
Bacterial Diversity and Lactic Acid Bacteria with High Alcohol Tolerance in the Fermented Grains of Soy Sauce Aroma Type Baijiu in North China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Bacterial Microbial Diversity Analysis of Fermented Grains
2.3. Isolation and Purification of Bacterial Strains
2.4. Identification of the Selected Strains by 16S rDNA Sequence Analysis
2.5. Determination of Lactic Acid Bacteria Tolerance
2.6. Determination of Adhesion Ability of Lactic Acid Bacteria
3. Results and Discussion
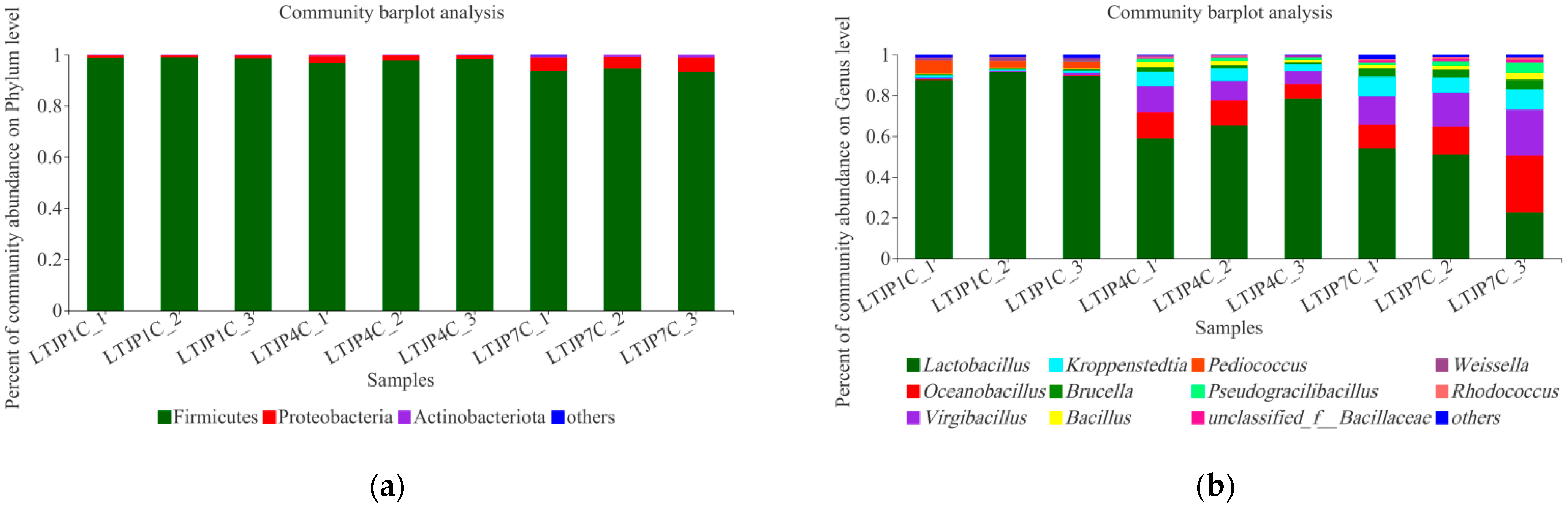
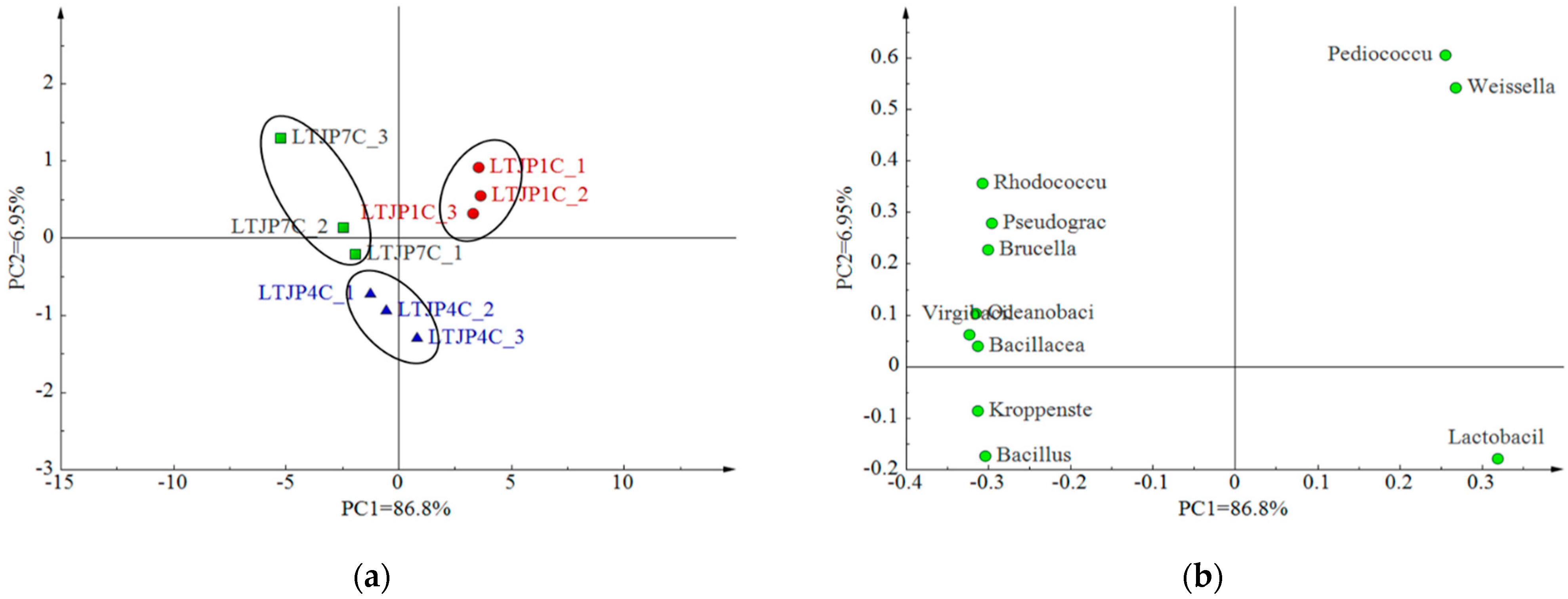
3.1. Bacterial Microbial Diversity during Baijiu Fermentation
3.2. Isolation and Identification of LAB in the Baijiu Fermentation Process
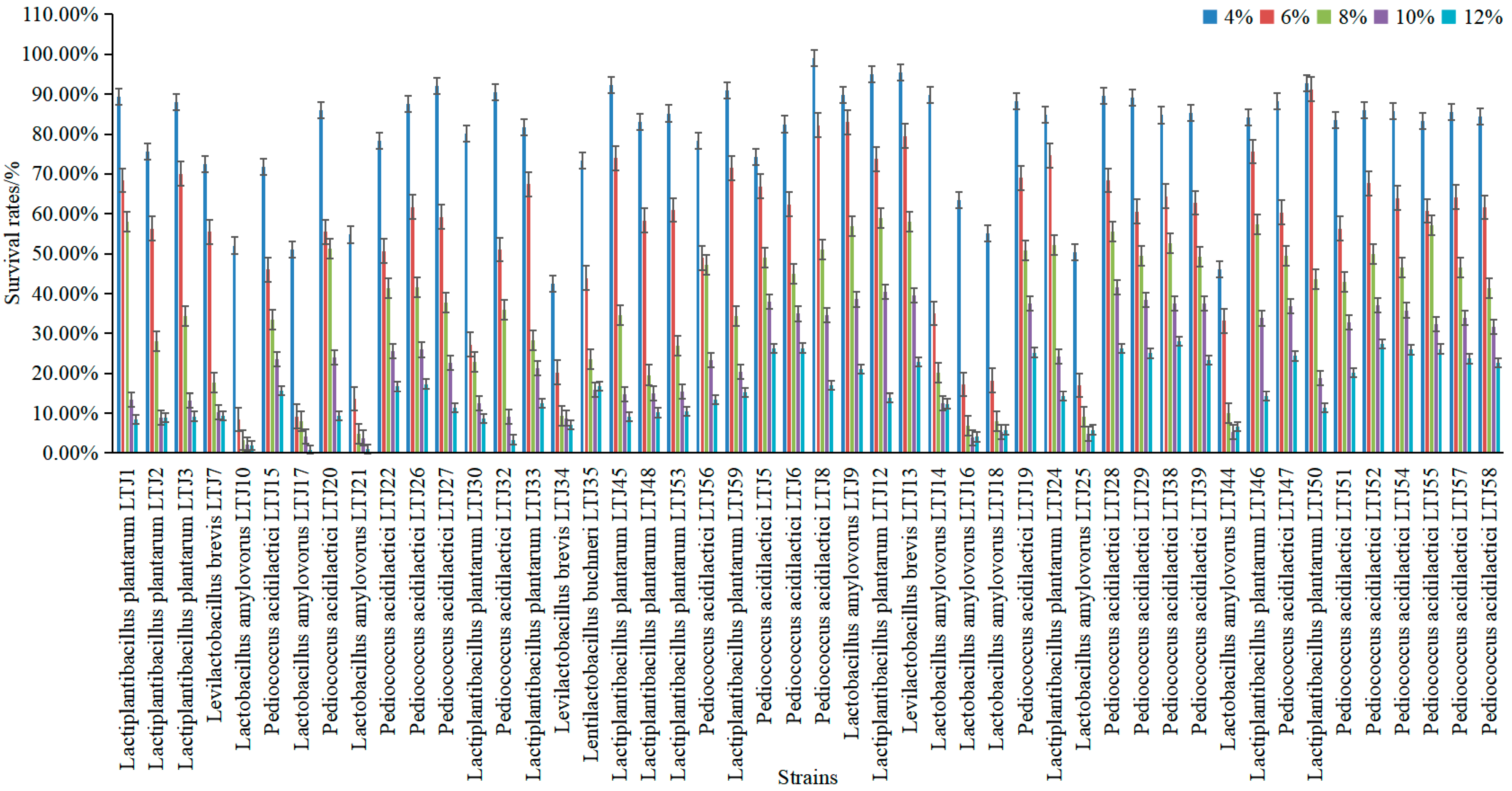
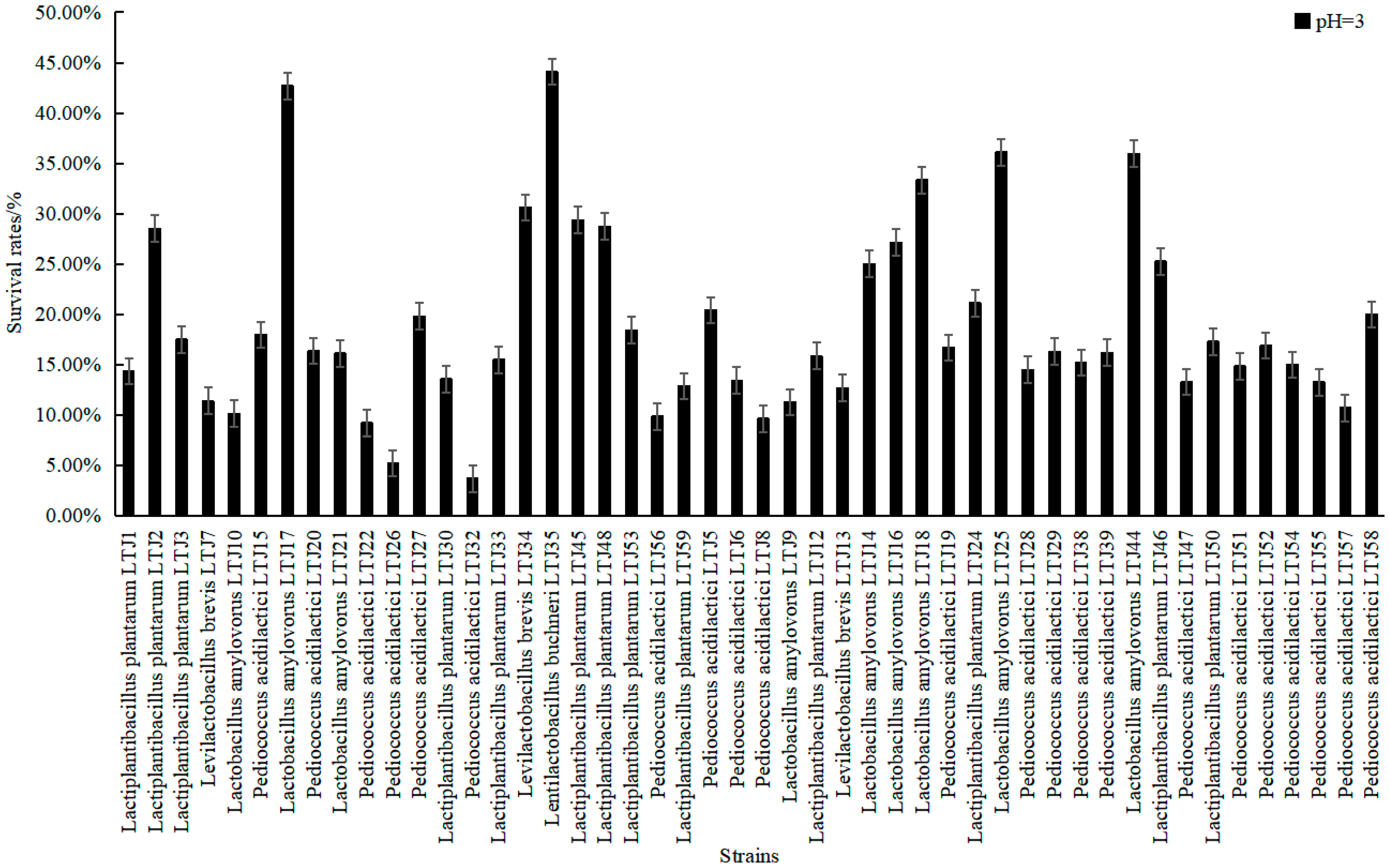
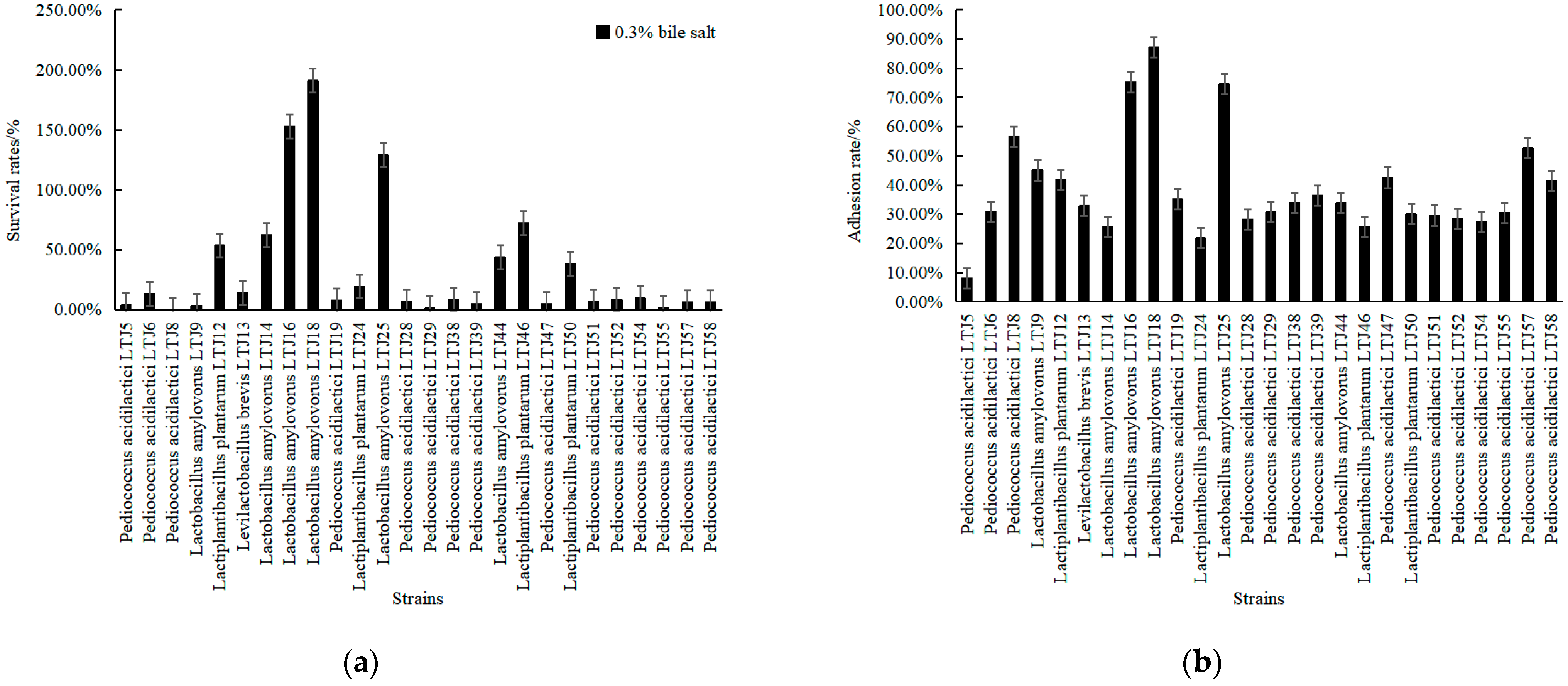
3.3. Screening of Alcohol-Tolerant LABs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.; Du, H.; Zhang, Y.; Xu, Y. Environmental microbiota drives microbial succession and metabolic profiles during Chinese liquor fermentation. Appl. Environ. Microb. 2018, 84, e02369-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Gong, X.; Zhao, G.; Htet, M.; Jia, Z.; Yan, Z.; Liu, L.; Zhai, Q.; Huang, T.; Deng, X. Liquor flavour is associated with the physicochemical property and microbial diversity of fermented grains in waxy and non-waxy sorghum (Sorghum bicolor) during fermentation. Front. Microbiol. 2021, 12, 618458. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Lei, X.; Zhang, X.; Guan, T.; Zhao, S. Characteristics of the microbial community in the production of Chinese Rice-flavor baijiu and comparisons with the microflora of other flavors of baijiu. Front. Microbiol. 2021, 12, 673670. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Wu, Z.; Zhang, W. Effect of pit mud on bacterial community and aroma components in yellow water and their changes during the fermentation of Chinese Strong-flavor liquor. Foods 2020, 9, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Zou, B.; Zhu, T.; Hozzein, W.N.; Quan, Z.; Smidt, H. Modified RNA-seq method for microbial community and diversity analysis using rRNA in different types of environmental samples. PLoS ONE 2017, 12, e0186161. [Google Scholar] [CrossRef]
- Wang, L. Research trends in Jiang-flavor baijiu fermentation: From fermentation microecology to environmental ecology. J. Food Sci. 2022, 87, 1362–1374. [Google Scholar] [CrossRef]
- Cui, F.; Li, Y.; Gao, D.; Chen, J. Microbial diversity and its relationships with stress factors in sesame-flavor liquor fermentation process. Microbiol. China 2019, 46, 2620–2629. [Google Scholar]
- Zheng, J.; Wu, C.; Huang, J.; Zhou, R.; Liao, X. Spatial distribution of bacterial communities and related biochemical properties in Luzhou-flavor liquor-fermented grains. J. Food Sci. 2015, 79, M2491–M2498. [Google Scholar] [CrossRef]
- Dysvik, A.; Rosa, S.; Liland, K.H.; Myhrer, K.S.; Wicklund, T. Co-fermentation involving Saccharomyces Cerevisiae and Lactobacillus species tolerant to brewing-related stress factors for controlled and rapid production of sour beer. Front. Microbiol. 2020, 11, 279. [Google Scholar] [CrossRef]
- Liong, M.T. Safety of probiotics: Translocation and infection. Nutr. Rev. 2008, 66, 192–202. [Google Scholar] [CrossRef]
- Zhang, M.; Yao, M.; Lai, T.; Zhao, H.; Wang, Y.; Yang, Z. Response of Lactiplantibacillus plantarum NMGL2 to combinational cold and acid stresses during storage of fermented milk as analyzed by data-independent acquisition proteomics. Foods 2021, 10, 1514. [Google Scholar] [CrossRef] [PubMed]
- Spano, G.; Massa, S. Environmental stress response in wine lactic acid bacteria: Beyond Bacillus subtilis. Crit. Rev. Microbiol. 2006, 32, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Eva, G.A.; Isabel, L.; Ignacio, R.J.; Julio, S.; Eva, F.E.; Myriam, Z.; Marta, D.; Carmen, T.; Fe Rnanda, R.L. High tolerance of wild Lactobacillus plantarum and Oenococcus oeni strains to lyophilisation and stress environmental conditions of acid pH and ethanol. Fems. Microbiol. Lett. 2004, 230, 53–61. [Google Scholar]
- Haddaji, N.; Mahdhi, A.K.; Ismaiil, M.B.; Bakhrouf, A. Effect of environmental stress on cell surface and membrane fatty acids of Lactobacillus plantarum. Arch. Microbiol. 2017, 199, 1243–1250. [Google Scholar] [CrossRef]
- Ma, R.; Sui, L.; Zhang, J.; Hu, J.; Liu, P. Polyphasic characterization of yeasts and lactic acid bacteria metabolic contribution in Semi-solid fermentation of Chinese baijiu (traditional fermented alcoholic drink): Towards the design of a tailored starter culture. Microorganisms 2019, 7, 147. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Hao, F.; Lv, X.; Chen, B.; Wang, L. Diversity of lactic acid bacteria in Moutai-flavor liquor fermentation process. Food Biotechnol. 2020, 34, 212–227. [Google Scholar] [CrossRef]
- Yang, F.; Chen, L.; Liu, Y.; Li, J.; Wang, L.; Chen, J. Identification of microorganisms producing lactic acid during solid-state fermentation of Maotai flavour liquor. J. Inst. Brew. 2019, 125, 171–177. [Google Scholar] [CrossRef]
- Yang, F.; Wang, Y.; Zhao, S.; Wang, Y. Lactobacillus plantarum inoculants delay spoilage of high moisture alfalfa silages by regulating bacterial community composition. Front. Microbiol. 2020, 11, 1989. [Google Scholar] [CrossRef]
- Wang, L.; Huang, Y.; Hu, X.; Li, Y. The impact of environmental factors on the environmental bacterial diversity and composition in the Jiang-flavoured baijiu production region. LWT 2021, 149, 111784. [Google Scholar] [CrossRef]
- Chen, J.; Pang, H.; Wang, L.; Ma, C.; Wu, G.; Liu, Y.; Guan, Y.; Zhang, M.; Qin, G.; Tan, Z. Bacteriocin-producing lactic acid bacteria strains with antimicrobial activity screened from Bamei pig feces. Foods 2022, 11, 709. [Google Scholar] [CrossRef]
- Hao, F.; Tan, Y.; Lv, X.; Chen, L.; Yang, F.; Wang, H.; Du, H.; Wang, L.; Xu, Y. Microbial community succession and its environment driving factors during initial fermentation of Maotai-flavor baijiu. Front. Microbiol. 2021, 12, 669201. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Y.H.; Lin, J.Y.; Liu, L.S.; Huang, Z.X. Analysis of microbial diversity in the fermented grains of Maotai-flavor liquor using high-throughput sequencing. J. Fujian Norm. Univ. 2017, 33, 51–59. [Google Scholar]
- Song, Z.; Du, H.; Zhang, Y.; Xu, Y. Unraveling core functional microbiota in traditional solid-state fermentation by high-throughput amplicons and metatranscriptomics sequencing. Front. Microbiol. 2017, 8, 1294. [Google Scholar] [CrossRef]
- Wang, S.; Wu, Q.; Nie, Y.; Wu, J.; Xu, Y. Construction of synthetic microbiota for reproducible flavor compound metabolism in Chinese light-aroma-type liquor produced by solid-state fermentation. Appl. Environ. Microbiol. 2019, 85, e03090-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, R.; Wu, Q.; Xu, Y. Chinese liquor fermentation: Identification of key flavor-producing Lactobacillus spp. by quantitative profiling with indigenous internal standards. Appl. Environ. Microbiol. 2020, 86, e00456-20. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, T.; Jin, M.; Tan, Y.; Du, P. Metabolomics analysis of growth inhibition of Lactobacillus plantarum under ethanol stress. Int. J. Food. Sci. Tech. 2020, 55, 3441–3454. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, Y.; Chen, Q.; Zhao, L.; Zhou, J. Isolation and culture of ethanol resistant Lactobacillus. Food Ind. 2018, 39, 114–119. [Google Scholar]
- Pradhan, P.; Tamang, J. Probiotic properties of lactic acid bacteria isolated from traditionally prepared dry starters of the Eastern Himalayas. World J. Microbiol. Biotechnol. 2021, 37, 7. [Google Scholar] [CrossRef]
- Bravo-Ferrada, B.; Hollmann, A.; Delfederico, L.; Valdés La Hens, D.; Caballero, A.; Semorile, L. Patagonian red wines: Selection of Lactobacillus plantarum isolates as potential starter cultures for malolactic fermentation. World J. Microbiol. Biotechnol. 2013, 29, 1537–1549. [Google Scholar] [CrossRef]
- Nawaz, A.; Jagadeesh, K.S.; Krishnaraj, P.U. Isolation and screening of lactic acid bacteria for acidic pH and bile tolerance. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 3975–3980. [Google Scholar] [CrossRef]
- Arellano, K.; Vazquez, J.; Park, H.; Lim, J.; Ji, Y.; Kang, H.; Cho, D.; Jeong, H.; Holzapfel, W. Safety evaluation and whole-genome annotation of Lactobacillus plantarum strains from different sources with special focus on isolates from green tea. Probiotics. Antimicrob. Proteins. 2020, 12, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Deen, B.; Diez-Gonzalez, F. Assessment of Pediococcus acidilactici ATCC 8042 as potential Salmonella surrogate for thermal treatments of toasted oats cereal and peanut butter. Food. Microbiol. 2019, 83, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Bansal, P.; Kumar, R.; Singh, J.; Dhanda, S. Next generation sequencing, biochemical characterization, metabolic pathway analysis of novel probiotic Pediococcus acidilactici NCDC 252 and it’s evolutionary relationship with other lactic acid bacteria. Mol. Biol. Rep. 2019, 46, 5883–5895. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
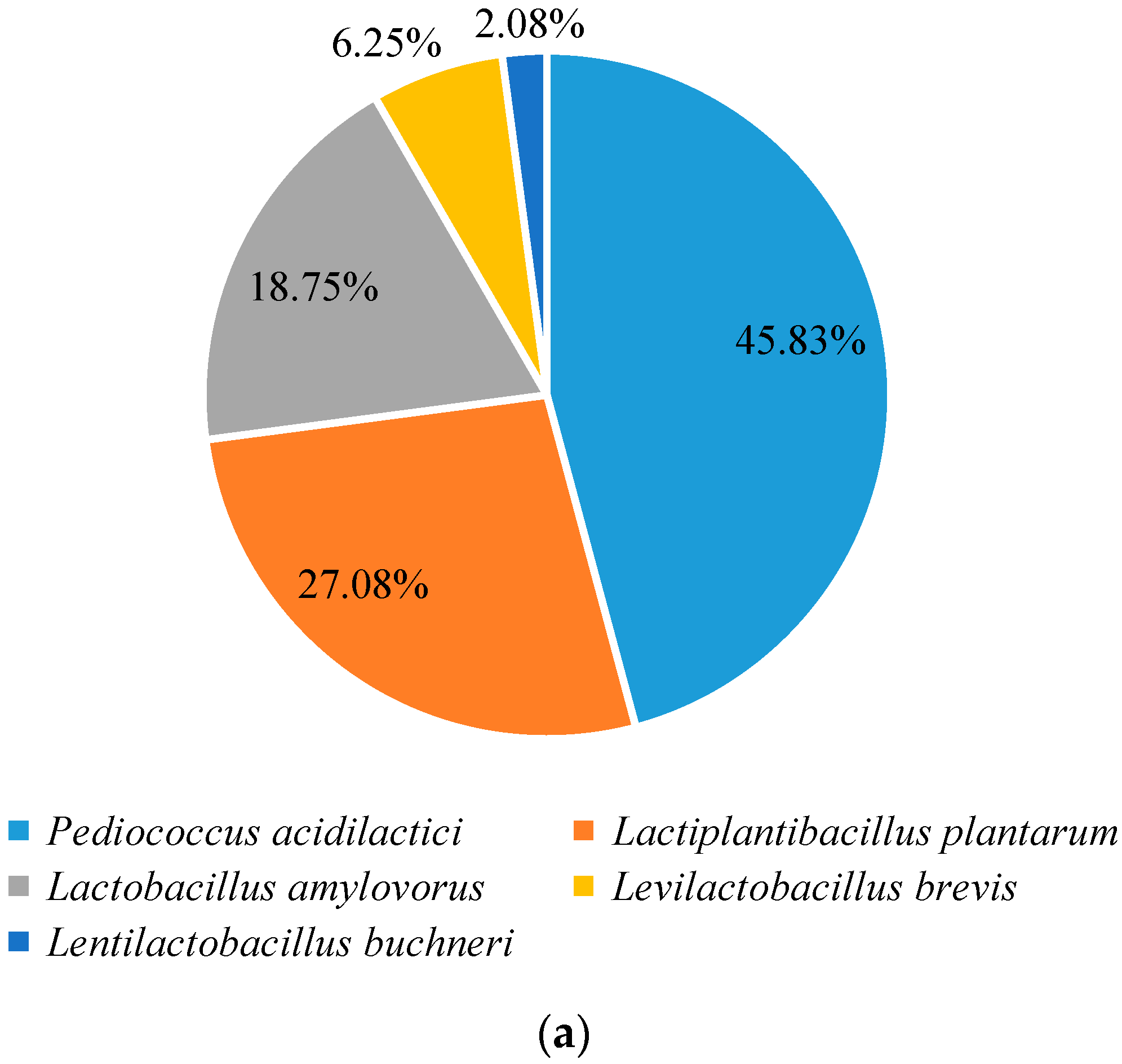{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Points | Samples | Shannon | Simpson | Ace | Chao |
|---|---|---|---|---|---|
| The first round out of the cellar (LTJP1C) | LTJP1C_1 | 1.67 | 0.28 | 61.89 | 57.50 |
| LTJP1C_2 | 1.47 | 0.35 | 58.43 | 48.75 | |
| LTJP1C_3 | 1.64 | 0.31 | 49.63 | 48.00 | |
| Average | 1.59 | 0.31 | 56.65 | 51.42 | |
| The fourth round out of the cellar (LTJP4C) | LTJP4C_1 | 1.51 | 0.38 | 56.26 | 45.20 |
| LTJP4C_2 | 1.35 | 0.45 | 56.02 | 50.17 | |
| LTJP4C_3 | 0.99 | 0.62 | 69.49 | 68.33 | |
| Average | 1.28 | 0.48 | 60.59 | 54.57 | |
| The seventh round out of the cellar (LTJP7C) | LTJP7C_1 | 1.70 | 0.33 | 81.98 | 80.60 |
| LTJP7C_2 | 1.70 | 0.30 | 61.12 | 45.00 | |
| LTJP7C_3 | 2.10 | 0.15 | 62.77 | 58.00 | |
| Average | 1.83 | 0.26 | 68.62 | 61.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Lu, C.; Xu, Q.; Li, Z.; Song, Y.; Zhou, S.; Zhang, T.; Luo, X. Bacterial Diversity and Lactic Acid Bacteria with High Alcohol Tolerance in the Fermented Grains of Soy Sauce Aroma Type Baijiu in North China. Foods 2022, 11, 1794. https://doi.org/10.3390/foods11121794
Wang J, Lu C, Xu Q, Li Z, Song Y, Zhou S, Zhang T, Luo X. Bacterial Diversity and Lactic Acid Bacteria with High Alcohol Tolerance in the Fermented Grains of Soy Sauce Aroma Type Baijiu in North China. Foods. 2022; 11(12):1794. https://doi.org/10.3390/foods11121794
Chicago/Turabian StyleWang, Jiali, Chengshun Lu, Qiang Xu, Zhongyuan Li, Yajian Song, Sa Zhou, Tongcun Zhang, and Xuegang Luo. 2022. "Bacterial Diversity and Lactic Acid Bacteria with High Alcohol Tolerance in the Fermented Grains of Soy Sauce Aroma Type Baijiu in North China" Foods 11, no. 12: 1794. https://doi.org/10.3390/foods11121794