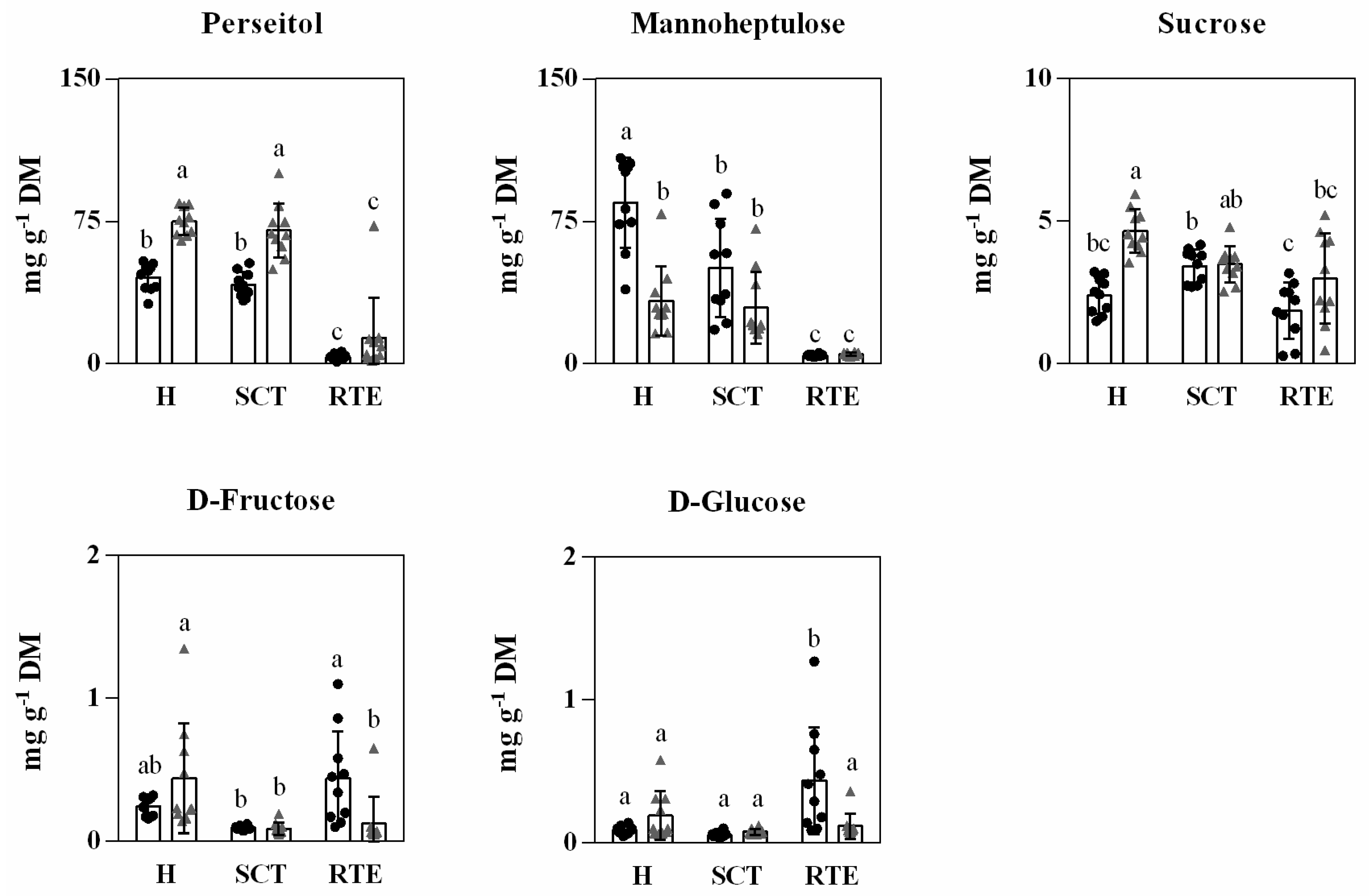
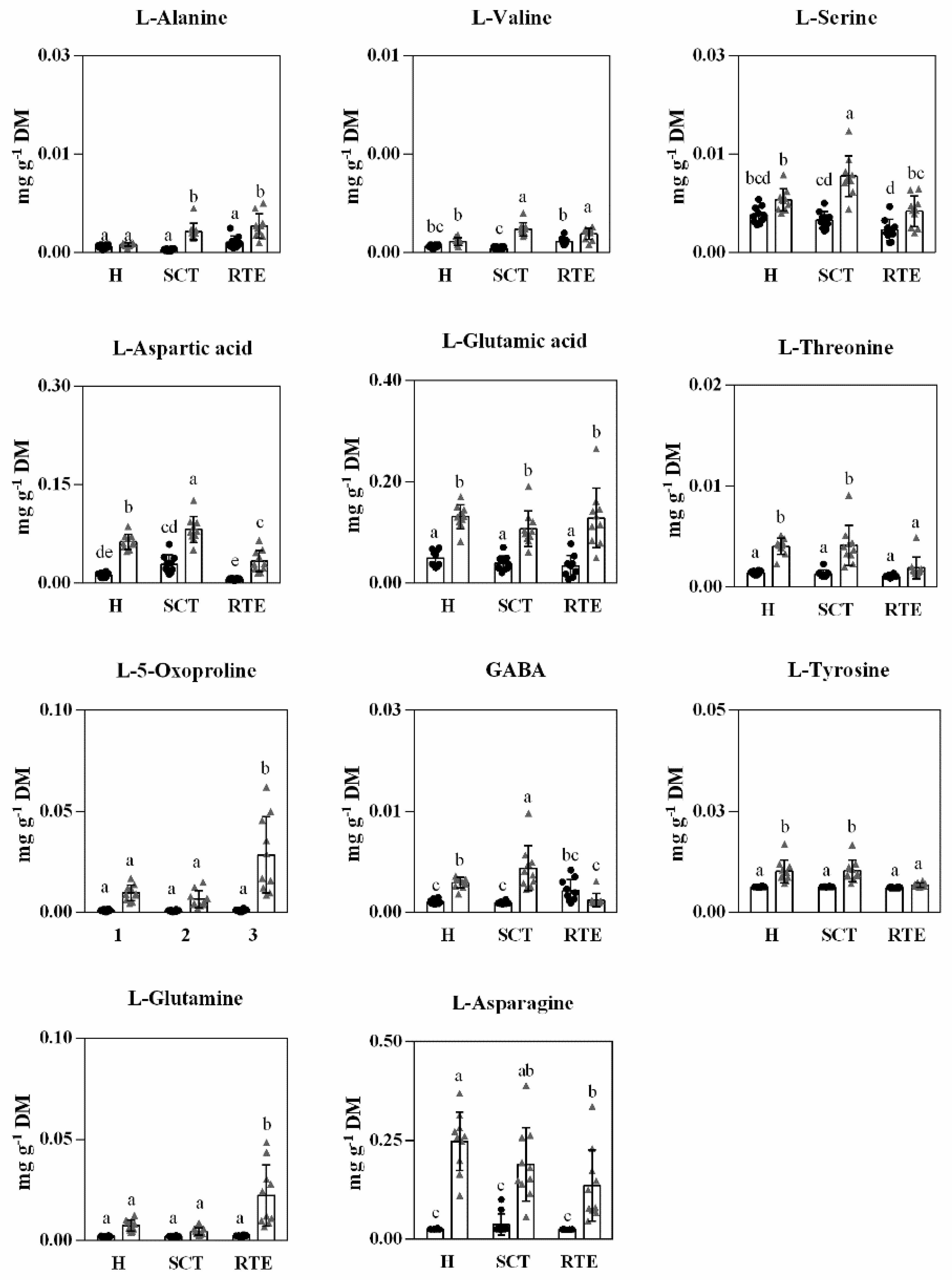
3.3. Differences in Primary Metabolites in Hass Avocado Fruits from Different Origins and Distance and Condition Supply Chains
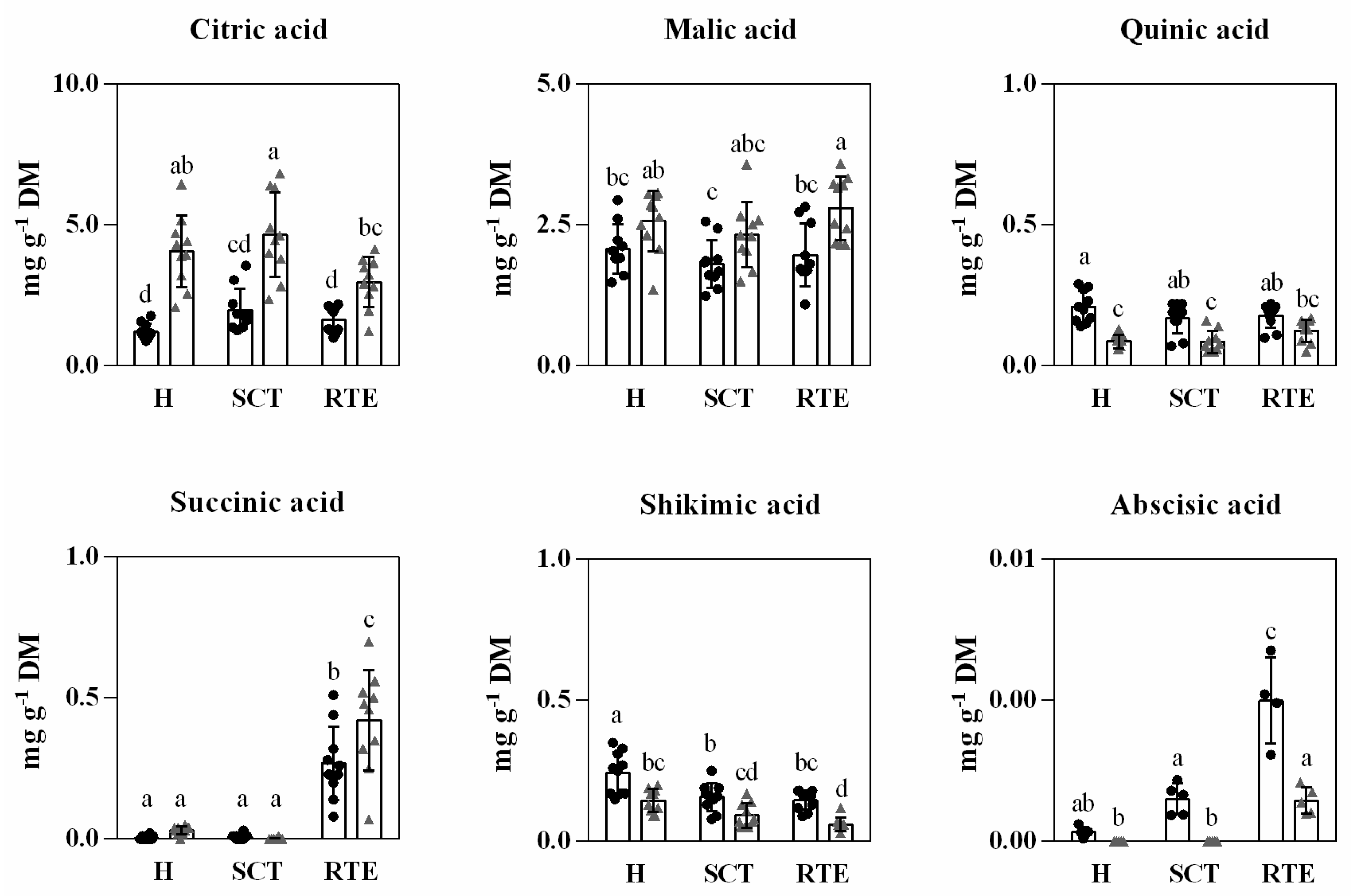
3.4. Differences in Secondary Metabolites and In Vitro Antioxidant Capacity in Hass Avocado Fruit from Different Origins and Distance and Condition Supply Chains
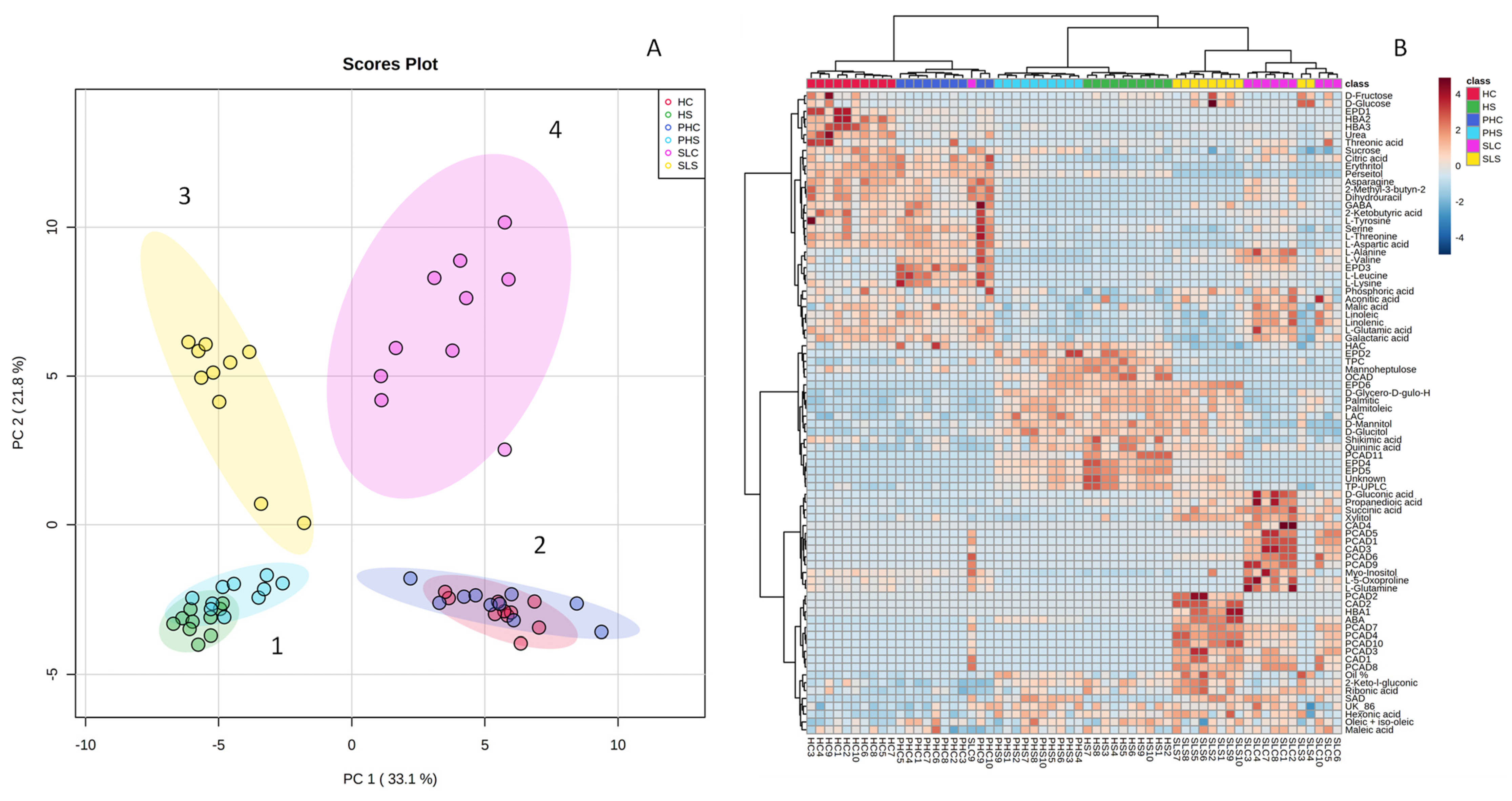
3.5. Integration of All Data by Multivariate Statistical Approaches
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slavin, J.; Lloyd, B. Health benefits of fruits and vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Hausch, B.; Arpaia, M.L.; Campisi-Pinto, S.; Obenland, D. Sensory characterization of two California-grown avocado varieties (Persea americana Mill) over the harvest season by descriptive analysis and consumer tests. J. Food Sci. 2021, 86, 4134–4141. [Google Scholar] [CrossRef]
- Marín-Obispo, L.M.; Villarreal-Lara, R.; Rodríguez-Sánchez, D.G.; Del Follo-Martínez, A.; Espíndola Barquera, M.d.l.C.; Jaramillo-De la Garza, J.S.; Díaz de la Garza, R.I.; Hernández-Brenes, C. Insights into drivers of liking for avocado pulp (Persea americana): Integration of descriptive variables and predictive modeling. Foods 2021, 10, 99. [Google Scholar] [CrossRef]
- Pedreschi, R.; Uarrota, V.; Fuentealba, C.; Alvaro, J.; Olmedo, P.; Defilippi, B.; Meneses, C.; Campos-Vargas, R. Primary metabolism in avocado fruit. Front. Plant Sci. 2019, 10, 795. [Google Scholar] [CrossRef]
- Villa-Rodríguez, J.; Yahia, E.; González-León, A.; Ifie, I.; Robles-Zepeda, R.; Domínguez-Ávila, J.; González-Aguilar, G. Ripening of ‘Hass’ avocado mesocarp alters its phytochemical profile and the in vitro cytotoxic activity of its methanolic extracts. S. Afr. J. Bot. 2020, 128, 1–8. [Google Scholar] [CrossRef]
- Dreher, M.; Davenport, A. ‘Hass’ avocado composition and potential health effects. Crit. Rev. Food Sci. Nutr. 2013, 53, 738–750. [Google Scholar] [CrossRef] [Green Version]
- ASAJA (Asociación Agraria de Jóvenes Agricultores) 2019. ASAJA MÁLAGA Hace Balance: Fin de la Campaña de Aguacate 2018/2019. Available online: https://www.asaja.com/publicaciones/asaja_malaga_hace_balance_fin_de_la_campana_de_aguacate_2018_2019_6864 (accessed on 15 January 2020).
- Frankowska, A.; Jeswani, H.K.; Azapagic, A. Life cycle environmental impacts of fruits consumption in the UK. J. Environ. Manag. 2019, 248, 109111. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, D.; Gutman, A.; Leet, W.; Drewnowski, A.; Fanzo, J.; Ingram, J. Seven food system metrics of sustainable nutrition security. Sustainability 2016, 8, 196. [Google Scholar] [CrossRef] [Green Version]
- González-García, S.; Esteve-Llorens, X.; Moreira, M.T.; Feijoo, G. Carbon footprint and nutritional quality of different human dietary choices. Sci. Total Environ. 2018, 644, 77–94. [Google Scholar] [CrossRef]
- Clark, M.; Springmann, M.; Hill, J.; Tilman, D. Multiple health and environmental impacts of foods. Proc. Natl. Acad. Sci. USA 2019, 116, 23357–23362. [Google Scholar] [CrossRef] [Green Version]
- Hallström, E.; Woodhouse, J.D.; Sovesson, U. Using dietary quality scores to assess sustainability of food products and human diets: A systematic review. Ecol. Indic. 2018, 93, 219–230. [Google Scholar] [CrossRef]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT-Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]
- Moreno-Ortega, G.; Pliego, C.; Sarmiento, D.; Barceló, A.; Martínez-Ferri, E. Yield and fruit quality of avocado trees under different regimes of water supply in the subtropical coast of Spain. Agric. Water Manag. 2019, 221, 192–201. [Google Scholar] [CrossRef]
- Silber, A.; Naor, A.; Cohen, H.; Bar-Noy, B.; Yechieli, N.; Levi, M.; Noy, M.; Peres, M.; Duan, D.; Narkes, K.; et al. Avocado fertilization: Matching the periodic demand for nutrients. Sci. Hortic. 2018, 241, 231–240. [Google Scholar] [CrossRef]
- Beyer, C.; Cuneo, I.; Alvaro, J.E.; Pedreschi, R. Evaluation of aerial and root plant growth behavior, water and nutrient use efficiency and carbohydrate dynamics for Hass avocado grown in a soilless and protected growing system. Sci. Hortic. 2021, 277, 109830. [Google Scholar] [CrossRef]
- Poore, J.; Nemecek, T. Reducing food’s environmental impacts through producers and consumers. Science 2018, 360, 987–992. [Google Scholar] [CrossRef] [Green Version]
- Donetti, M.; Terry, L. Biochemical markers defining growing area and ripening stage of imported avocado fruit cv. Hass. J. Food Compos. Anal. 2014, 34, 90–98. [Google Scholar] [CrossRef]
- Campos, D.; Terán-Hilares, F.; Chirinos, R.; Aguilar-Galvez, A.; García-Ríos, D.; Pacheco-Avalos, A.; Pedreschi, R. Bioactive compounds and antioxidant activity from harvest to edible ripeness of avocado cv. Hass (Persea americana) throughout the harvest seasons. Int. J. Food Sci. Technol. 2020, 55, 2208–2218. [Google Scholar] [CrossRef]
- Chirinos, R.; Campos, D.; Martínez, S.; Llanos, S.; Betallaluz-Pallardel, I.; García-Ríos, D.; Pedreschi, R. Metabolite analysis of avocado cv. Hass subjected to hydrothermal and controlled atmosphere treatments for distant markets. Plants 2021, 10, 2427. [Google Scholar] [CrossRef]
- Villa-Rodríguez, J.A.; Molina-Corral, F.J.; Ayala-Zavala, J.F.; Olivas, G.I.; González-Aguilar, G.A. Effect of maturity stage on the content of fatty acids and antioxidant activity of “Hass” avocado. Food Res. Int. 2011, 44, 1231–1237. [Google Scholar] [CrossRef]
- Iriarte, A.; Yáñez, P.; Villalobos, P.; Huenchuleo, P.; Rebolledo-Leiva, R. Carbon footprint of southern hemisphere fruit exported to Europe: The case of Chilean apple to the UK. J. Clean. Prod. 2021, 293, 126118. [Google Scholar] [CrossRef]
- ISO14044:2006; Life Cycle Assessment. Requirements and Guidelines. ISO: Geneva, Switzerland, 2006.
- Huijbregts, M.A.; Steinmann, Z.J.; Elshout, P.M.; Stam, G.; Verones, F.; Vieira, M.; Zijp, M.; Hollander, A.; van Zelm, R. ReCiPe2016: A harmonised life cycle impact assessment method at midpoint and endpoint level. Int. J. Life Cycle Assess. 2017, 22, 138–147. [Google Scholar] [CrossRef]
- IPCC. Climate change 2013: The physical science basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013; p. 1535. [Google Scholar] [CrossRef] [Green Version]
- Joos, F.; Roth, R.; Fuglestvedt, J.S.; Peters, G.P.; Enting, I.G.; Von Bloh, W.; Brovkin, V.; Burke, E.J.; Eby, M.; Edwards, N.R.; et al. Carbon dioxide and climate impulse response functions for the computation of greenhouse gas metrics: A multi-model analysis. Atmos. Chem. Phys. 2013, 13, 2793–2825. [Google Scholar] [CrossRef] [Green Version]
- Jungbluth, N.; Frischknecht, R. Cumulative energy demand. In Implementation of Life Cycle Impact Assessment Methods; Hischier, R., Weidema, B., Eds.; Ecoinvent centre: St. Gallen, Switzerland, 2010; pp. 33–40. [Google Scholar]
- Hischier, R.; Weidema, B.; Althaus, H.-J.; Bauer, C.; Doka, G.; Dones, R.; Frischknecht, R.; Hellweg, S.; Humbert, S.; Jungbluth, N.; et al. Implementation of Life Cycle Impact Assessment Methods Data v2.2 (2010). Ecoinvent Rep. No. 3. Available online: https://ecoinvent.org/wp-content/uploads/2020/08/201007_hischier_weidema_implementation_of_lcia_methods.pdf (accessed on 15 February 2020).
- Girgenti, V.; Peano, C.; Bounous, M.; Baudino, C. A life cycle assessment of non-renewable energy use and greenhouse gas emissions associated with blueberry and raspberry production in northern Italy. Sci. Total Environ. 2013, 458, 414–418. [Google Scholar] [CrossRef] [Green Version]
- Ecoinvent Database 3.8 (Swiss Database). Available online: https://www.ecoinvent.org/ (accessed on 20 January 2021).
- SimaPro 9.1.1. Software to Measure and Improve the Impact of Your Product Life Cycle. Available online: https://simapro.com/ (accessed on 12 March 2020).
- Available online: http://ports.com/ (accessed on 15 January 2020).
- Available online: https://www.google.es/maps/ (accessed on 31 January 2020).
- Weidema, B.P.; Bauer, C.; Hischier, R.; Mutel, C.; Nemecek, T.; Reinhard, J.; Vadenbo, C.O.; Wernet, G. Overview and Methodology. Data Quality Guideline for the Ecoinvent Database Version 3; The Ecoinvent Centre: St. Gallen, Switzerland, 2013. [Google Scholar]
- Rivera, S.; Ferreyra, R.; Robledo, P.; Selles, G.; Arpaia, M.L.; Saavedra, J.; Defilippi, B. Identification of preharvest factors determining postharvest ripening behavior in Hass avocado under long term storage. Sci. Hortic. 2017, 216, 29–37. [Google Scholar] [CrossRef]
- Meurens, M.; Baeten, V.; Yan, S.H.; Mignolet, E.; Larondelle, Y. Determination of the conjugated linoleic acids in cow’s milk fat by Fourier transform Raman spectroscopy. J. Agric. Food Chem. 2005, 53, 5831–5835. [Google Scholar] [CrossRef]
- Hatoum, D.; Annaratone, C.; Hertog, M.L.A.T.M.; Geeraerd, A.; Nicolai, B. Targeted metabolomics study of ‘Braeburn’ apples during long-term storage. Postharvest Biol. Technol. 2014, 96, 33–41. [Google Scholar] [CrossRef]
- Fuentealba, C.; Hernandez, I.; Saa, S.; Toledo, L.; Burdiles, P.; Chirinos, C.; Campos, D.; Brown, P.; Pedreschi, R. Color and in vitro quality attributes of walnuts from different growing conditions correlate with key precursors of primary and secondary metabolism. Food Chem. 2017, 232, 664–672. [Google Scholar] [CrossRef]
- Uarrota, V.; Hernandez, I.; Ponce Guequen Vidal, J.; Fuentealba, C.; Defilippi, B.; Lindh, V.; Zulueta, C.; Chirinos, R.; Campos, D.; Pedreschi, R. Unraveling factors associated with “Blackspot” disorder in stored Hass avocado (Persea americana Mill) fruit. J. Hortic. Sci. Biotechnol. 2020, 95, 804–815. [Google Scholar] [CrossRef]
- Arnao, M.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- CBI Ministry of Foreign Affairs. 2021. Available online: https://www.cbi.eu/market-information/fresh-fruit-vegetables/avocados/market-potential (accessed on 18 March 2021).
- Roibás, L.; Elbehri, A.; Hospido, A. Carbon footprint along the Ecuadorian banana supply chain: Methodological improvements and calculation tool. J. Clean. Prod. 2016, 112, 2441–2451. [Google Scholar] [CrossRef]
- Pedreschi, R.; Hollak, S.; Harkema, H.; Otma, E.; Robledo, P.; Westra, E.; Somhorst, D.; Ferreyra, R.; Defilippi, B. Impact of postharvest ripening strategies on Hass avocado fatty acid profiles. S. Afr. J. Bot. 2016, 103, 32–35. [Google Scholar] [CrossRef]
- Huamán-Alvino, C.; Chirinos, R.; Gonzales-Parinova, F.; Pedreschi, R.; Campos, D. Physicochemical and bioactive compounds at edible ripeness of eleven varieties of avocado (Persea americana) cultivated in the Andean Region of Peru. Int. J. Food Sci. Technol. 2021, 56, 5040–5049. [Google Scholar] [CrossRef]
- Hernández, I.; Uarrota, V.; Paredes, D.; Fuentealba, C.; Defilippi, B.; Campos-Vargas, R.; Meneses, C.; Hertog, M.; Pedreschi, R. Can metabolites at harvest be used as physiological markers for modelling the softening behaviour of Chilean Hass avocados destined to local and distant markets? Postharvest Biol. Technol. 2021, 174, 111457. [Google Scholar] [CrossRef]
- Ramos-Aguilar, A.; Ornelas-Paz, J.; Tapia-Vargas, L.; Gardia-Béjar, A.; Yahia, E.; Ornelas-Paz, J.J.; Pérez-Martínez, J.; Ríos-Velasco, C.; Escalona-Minalota, P. Metabolomic analysis and physical attributes of ripe fruits from Mexican Creole (Persea americana var. Prymifolia) and Hass avocados. Food Chem. 2021, 354, 129571. [Google Scholar] [CrossRef]
- Olivares, D.; Alvarez, E.; Véliz, D.; García-Rojas, M.; Díaz, C.; Defilippi, B. Effects of 1-Methylcyclopropene and controlled atmosphere on ethylene synthesis and quality attributes of avocados cvs. Edranol and Fuerte. J. Food Qual. 2020, 2020, 5075218. [Google Scholar] [CrossRef]
- Defilippi, B.; Ferreyra, R.; Rivera, S. “Sabor en Palta Hass: Algo Más Que Sólo Aceite” in Optimización de la Calidad de Palta “Hass”; Bulletin INIA No. 307; Instituto de Investigaciones Agropecuarias, INIA: Santiago, Chile, 2015; pp. 103–119. [Google Scholar]
- Bochkov, D.; Sysolyatin, S.; Kalashnikov, A.; Sumacheka, I. Shikimic acid: Review of its analytical isolation and purification techniques from plant and microbial sources. J. Chem. Biol. 2012, 5, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.; Chope, G.; Terry, L. Investigation into the role of endogenous abscisic acid during ripening of imported avocado cv. Hass. J. Sci. Food Agric. 2017, 97, 3656–3664. [Google Scholar] [CrossRef] [Green Version]
- Vincent, C.; Mesa, T.; Munné-Bosch, S. Hormonal interplay in the regulation of fruit ripening and cold acclimation in avocados. J. Plant Physiol. 2020, 251, 153225. [Google Scholar] [CrossRef]
- Pedreschi, R.; Muñoz, P.; Robledo, P.; Becerra, C.; Defilippi, B.; van Eekelen, H.; Mumm, R.; Westra, E.; de Vos, R. Metabolomics analysis of postharvest ripening heterogeneity of ‘Hass’ avocadoes. Postharvest Biol. Technol. 2014, 92, 172–179. [Google Scholar] [CrossRef]
- Brizzolara, S.; Manganaris, G.; Fotopoulos, V.; Watkins, C.; Tonutti, P. Primary metabolism in fresh fruits during storage. Front. Plant Sci. 2020, 11, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narsai, R.; Rocha, M.; Geigenberger, P.; Whelan, J.; Van Dongen, J.T. Comparative analysis between plant species of transcriptional and metabolic responses to hypoxia. New Phytol. 2011, 190, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Cukrov, D.; Zermiani, M.; Brizzolara, S.; Cestaro, A.; Licausi, F.; Luchinat, C.; Santucci, C.; Tenori, L.; Van Veen, H.; Zuccolo, A.; et al. Extreme hypoxic conditions induce selective molecular responses and metabolic reset in detached apple fruit. Front. Plant Sci. 2016, 7, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mpai, S.; Sivakumar, D. Influence of growing seasons and metabolic composition and fruit quality of avocado cultivar at the “ready to eat stage”. Sci. Hortic. 2020, 265, 109159. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Country of Origin to Port (by Road) | Distance to Export Country (Port to Port) | Main European Countries Importing Avocados | Distance by Road from Port to City |
|---|---|---|---|---|
| Chile | 116 km (Quillota–San Antonio) | Rotterdam 18,020 km | The Netherlands/France | 78 km to Amsterdam 456 km to Paris |
| Marseille 17,011 km | France | 776 km to Paris | ||
| Portsmouth 17,309 km | United Kingdom | 144 km to London | ||
| Chile | 116 km (Quillota–San Antonio) | Algeciras 15,570 km | France | 1938 km to Paris |
| The Netherlands | 2437 km to Amsterdam | |||
| United Kingdom | 2673 km to London | |||
| Germany | 2964 km to Berlin | |||
| Spain | 675 km to Madrid | |||
| Spain | -- | 0 km (Production in Malaga) | France | 1802 km to Paris |
| The Netherlands | 2301 km to Amsterdam | |||
| United Kingdom | 2537 km to London | |||
| Germany | 2832 km to Berlin | |||
| Spain | 539 km to Madrid |
| Spain | Chile | |||||
|---|---|---|---|---|---|---|
| Harvest | Short Supply Chain | Shelf Life—Ready-to-Eat | Harvest | Long Supply Chain | Shelf Life—Ready-to-Eat | |
| Oil (%) | 11.83 ± 2.00 a | 14.74 ± 1.82 b | 18.27 ± 2.91 c | 10.05 ± 0.57 a | 10.94 ± 0.65 a | 10.84 ± 1.13 a |
| Fatty acids | ||||||
| Palmitic | 197.63 ± 17.46 a | 182.50 ± 23.10 a | 179.06 ± 23.88 a | 115.40 ± 10.69 b | 131.78 ± 13.64 b | 126.54 ± 23.57 b |
| Palmitoleic | 95.38 ± 8.79 a | 89.49 ± 16.11 a | 85.38 ± 13.46 a | 42.18 ± 3.75 b | 47.82 ± 6.10 b | 47.53 ± 7.86 b |
| Oleic | 511.20 ± 61.63 a | 485.32 ± 66.70 a,b | 470.12 ± 76.80 a,b | 435.17 ± 34.3 b | 505.08 ± 50.1 a,b | 487.84 ± 42.0 a,b |
| Linoleic | 134.96 ± 9.28 a | 131.84 ± 18.50 a | 120.95 ± 21.25 a | 169.44 ± 20.50 b | 189.65 ± 19.78 b | 194.93 ± 33.60 b |
| α-linolenic | 9.17 ± 0.93 a | 9.24 ± 1.76 a | 8.34 ± 1.70 a | 14.73 ± 2.15 b | 15.26 ± 1.80 b | 18.78 ± 3.56 c |
| Spain | Chile | |||||
|---|---|---|---|---|---|---|
| Harvest | Short Supply Chain | Shelf Life—Ready-to-Eat | Harvest | Long Supply Chain | Shelf Life—Ready-to-Eat | |
| TPC | 1.59 ± 0.21 a | 1.42 ± 0.20 a | 0.90 ± 0.16 b | 0.72 ± 0.09 b | 0.94 ± 0.18 b | 0.94 ± 0.22 b |
| HAC | 4.31 ± 0.55 a | 4.02 ± 0.77 a,b | 1.73 ± 0.58 c | 3.16 ± 0.65 a,b | 4.22 ± 1.65 a | 2.92 ± 0.50 b |
| LAC | 1.46 ± 0.26 a | 1.72 ± 0.30 a | 1.48 ± 0.23 a | 0.94 ± 0.18 b | 1.04 ± 0.18 b | 1.10 ± 0.19 b |
| Spain | Chile | ||||||
|---|---|---|---|---|---|---|---|
| Peak Number | Phenolic Compound Assigned (mg GAE 100 g−1 DW) | Harvest | Short Supply Chain | Shelf Life—Ready to Eat | Harvest | Long Supply Chain | Shelf Life—Ready to Eat |
| 1 | Hydroxybenzoic acid derivative 1 (HBA1) | 0.0 a | 0.0 a | 0.30 ± 0.16 b | 0.0 a | 0.0 a | 0.0 a |
| 2 | Epicatechin derivative 1 (EPD 1) | 0.0 a | 0.0 a | 0.0 a | 3.05 ± 2.42 b | 0.76 ± 0.30 a | 0.0 a |
| 3 | Syringic acid derivative (SAD) | 0.0 a | 0.11 ± 0.04 b | 0.11 ± 0.05 b | 0.0 a | 0.0 a | 0.11 ± 0.04 b |
| 4 | Hydroxybenzoic acid derivative 2 (HBA2) | 0.0 a | 0.0 a | 0.0 a | 1.03 ± 0.54 b | 0.15 ± 0.08 a | 0.0 a |
| 5 | Hydroxybenzoic acid derivative 3 (HBA3) | 0.13 ± 0.05 a | 0.80 ± 0.49 a | 0.87 ± 0.15 a | 4.21 ± 1.85 b | 1.05 ± 0.37 a | 0.90 ± 0.17 a |
| 6 | p-Coumaric acid derivative 1 (PCAD1) | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 1.20 ± 0.31 b |
| 7 | p-Coumaric acid derivative 2 (PCAD2) | 0.0 a | 0.0 a | 0.41 ± 0.24 b | 0.0 a | 0.0 a | 0.0 a |
| 8 | p-Coumaric acid derivative 3 (PCAD3) | 0.0 a | 0.0 a | 0.22 ± 0.10 b | 0.0 a | 0.0 a | 0.13 ± 0.07 b |
| 9 | Caffeic acid derivative 1 (CAD1) | 0.0 a | 0.0 a | 0.11 ± 0.05 b | 0.0 a | 0.0 a | 0.09 ± 0.03 b |
| 10 | Epicatechin derivative 2 (EPD2) | 0.36 ± 0.22 a | 0.42 ± 0.28 a | 0.0 b | 0.0 b | 0.0 b | 0.0 b |
| 11 | p-Coumaric acid derivative 4 (PCAD4) | 0.0 a | 0.0 a | 0.58 ± 0.09 b | 0.0 a | 0.0 a | 0.27 ± 0.08 c |
| 12 | p-Coumaric acid derivative 5 (PCAD5) | 0.0 a | 0.0 a | 0.04 ± 0.01 a | 0.0 a | 0.0 a | 0.12 ± 0.05 b |
| 13 | p-Coumaric acid derivative 6 (PCAD6) | 0.0 a | 0.0 a | 1.25 ± 0.21 b | 0.0 a | 0.0 a | 3.75 ± 1.16 c |
| 14 | Caffeic acid derivative 2 (CAD2) | 0.0 a | 0.0 a | 0.11 ± 0.03 b | 0.0 a | 0.0 a | 0.0 a |
| 15 | p-Coumaric acid derivative 7 (PCAD7) | 0.0 a | 0.0 a | 1.04 ± 0.13 b | 0.0 a | 0.0 a | 0.82 ± 0.11 c |
| 16 | p-Coumaric acid derivative 8 (PCAD8) | 0.0 a | 0.0 a | 0.14 ±0.03 b | 0.0 a | 0.0 a | 0.13 ± 0.07 b |
| 17 | Caffeic acid derivative 3 (CAD3) | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.03 ± 0.02 b |
| 18 | Caffeic acid derivative 4 (CAD4) | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.03 ± 0.00 b |
| 19 | p-Coumaric acid derivative 9 (PCAD9) | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.0 a | 0.06 ± 0.03 b |
| 20 | o-Coumaric acid derivative (OCAD) | 0.12 ± 0.05 a | 0.08 ± 0.05 a | 0.0 b | 0.0 b | 0.0 b | 0.0 b |
| 21 | Epicatechin derivative 3 (EPD3) | 0.0 a | 0.0 a | 0.0 a | 0.22 ± 0.13 a | 7.96 ± 1.99 b | 2.87 ± 1.08 c |
| 22 | p-Coumaric acid derivative 10 (PCAD10) | 0.0 a | 0.0 a | 0.34 ± 0.10 b | 0.0 a | 0.0 a | 0.15 ± 0.04 c |
| 23 | Unknown * | 5.54 ± 2.46 a | 3.38 ± 1.32 a | 3.22 ± 1.33 a | 0.0 b | 0.0 b | 0.0 b |
| 24 | p-Coumaric acid derivative 11 (PCAD11) | 0.41 ± 0.15 a | 0.12 ± 0.05 bc | 0.24 ± 0.05 b | 0.0 c | 0.0 c | 0.0 c |
| 25 | Epicatechin derivative 4 (EPD4) | 7.29 ± 2.04 a | 4.51 ± 0.92 b | 2.21 ± 0.51 c | 0.0 d | 0.0 d | 0.0 d |
| 26 | Epicatechin derivative 5 (EPD5) | 15.72 ± 2.71 a | 8.23 ± 2.05 b | 5.05 ± 1.23 c | 0.04 ± 0.02 d | 0.0 d | 0.0 d |
| 27 | Epicatechin derivative 6 (EPD6) | 0.46 ± 0.07 ab | 0.43 ± 0.25 b | 0.69 ± 0.15 a | 0.0 c | 0.0 c | 0.0 c |
| Total | 30.03 ± 6.83 a | 18.09 ± 4.16 b | 17.10 ± 3.14 bc | 8.56 ± 3.73 d | 9.93 ± 2.08 cd | 10.65 ± 2.05 bcd | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pedreschi, R.; Ponce, E.; Hernández, I.; Fuentealba, C.; Urbina, A.; González-Fernández, J.J.; Hormaza, J.I.; Campos, D.; Chirinos, R.; Aguayo, E. Short vs. Long-Distance Avocado Supply Chains: Life Cycle Assessment Impact Associated to Transport and Effect of Fruit Origin and Supply Conditions Chain on Primary and Secondary Metabolites. Foods 2022, 11, 1807. https://doi.org/10.3390/foods11121807
Pedreschi R, Ponce E, Hernández I, Fuentealba C, Urbina A, González-Fernández JJ, Hormaza JI, Campos D, Chirinos R, Aguayo E. Short vs. Long-Distance Avocado Supply Chains: Life Cycle Assessment Impact Associated to Transport and Effect of Fruit Origin and Supply Conditions Chain on Primary and Secondary Metabolites. Foods. 2022; 11(12):1807. https://doi.org/10.3390/foods11121807
Chicago/Turabian StylePedreschi, Romina, Excequel Ponce, Ignacia Hernández, Claudia Fuentealba, Antonio Urbina, Jose J. González-Fernández, Jose I. Hormaza, David Campos, Rosana Chirinos, and Encarna Aguayo. 2022. "Short vs. Long-Distance Avocado Supply Chains: Life Cycle Assessment Impact Associated to Transport and Effect of Fruit Origin and Supply Conditions Chain on Primary and Secondary Metabolites" Foods 11, no. 12: 1807. https://doi.org/10.3390/foods11121807