
The Investigation of Protein Profile and Meat Quality in Bovine Longissimus thoracic Frozen under Different Temperatures by Data-Independent Acquisition (DIA) Strategy

Abstract
:
1. Introduction
2. Materials and Methods
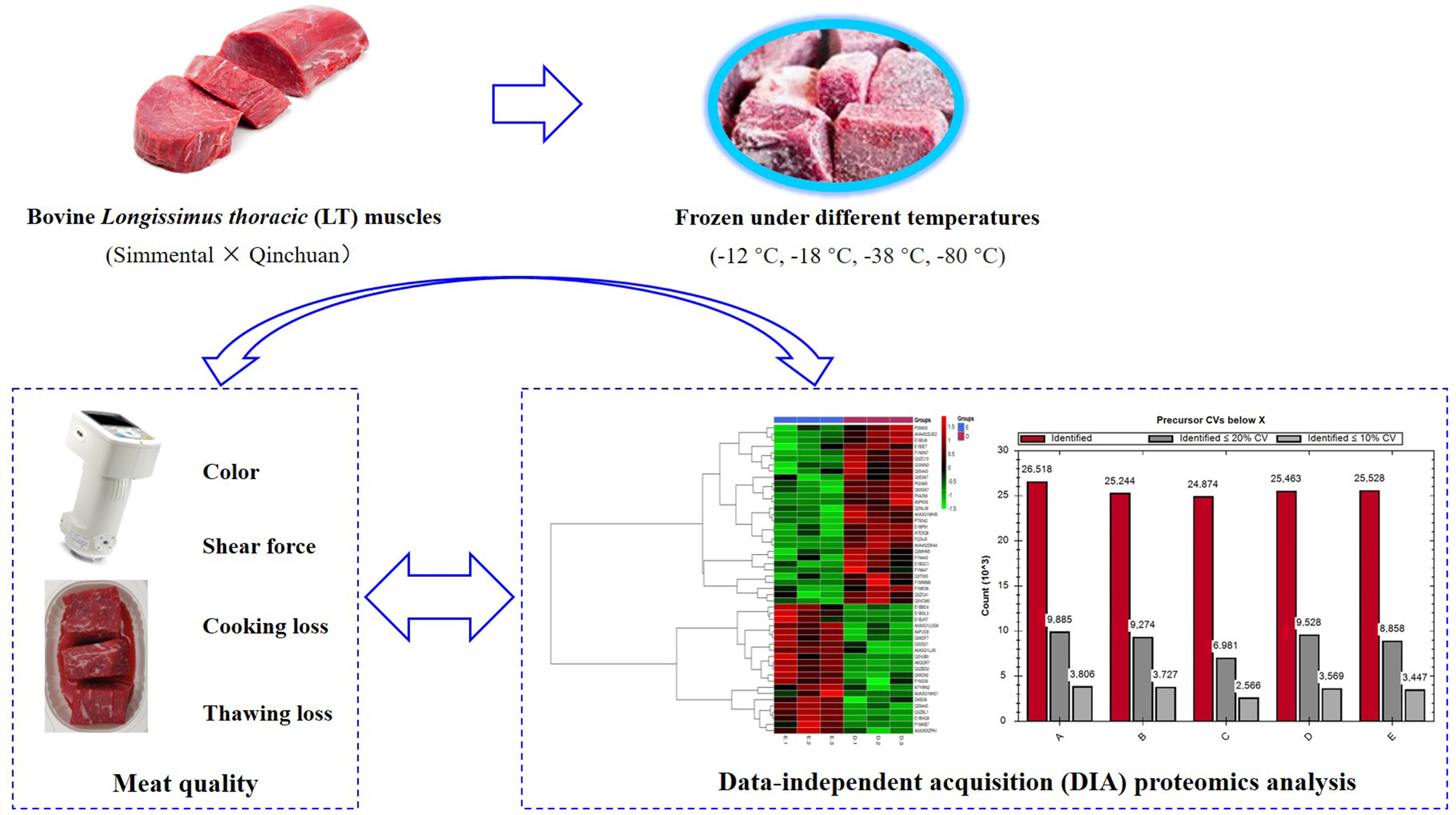
2.1. Sample Collection and Preparation
2.2. Meat Quality Determination
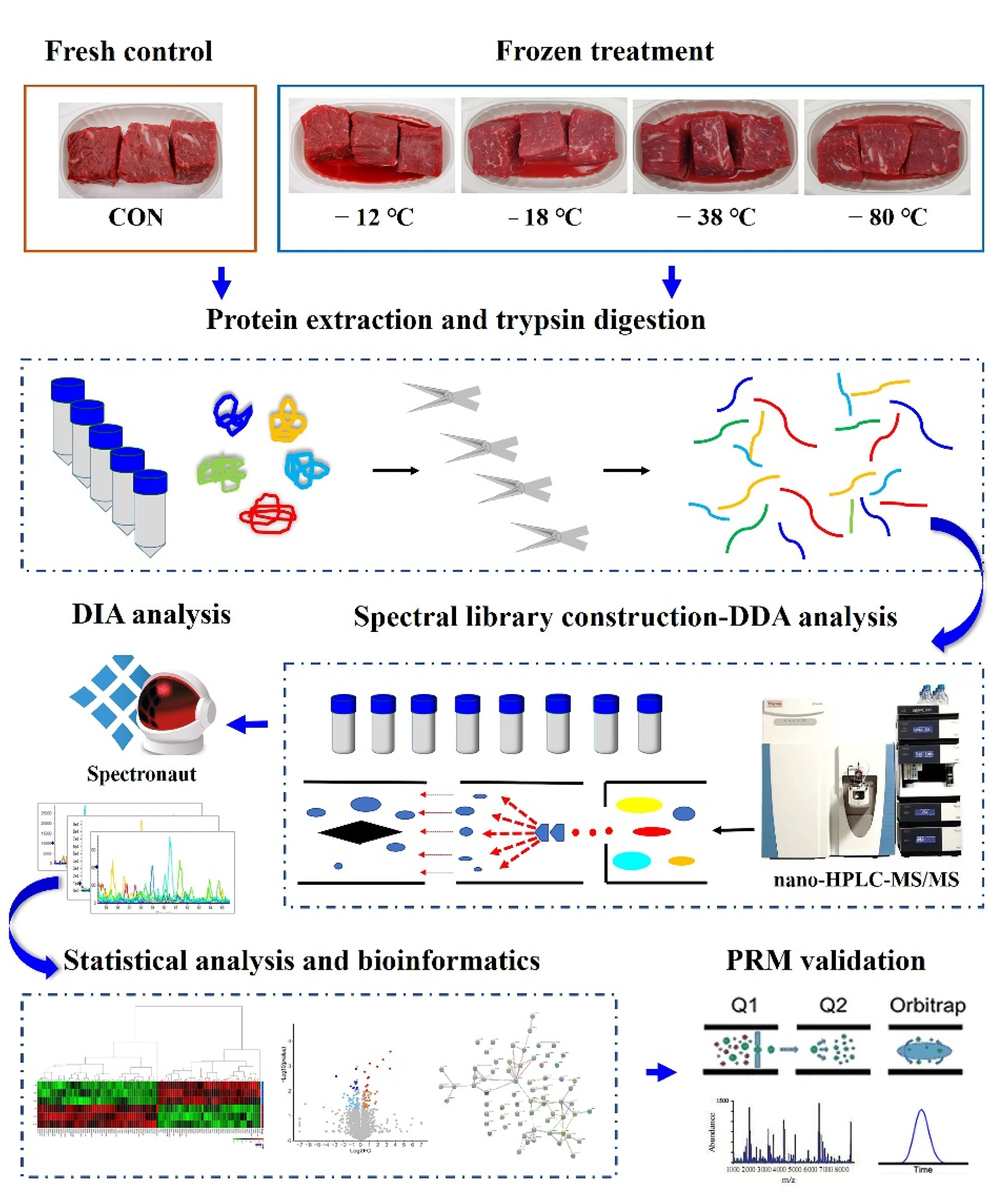
2.3. Protein Extraction and Digestion
2.4. Spectral Library Generation
2.4.1. High-pH Reversed-Phase Separation
2.4.2. Nano-HPLC-MS/MS Analysis for DDA and DIA Analysis
2.4.3. Database Search and Analysis
2.5. Parallel Reaction Monitoring (PRM)
2.6. Bioinformatics and Statistical Analysis
3. Results and Discussion
3.1. Meat Quality of the Beef Samples
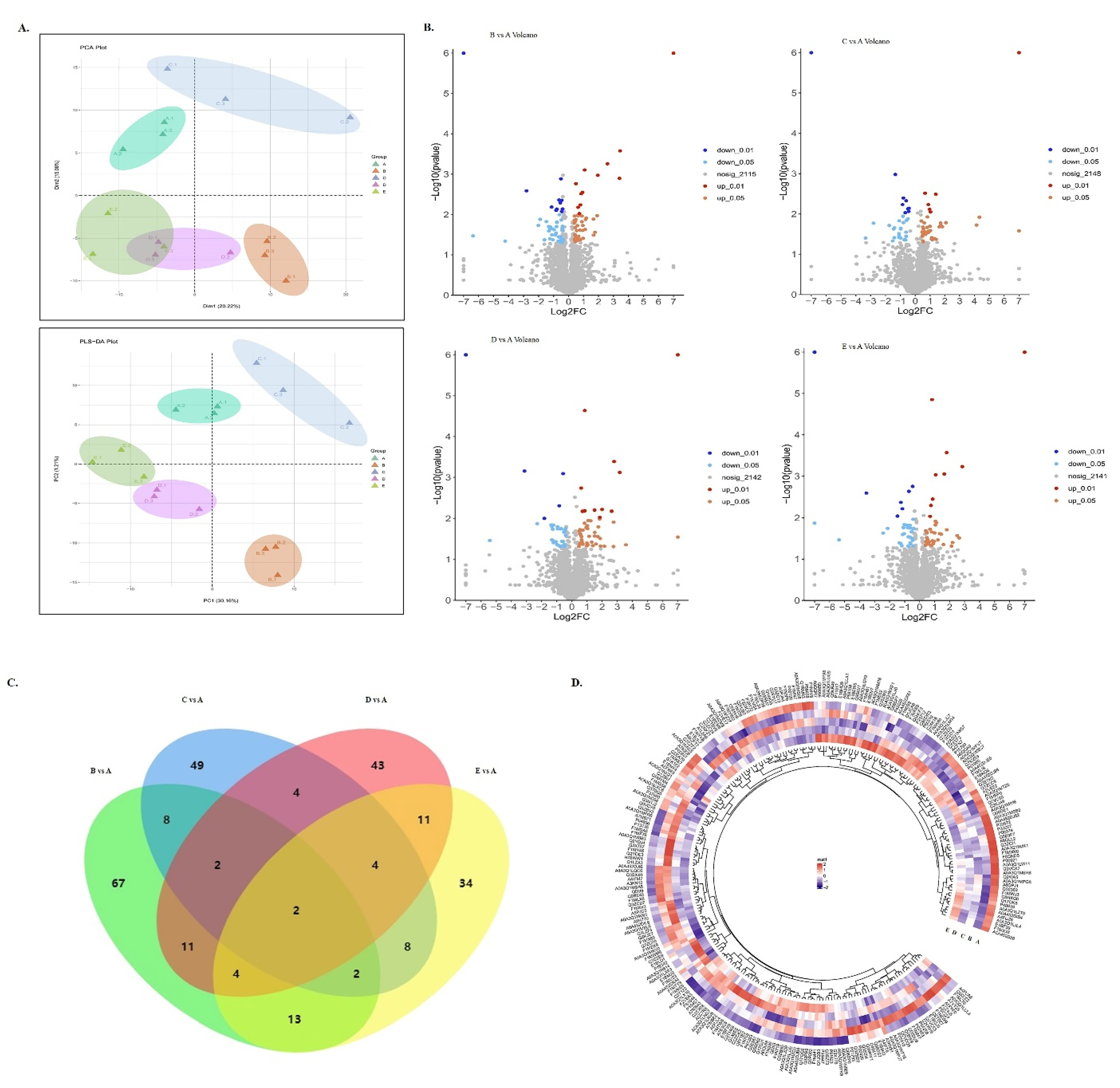
3.2. Protein Profiling in Fresh and Frozen Samples
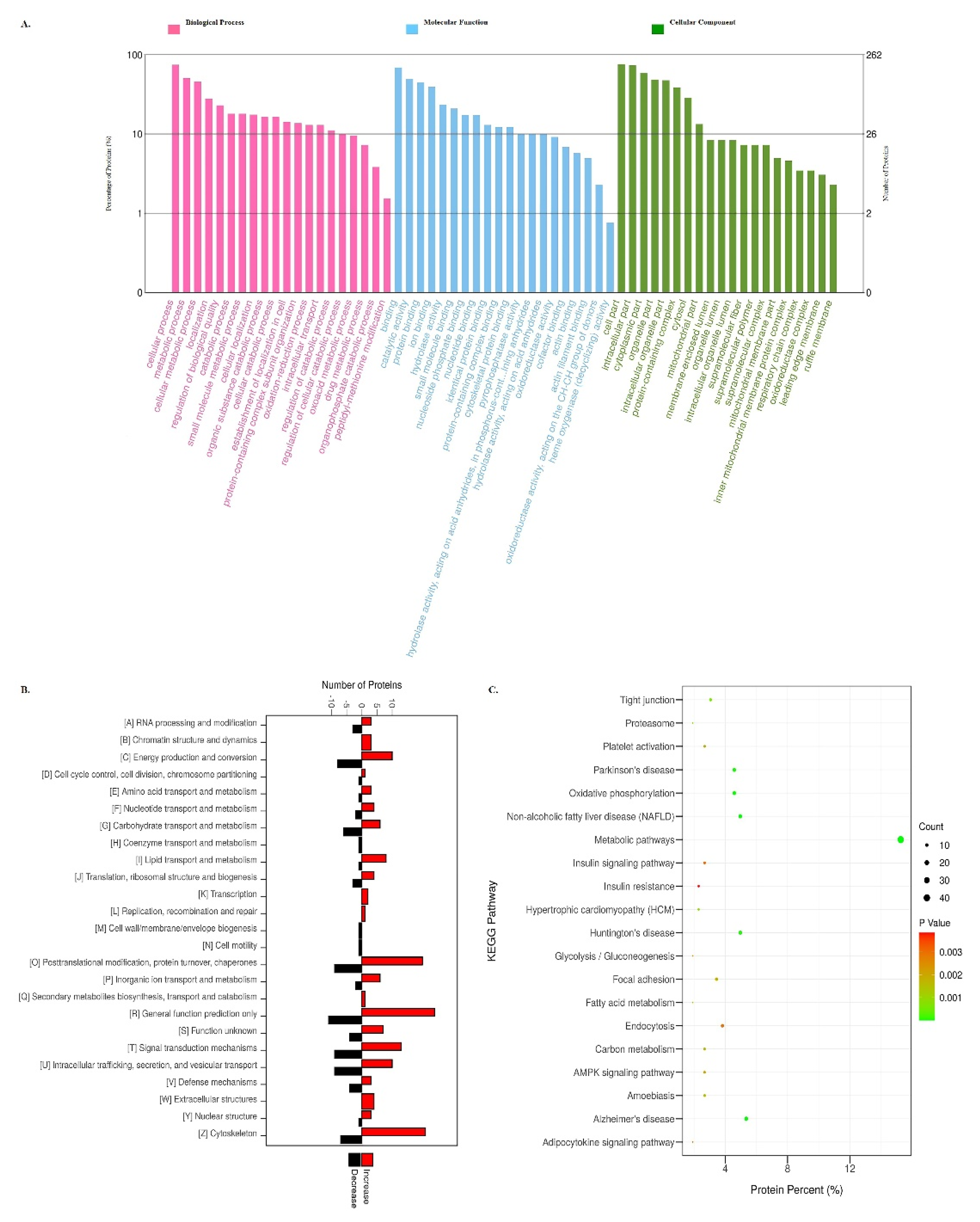
3.3. GO, KOG, and KEGG Pathway Analysis of DAPs
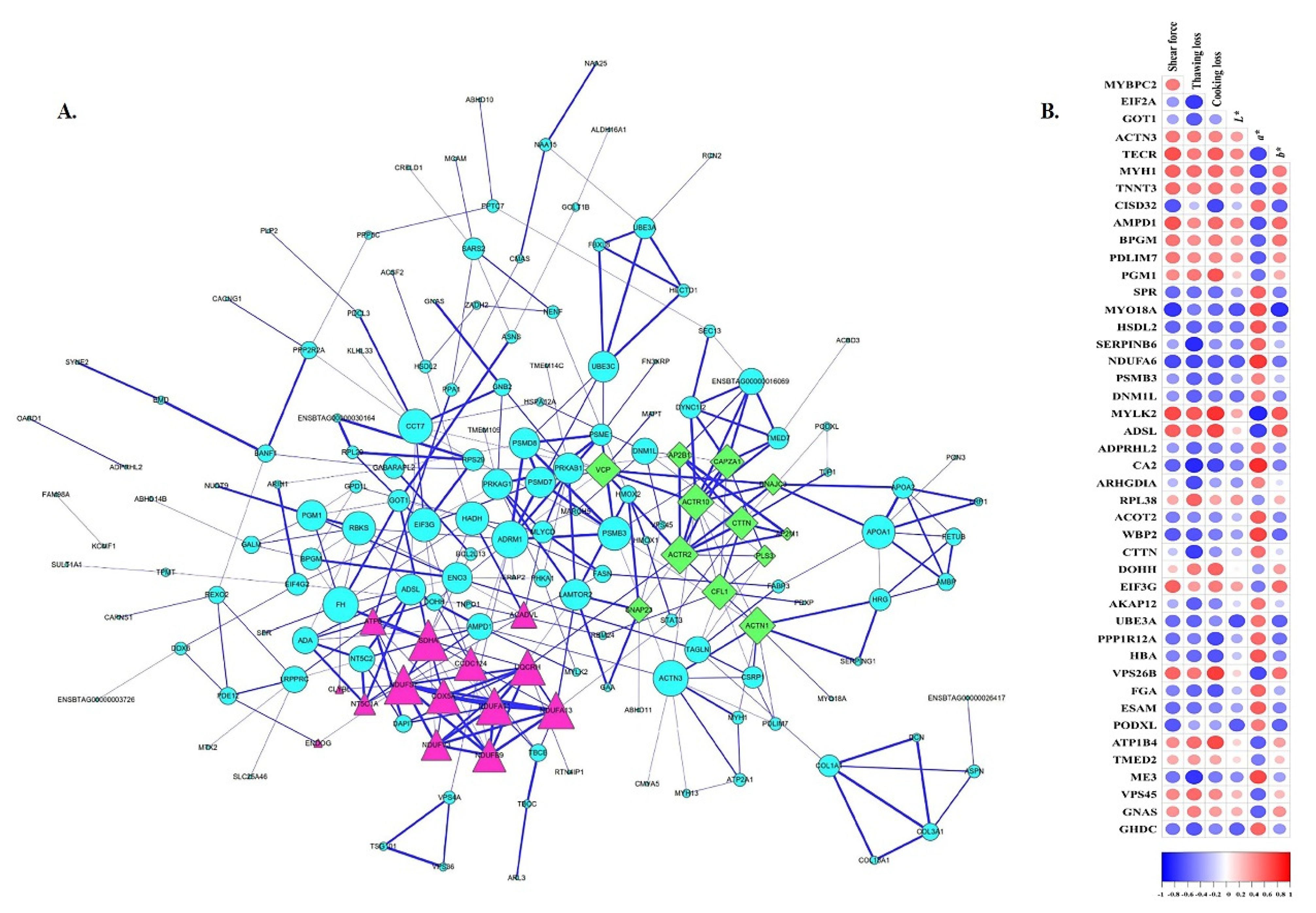
3.4. Protein–Protein Interaction Analysis
3.5. Correlations between DAPs and Meat Quality
3.5.1. Structure Proteins
3.5.2. Enzymes
3.5.3. Other Proteins
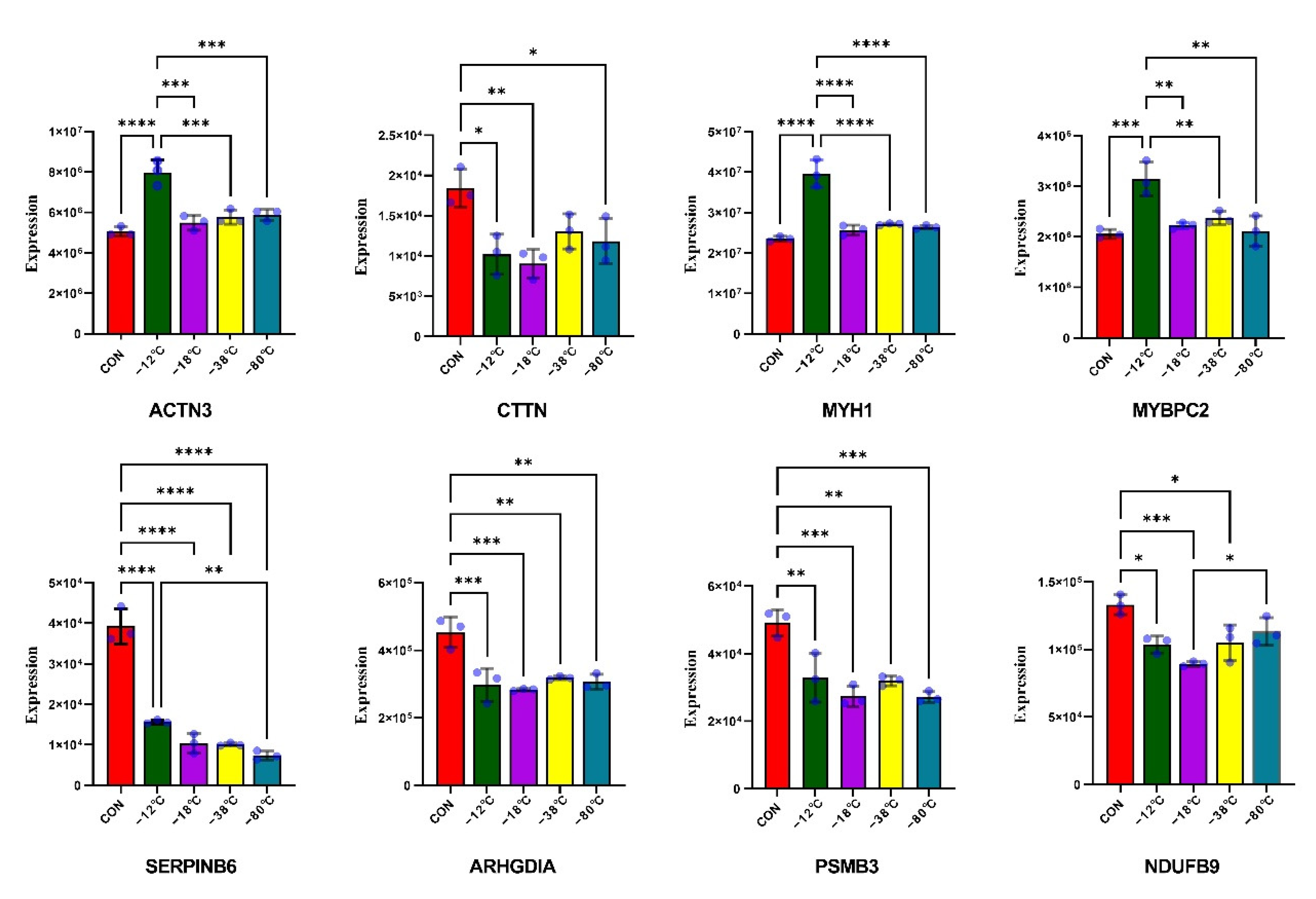
3.6. Validation of DAPs by PRM
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, S.B.; Gotoh, T.; Greenwood, P.L. Current situation and future prospects for global beef production: Overview of special issue. Asian Australas. J. Anim. Sci. 2018, 31, 927–932. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.B.; Chen, Y.F.; Zhao, J.; Wu, B.B. Impacts of household income on beef at-home consumption: Evidence from urban China. J. Integr. Agric. 2021, 20, 1701–1715. [Google Scholar] [CrossRef]
- OECD and Food and Agriculture Organization of the United Nations. OECD-FAO Agricultural Outlook 2019–2028; OECD: Paris, France, 2019; pp. 12–20. [Google Scholar]
- Kim, H.W.; Kim, J.H.; Seo, J.K.; Setyabrata, D.; Kim, Y.H.B. Effects of aging/freezing sequence and freezing rate on meat quality and oxidative stability of pork loins. Meat Sci. 2018, 139, 162–170. [Google Scholar] [CrossRef]
- Gao, S.J.; Wang, R.G.; Bi, Y.L.; Qu, H.; Chen, Y.; Zheng, L. Identification of frozen/thawed beef based on label-free detection of hemin (Iron Porphyrin) with solution-gated graphene transistor sensors. Sens. Actuators B Chem. 2020, 305, 127167. [Google Scholar] [CrossRef]
- Leygonie, C.; Britz, T.J.; Hoffman, L.C. Impact of freezing and thawing on the quality of meat: Review. Meat Sci. 2012, 91, 93–98. [Google Scholar] [CrossRef]
- Lagerstedt, A.; Enfalt, L.; Johansson, L.; Lundstrom, K. Effect of freezing on sensory quality, shear force and water loss in beef M. longissimus dorsi. Meat Sci. 2008, 80, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, E.J.; Park, D.H.; Ji, Y.R.; Kang, G.; Choi, M.J. Deep freezing to maintain the freshness of pork loin during long-term storage. Food Sci. Biotechnol. 2021, 30, 701–710. [Google Scholar] [CrossRef]
- Wang, F.; Liang, R.; Zhang, Y.; Gao, S.; Zhu, L.; Niu, L.; Hopkins, D.L. Effects of packaging methods combined with frozen temperature on the color of frozen beef rolls. Meat Sci. 2021, 171, 108292. [Google Scholar] [CrossRef]
- Holman, B.W.B.; Coombs, C.E.O.; Morris, S.; Kerr, M.J.; Hopkins, D.L. Effect of long term chilled (up to 5weeks) then frozen (up to 12months) storage at two different sub-zero holding temperatures on beef: 1. Meat quality and microbial loads. Meat Sci. 2017, 133, 133–142. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Yan, J.K.; Ding, Y.H.; Ma, H.L. Effects of ultrasound on the thawing of quick-frozen small yellow croaker (Larimichthys polyactis) based on TMT-labeled quantitative proteomic. Food Chem. 2022, 366, 130600. [Google Scholar] [CrossRef]
- Yu, Q.Q.; Wu, W.; Tian, X.J.; Hou, M.; Dai, R.T.; Li, X.M. Unraveling proteome changes of Holstein beef M. semitendinosus and its relationship to meat discoloration during post-mortem storage analyzed by label-free mass spectrometry. J. Proteom. 2017, 154, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Picard, B.; Gagaoua, M.; Micol, D.; Cassar-Malek, I.; Hocquette, J.F.; Terlouw, C.E. Inverse relationships between biomarkers and beef tenderness according to contractile and metabolic properties of the muscle. J. Agric. Food Chem. 2014, 62, 9808–9818. [Google Scholar] [CrossRef] [PubMed]
- Picard, B.; Gagaoua, M. Meta-proteomics for the discovery of protein biomarkers of beef tenderness: An overview of integrated studies. Food Res. Int. 2020, 127, 108739. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.H.; Liu, Q.F.; Meng, Q.S.; Wang, L.G.; Yan, H.; Zhang, L.C.; Wang, L.X. TMT-based quantitative proteomic analysis of porcine muscle associated with postmortem meat quality. Food Chem. 2020, 328, 127133. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Huang, F.; Li, X.; Zhang, H.R.; Liu, J.Q.; Han, D.; Zhang, C.H. DIA-based quantitative proteomic analysis on the meat quality of porcine Longissimus thoracis et lumborum cooked by different procedures. Food Chem. 2022, 371, 131206. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; He, F.; Rao, W.L.; Ni, N.; Shen, Q.W.; Zhang, D.Q. Proteomic analysis of goat Longissimus dorsi muscles with different drip loss values related to meat quality traits. Food Sci. Biotechnol. 2016, 25, 425–431. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, L.T.; Lei, Y.T.; Shen, H.X.; Yu, X.P.; Luo, Y.K. Differential proteomic analysis to identify proteins associated with quality traits of frozen mud shrimp (Solenocera melantho) using an iTRAQ-based strategy. Food Chem. 2018, 251, 25–32. [Google Scholar] [CrossRef]
- Wang, C.; Chu, J.J.; Fu, L.L.; Wang, Y.B.; Zhao, F.; Zhou, D.Q. iTRAQ-based quantitative proteomics reveals the biochemical mechanism of cold stress adaption of razor clam during controlled freezing-point storage. Food Chem. 2018, 247, 73–80. [Google Scholar] [CrossRef]
- Jia, W.; Zhang, R.; Liu, L.; Zhu, Z.B.; Xu, M.D.; Shi, L. Molecular mechanism of protein dynamic change for Hengshan goat meat during freezing storage based on high-throughput proteomics. Food Res. Int. 2021, 143, 110289. [Google Scholar] [CrossRef]
- Kim, G.D.; Jeong, T.C.; Yang, H.S.; Joo, S.T.; Hur, S.J.; Jeong, J.Y. Proteomic analysis of meat exudates to discriminate fresh and freeze-thawed porcine longissimus thoracis muscle. LWT Food Sci. Technol. 2015, 62, 1235–1238. [Google Scholar] [CrossRef]
- Lin, L.; Zheng, J.X.; Yu, Q.; Chen, W.D.; Xing, J.C.; Chen, C.X.; Tian, R.J. High throughput and accurate serum proteome profiling by integrated sample preparation technology and single-run data independent mass spectrometry analysis. J. Proteom. 2018, 174, 9–16. [Google Scholar] [CrossRef]
- Malmstrom, L.; Bakochi, A.; Svensson, G.; Kilsgard, O.; Lantz, H.; Petersson, A.C.; Malmstrom, J. Quantitative proteogenomics of human pathogens using DIA-MS. J. Proteom. 2015, 129, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Qian, S.Y.; Li, X.; Wang, H.; Mehmood, W.; Zhong, M.; Zhang, C.H.; Blecker, C. Effects of low voltage electrostatic field thawing on the changes in physicochemical properties of myofibrillar proteins of bovine Longissimus dorsi muscle. J. Food Eng. 2019, 261, 140–149. [Google Scholar] [CrossRef]
- CIE. Colorimetry, 2nd ed.; Centre International de L’Eclairage: Vienna, Austria, 1986; Volume 15.2. [Google Scholar]
- Qian, S.Y.; Hu, F.F.; Mehmood, W.; Li, X.; Zhang, C.H.; Blecker, C. The rise of thawing drip: Freezing rate effects on ice crystallization and myowater dynamics changes. Food Chem. 2022, 373, 131461. [Google Scholar] [CrossRef]
- Muela, E.; Monge, P.; Sanudo, C.; Campo, M.M.; Beltran, J.A. Meat quality of lamb frozen stored up to 21 months: Instrumental analyses on thawed meat during display. Meat Sci. 2015, 102, 35–40. [Google Scholar] [CrossRef]
- Coluccio, L.M. Myosins: A Superfamily of Molecular Motors, 2nd ed.; Springer Nature: Gewerbestrasse, Switzerland, 2020; pp. 1, 284–285, 422–430. [Google Scholar]
- Wang, H.N.; Wu, J.P.; Betti, M. Chemical, rheological and surface morphologic characterisation of spent hen proteins extracted by pH-shift processing with or without the presence of cryoprotectants. Food Chem. 2013, 139, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Gagaoua, M.; Hughes, J.; Terlouw, E.M.C.; Warner, R.D.; Purslow, P.P.; Lorenzo, J.M.; Picard, B. Proteomic biomarkers of beef colour. Trends Food Sci. Tech. 2020, 101, 234–252. [Google Scholar] [CrossRef]
- Liu, Y.B.; Chu, A.; Chakroun, I.; Islam, U.; Blais, A. Cooperation between myogenic regulatory factors and SIX family transcription factors is important for myoblast differentiation. Nucleic Acids Res. 2010, 38, 6857–6871. [Google Scholar] [CrossRef] [Green Version]
- Boudon, S.; Henry-Berger, J.; Cassar-Malek, I. Aggregation of Omic Data and Secretome Prediction Enable the Discovery of Candidate Plasma Biomarkers for Beef Tenderness. Int. J. Mol. Sci. 2020, 21, 664. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.H.P.; Rodrigues, R.T.S.; Assis, D.E.F.; Benedeti, P.D.B.; Duarte, M.S.; Chizzotti, M.L. Explaining meat quality of bulls and steers by differential proteome and phosphoproteome analysis of skeletal muscle. J. Proteom. 2019, 199, 51–66. [Google Scholar] [CrossRef]
- Yan, Y.; Zhou, X.E.; Xu, H.E.; Melcher, K. Structure and Physiological Regulation of AMPK. Int. J. Mol. Sci. 2018, 19, 3534. [Google Scholar] [CrossRef] [Green Version]
- Franco, D.; Mato, A.; Salgado, F.J.; Lopez-Pedrouso, M.; Carrera, M.; Bravo, S.; Zapata, C. Tackling proteome changes in the longissimus thoracis bovine muscle in response to pre-slaughter stress. J. Proteom. 2015, 122, 73–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagaoua, M.; Bonnet, M.; De Koning, L.; Picard, B. Reverse Phase Protein array for the quantification and validation of protein biomarkers of beef qualities: The case of meat color from Charolais breed. Meat Sci. 2018, 145, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Men, L.; Li, Y.Z.; Wang, X.L.; Li, R.J.; Zhang, T.; Meng, X.S.; Liu, S.C.; Gong, X.J.; Gou, M. Protein biomarkers associated with frozen Japanese puffer fish (Takifugu rubripes) quality traits. Food Chem. 2020, 327, 127002. [Google Scholar] [CrossRef] [PubMed]
- Faustman, C.; Sun, Q.; Mancini, R.; Suman, S.P. Myoglobin and lipid oxidation interactions: Mechanistic bases and control. Meat Sci. 2010, 86, 86–94. [Google Scholar] [CrossRef]
- Weng, K.Q.; Huo, W.R.; Gu, T.T.; Bao, Q.; Cao, Z.F.; Zhang, Y.; Zhang, Y.; Xu, Q.; Chen, G.H. Quantitative phosphoproteomic analysis unveil the effect of marketable ages on meat quality in geese. Food Chem. 2021, 361, 130093. [Google Scholar] [CrossRef]
- Gill, J.L.; Bishop, S.C.; McCorquodale, C.; Williams, J.L.; Wiener, P. Identification of polymorphisms in the malic enzyme 1, NADP(+)-dependent, cytosolic and nuclear receptor subfamily 0, group B, member 2 genes and their associations with meat and carcass quality traits in commercial Angus cattle. Anim. Genet. 2012, 43, 88–92. [Google Scholar] [CrossRef]
- Lopez-Pedrouso, M.; Franco, D.; Serrano, M.P.; Maggiolino, A.; Landete-Castillejos, T.; De Palo, P.; Lorenzo, J.M. A proteomic-based approach for the search of biomarkers in Iberian wild deer (Cervus elaphus) as indicators of meat quality. J. Proteom. 2019, 205, 103422. [Google Scholar] [CrossRef]
- Shen, Y.N.; Kim, S.H.; Yoon, D.H.; Lee, H.G.; Kang, H.S.; Seo, K.S. Proteome Analysis of Bovine Longissimus dorsi Muscle Associated with the Marbling Score. Asian Australas. J. Anim. Sci. 2012, 25, 1083–1088. [Google Scholar] [CrossRef]
- Xia, X.T.; Zhang, S.J.; Zhang, H.J.; Zhang, Z.J.; Li, Z.G.; Sun, H.X.; Liu, X.; Lyu, S.J.; Wang, X.W.; Li, Z.M.; et al. Assessing genomic diversity and signatures of selection in Jiaxian Red cattle using whole-genome sequencing data. BMC Genom. 2021, 22, 43. [Google Scholar] [CrossRef]
- Kimball, S.R. Eukaryotic initiation factor eif2. Int J Biochem Cell Biol. 1999, 31, 25–29. [Google Scholar] [CrossRef]
- Mccollum, A.K.; Angelos, M.G.; Fischione, A.D.; Mineo, M.; Kohn, E.C. Abstract 2032: A novel function of ww domain binding protein 2 (wbp2) in regulating cytoskeletal function and cellular division through binding to co-chaperone bag3. Cancer Res. 2012, 72 (Suppl. S8), 2032. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attribute | CON | −12 °C | −18 °C | −38 °C | −80 °C | p-Value |
|---|---|---|---|---|---|---|
| L* | 38.05 ± 0.51 b | 40.06 ± 0.70 a | 39.59 ± 1.81 ab | 39.17 ± 0.90 ab | 39.04 ± 0.79 ab | 0.034 |
| a* | 15.38 ± 0.71 a | 12.94 ± 0.34 c | 13.40 ± 0.18 c | 14.42 ± 0.28 b | 14.47 ± 0.24 b | <0.001 |
| b* | 11.66 ± 1.38 c | 13.19 ± 0.23 a | 12.78 ± 0.39 ab | 12.03 ± 0.12 bc | 11.75 ± 0.18 bc | 0.001 |
| Thawing Loss/% | – | 10.50 ± 0.84 a | 8.83 ± 0.75 b | 7.67 ± 0.52 bc | 7.17 ± 0.7 c | <0.001 |
| Cooking Loss/% | 28.28 ± 2.12 c | 34.0 ± 26 a | 32.33 ± 2.34 ab | 30.33 ± 3.14 abc | 29.83 ± 172 bc | 0.001 |
| Shear Force/kg | 7.81 ± 0.69 b | 10.72 ± 0.52 a | 9.94 ± 0.64 a | 8.36 ± 0.57 b | 8.22 ± 0.58 b | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Qian, S.; Huang, F.; Li, K.; Song, Y.; Liu, J.; Guo, Y.; Zhang, C.; Blecker, C. The Investigation of Protein Profile and Meat Quality in Bovine Longissimus thoracic Frozen under Different Temperatures by Data-Independent Acquisition (DIA) Strategy. Foods 2022, 11, 1791. https://doi.org/10.3390/foods11121791
Li X, Qian S, Huang F, Li K, Song Y, Liu J, Guo Y, Zhang C, Blecker C. The Investigation of Protein Profile and Meat Quality in Bovine Longissimus thoracic Frozen under Different Temperatures by Data-Independent Acquisition (DIA) Strategy. Foods. 2022; 11(12):1791. https://doi.org/10.3390/foods11121791
Chicago/Turabian StyleLi, Xia, Shuyi Qian, Feng Huang, Kaimin Li, Yu Song, Jiqian Liu, Yujie Guo, Chunhui Zhang, and Christophe Blecker. 2022. "The Investigation of Protein Profile and Meat Quality in Bovine Longissimus thoracic Frozen under Different Temperatures by Data-Independent Acquisition (DIA) Strategy" Foods 11, no. 12: 1791. https://doi.org/10.3390/foods11121791