
Effect of Commercial Yeast Starter Cultures on Cabernet Sauvignon Wine Aroma Compounds and Microbiota

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Industrial Fermentation
2.2. Physicochemical Analysis
2.3. Microbiological Analysis
2.4. Volatile Compound Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Dynamics of Physicochemical Properties during Fermentation
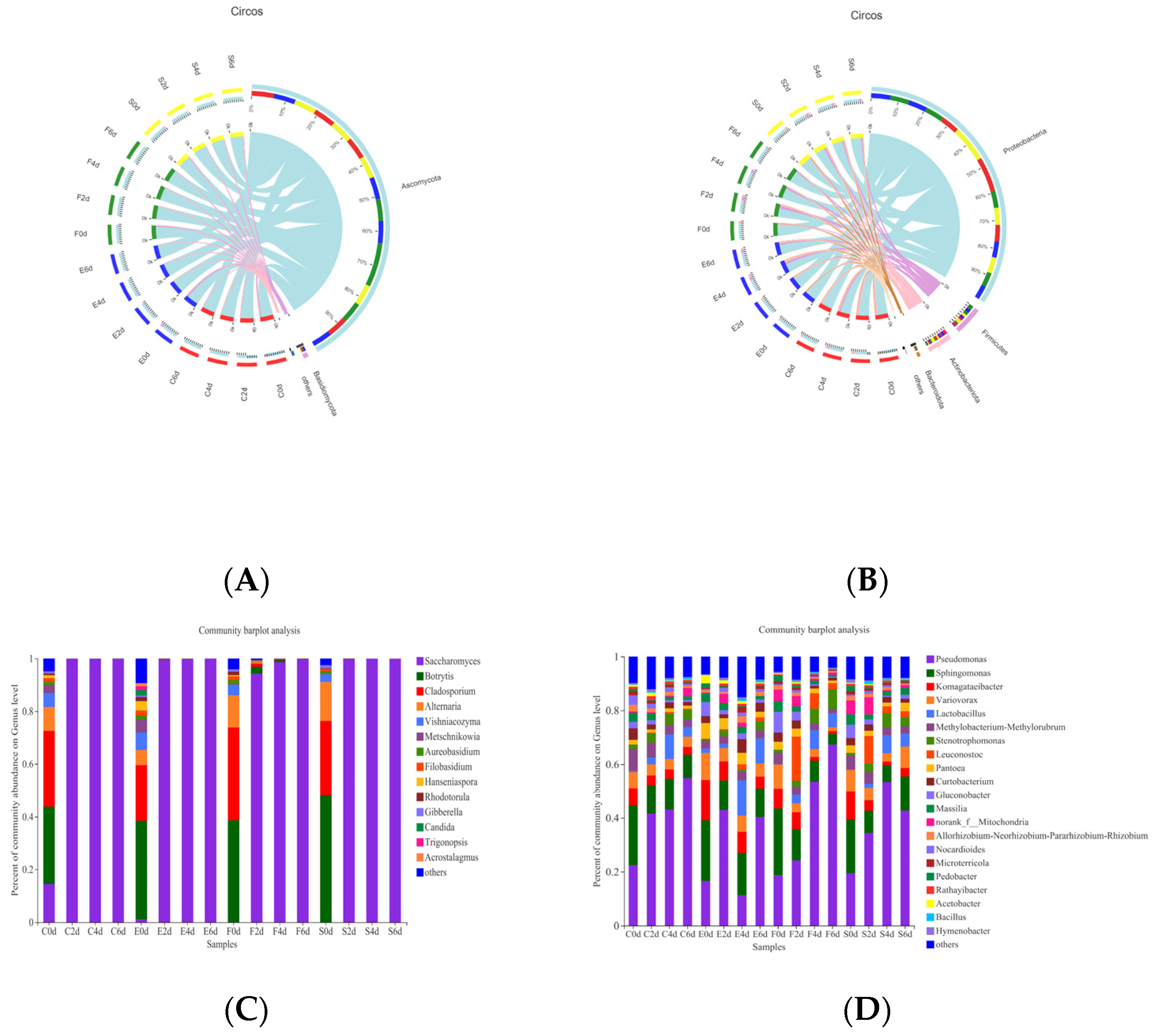
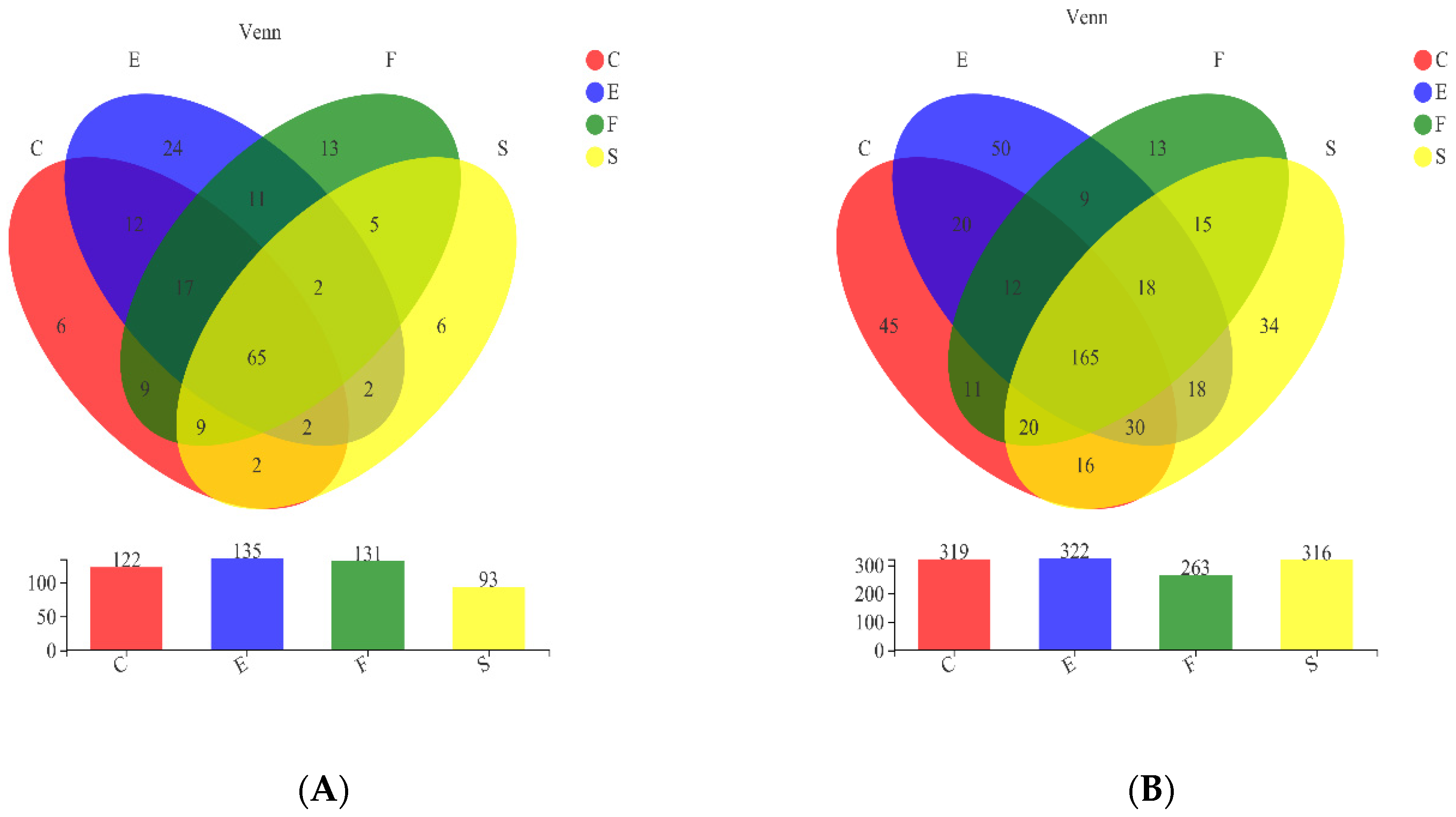
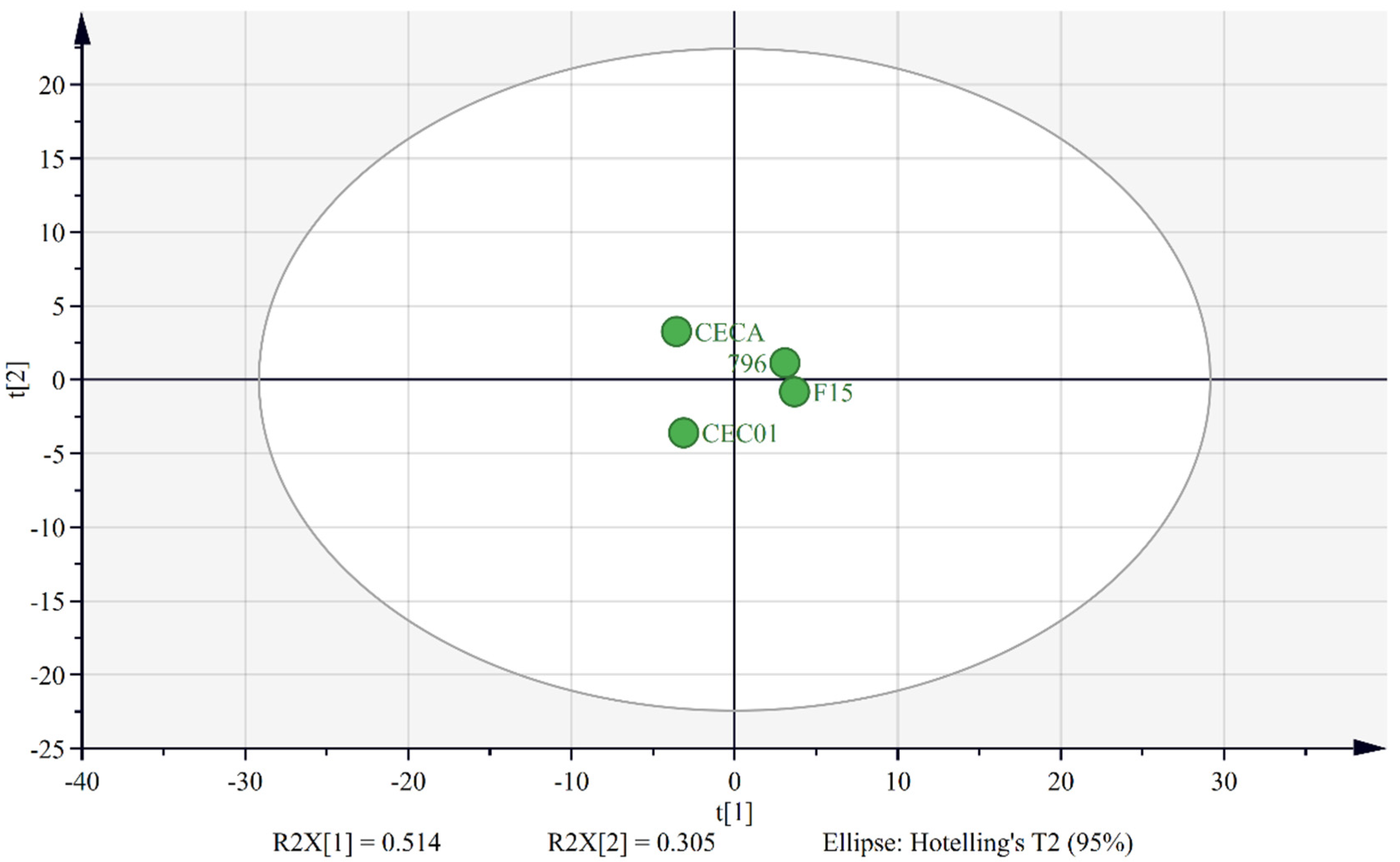
3.2. Comparison of Microbial Diversity among Samples
3.3. Change in Volatile Compounds during Fermentation
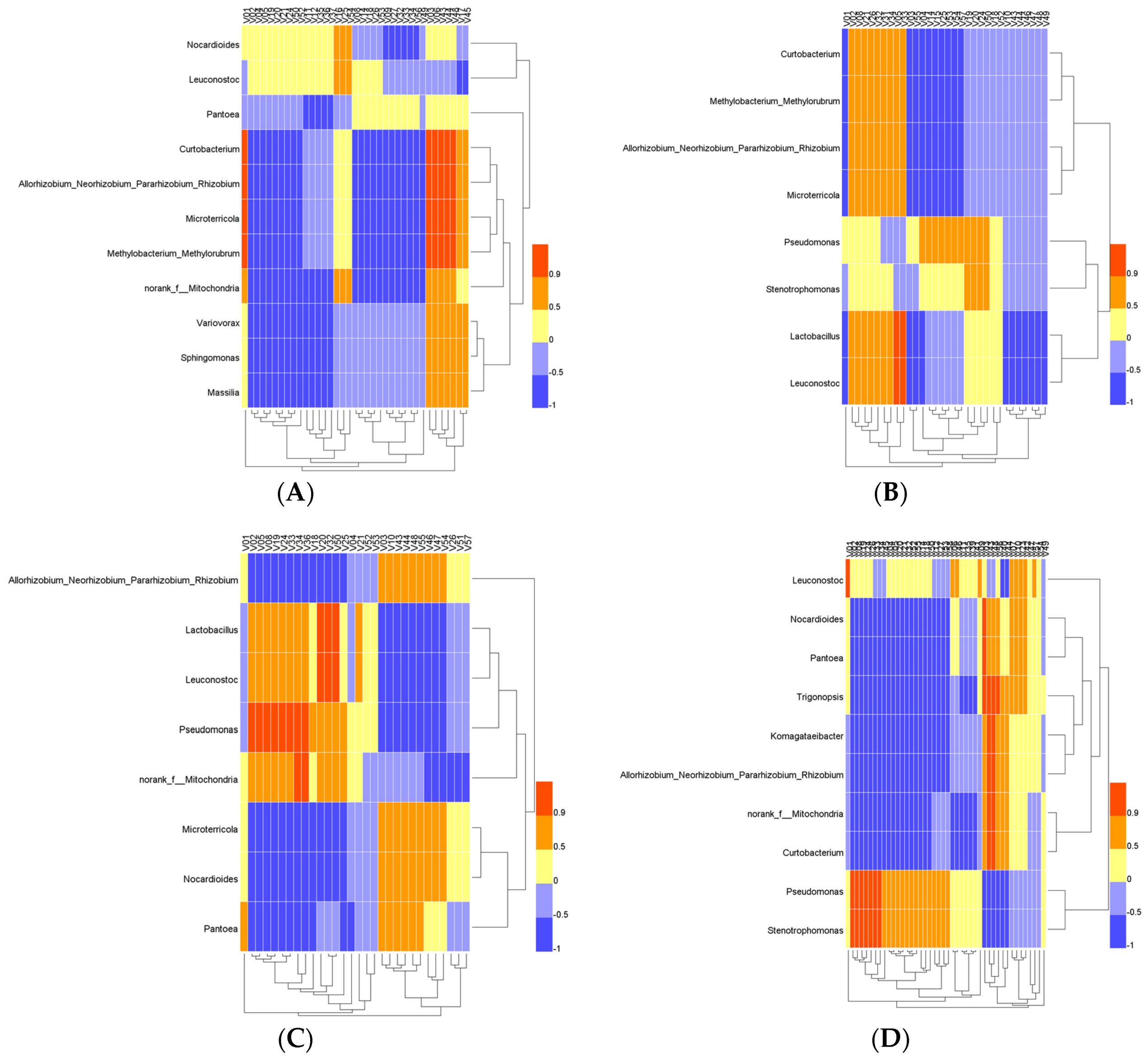
3.4. Correlation Analysis between Microbiota and Volatile Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- van Wyk, N.; Grossmann, M.; Wendland, J.; von Wallbrunn, C.; Pretorius, I.S. The Whiff of Wine Yeast Innovation: Strategies for Enhancing Aroma Production by Yeast during Wine Fermentation. J. Agric. Food Chem. 2019, 67, 13496–13505. [Google Scholar] [CrossRef]
- Molina, A.M.; Guadalupe, V.; Varela, C.; Swiegers, J.H.; Pretorius, I.S.; Agosin, E. Differential synthesis of fermentative aroma compounds of two related commercial wine yeast strains. Food Chem. 2009, 117, 189–195. [Google Scholar] [CrossRef]
- King, E.S.; Swiegers, J.H.; Travis, B.; Francis, I.L.; Bastian, S.E.P.; Pretorius, I.S. Coinoculated Fermentations Using Saccharomyces Yeasts Affect the Volatile Composition and Sensory Properties of Vitis vinifera L. cv. Sauvignon Blanc Wines. J. Agric. Food Chem. 2008, 56, 10829–10837. [Google Scholar] [CrossRef] [PubMed]
- King, E.S.; Kievit, R.L.; Curtin, C.; Swiegers, J.H.; Pretorius, I.S.; Bastian, S.E.P.; Francis, I.L. The effect of multiple yeasts co-inoculations on Sauvignon Blanc wine aroma composition, sensory properties and consumer preference. Food Chem. 2010, 122, 618–626. [Google Scholar] [CrossRef]
- Saberi, S.; Cliff, M.A.; van Vuuren, H.J.J. Impact of mixed S. cerevisiae strains on the production of volatiles and estimated sensory profiles of Chardonnay wines. Food Res. Int. 2012, 48, 725–735. [Google Scholar] [CrossRef]
- Stefanini, I.; Albanese, D.; Cavazza, A.; Franciosi, E.; De Filippo, C.; Donati, C.; Cavalieri, D. Dynamic changes in microbiota and mycobiota during spontaneous “Vino Santo Trentino” fermentation. Microb. Biotechnol. 2016, 9, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Chen, Q.; Zhang, P.; Chen, D.; Howell, K.S. The Fungal Microbiome Is an Important Component of Vineyard Ecosystems and Correlates with Regional Distinctiveness of Wine. Msphere 2020, 5, e00534. [Google Scholar] [CrossRef]
- Gonzalez-Barreiro, C.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gandara, J. Wine Aroma Compounds in Grapes: A Critical Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 202–218. [Google Scholar] [CrossRef]
- Vararu, F.; Moreno-Garcia, J.; Zamfir, C.-I.; Cotea, V.V.; Moreno, J. Selection of aroma compounds for the differentiation of wines obtained by fermenting musts with starter cultures of commercial yeast strains. Food Chem. 2016, 197, 373–381. [Google Scholar] [CrossRef]
- Chen, L.; Renata, R.; Robyn, S.; Vladimir, J.; David, J. Green characters: Using magnetic polymers to remove overpowering green capsicum flavour from Cabernet Sauvignon wine. Wine Vitic. J. 2019, 34, 24–26. [Google Scholar]
- Englezos, V.; Torchio, F.; Cravero, F.; Marengo, F.; Giacosa, S.; Gerbi, V.; Rantsiou, K.; Rolle, L.; Cocolin, L. Aroma profile and composition of Barbera wines obtained by mixed fermentations of Starmerella bacillaris (synonym Candida zemplinina) and Saccharomyces cerevisiae(Article). LWT-Food Sci. Technol. 2016, 73, 567–575. [Google Scholar] [CrossRef]
- Morales, M.L.; Ochoa, M.; Valdivia, M.; Ubeda, C.; Romero-Sanchez, S.; Ibeas, J.I.; Valero, E. Volatile metabolites produced by different flor yeast strains during wine biological ageing. Food Res. Int. 2020, 128, 108771. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.R.; Nie, C.; Xu, X.L. Study on fermentation performance of Chardonnay dry White Wine produced by different yeasts. Brew. China 2016, 35, 98–102. [Google Scholar]
- Xie, S.Y.; Shu, N.; Jin, Y.N.; Xiao, J.M.; Lu, W.P. Effects of different Saccharomyces cerevisiae on volatile compounds of Vitis amurensis Beiguo Distilled Liquor. Brew. China 2020, 39, 150–154. [Google Scholar]
- Zhang, M.Z.; Nan, J. Comparative analysis of different yeasts to improve the quality of Chardonnay wine. Mod. Food Sci. Technol. 2020, 36, 234–240, 159. [Google Scholar] [CrossRef]
- Fan, S.X.; Zhong, Q.D.; Li, G.H.; Chang, D.; Zhang, S.W.; Huang, Z.B. A new method for rapid determination of volatile acid in wine. Food Ferment. Ind. 2018, 44, 216–219. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, W.; Yi, H.; Wang, B.; Xiao, J.; Zhou, X.; Xu, J.; Jiang, L.; Shi, X. Microbial community composition and its role in volatile compound formation during the spontaneous fermentation of ice wine made from Vidal grapes. Process Biochem. 2020, 92, 365–377. [Google Scholar] [CrossRef]
- Wang, X.; Schlatter, D.C.; Glawe, D.A.; Edwards, C.G.; Weller, D.M.; Paulitz, T.C.; Abatzoglou, J.T.; Okubara, P.A. Native yeast and non-yeast fungal communities of Cabernet Sauvignon berries from two Washington State vineyards, and persistence in spontaneous fermentation. Int. J. Food Microbiol. 2021, 350, 109225. [Google Scholar] [CrossRef]
- Sun, D.; Qu, J.; Huang, Y.; Lu, J.; Yin, L. Analysis of microbial community diversity of muscadine grape skins. Food Res. Int. 2021, 145, 110417. [Google Scholar] [CrossRef]
- Martins, G.; Miot-Sertier, C.; Lauga, B.; Claisse, O.; Lonvaud-Funel, A.; Soulas, G.; Masneuf-Pomarede, I. Grape berry bacterial microbiota: Impact of the ripening process and the farming system. Int. J. Food Microbiol. 2012, 158, 93–100. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, E.T.; Singh, R.P.; Guo, C.; Shang, Y.; Chen, J.; Liu, C. Grape berry surface bacterial microbiome: Impact from the varieties and clones in the same vineyard from central China. J. Appl. Microbiol. 2019, 126, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, X.; Zhong, Q.; Zhuang, X.; Bai, Z. Microbial Community Analyses Associated with Nine Varieties of Wine Grape Carposphere Based on High-Throughput Sequencing. Microorganisms 2019, 7, 668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Joseph, C.M.L.; Allen, G.; Benson, A.K.; Mills, D.A. Next-Generation Sequencing Reveals Significant Bacterial Diversity of Botrytized Wine. PLoS ONE 2012, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Godalova, Z.; Krakova, L.; Puskarova, A.; Buckova, M.; Kuchta, T.; Piknova, L.; Pangallo, D. Bacterial consortia at different wine fermentation phases of two typical Central European grape varieties: Blaufrankisch (Frankovka modra) and Gruner Veltliner (Veltlinske zelene). Int. J. Food Microbiol. 2016, 217, 110–116. [Google Scholar] [CrossRef]
- Rivas, G.A., II; Semorile, L.; Delfederico, L. Microbial diversity of the soil, rhizosphere and wine from an emerging wine-producing region of Argentina. LWT-Food Sci. Technol. 2022, 153, 112429. [Google Scholar] [CrossRef]
- Mercado, L.; Masuelli, R.; Dalcero, A.; Combina, M. Diversity of Saccharomyces strains on grapes and winery surfaces: Analysis of their contribution to fermentative flora of Malbec wine from Mendoza (Argentina) during two consecutive years. Food Microbiol. 2007, 24, 403–412. [Google Scholar] [CrossRef]
- Morgan, S.C.; McCarthy, G.C.; Watters, B.S.; Tantikachornkiat, M.; Zigg, I.; Cliff, M.A.; Durall, D.M. Effect of sulfite addition and pied de cuve inoculation on the microbial communities and sensory profiles of Chardonnay wines: Dominance of indigenous Saccharomyces uvarum at a commercial winery. FEMS Yeast Res. 2019, 19, foz049. [Google Scholar] [CrossRef]
- Mendes Ferreira, A.; Mendes-Faia, A. The Role of Yeasts and Lactic Acid Bacteria on the Metabolism of Organic Acids during Winemaking. Foods 2020, 9, 1231. [Google Scholar] [CrossRef]
- Lin, X.; Wang, Q.; Hu, X.; Wu, W.; Zhang, Y.; Liu, S.; Li, C. Evaluation of different Saccharomyces cerevisiae strains on the profile of volatile compounds in pineapple wine. J. Food Sci. Technol.-Mysore 2018, 55, 4119–4130. [Google Scholar] [CrossRef]
- Welke, J.E.; Zanus, M.; Lazzarotto, M.; Zini, C.A. Quantitative analysis of headspace volatile compounds using comprehensive two-dimensional gas chromatography and their contribution to the aroma of Chardonnay wine. Food Res. Int. 2014, 59, 85–99. [Google Scholar] [CrossRef] [Green Version]
- de-la-Fuente-Blanco, A.; Saenz-Navajas, M.-P.; Valentin, D.; Ferreira, V. Fourteen ethyl esters of wine can be replaced by simpler ester vectors without compromising quality but at the expense of increasing aroma concentration. Food Chem. 2020, 307, 125553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado de Castilhos, M.B.; Del Bianchi, V.L.; Gomez-Alonso, S.; Garcia-Romero, E.; Hermosin-Gutierrez, I. Sensory descriptive and comprehensive GC-MS as suitable tools to characterize the effects of alternative winemaking procedures on wine aroma. Part II: BRS Rubea and BRS Cora. Food Chem. 2020, 311, 126025. [Google Scholar] [CrossRef]
- Styger, G.; Prior, B.; Bauer, F.F. Wine flavor and aroma. J. Ind. Microbiol. Biotechnol. 2011, 38, 1145–1159. [Google Scholar] [CrossRef] [PubMed]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Lan, Y.; Liu, M.; Zhang, X.; Li, S.; Shi, Y.; Duan, C. Regional Variation of Chemical Characteristics in Young Marselan (Vitis vinifera L.) Red Wines from Five Regions of China. Foods 2022, 11, 787. [Google Scholar] [CrossRef]
- de-la-Fuente-Blanco, A.; Sáenz-Navajas, M.-P.; Ferreira, V. On the effects of higher alcohols on red wine aroma. Food Chem. 2016, 210, 107–114. [Google Scholar] [CrossRef]
- Sadoudi, M.; Tourdot-Maréchal, R.; Rousseaux, S. Yeast–yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef]
- Rebecca, C.; Deed, C.; Fedrizzi, B.; Gardner, R.C. Influence of Fermentation Temperature, Yeast Strain, and Grape Juice on the Aroma Chemistry and Sensory Profile of Sauvignon Blanc Wines(Article). J. Agric. Food Chem. 2017, 65, 8902–8912. [Google Scholar]
- Schreier, P. Flavor composition of wines: A review. CRC Crit. Rev. Food Sci. Nutr. 1979, 12, 59–111. [Google Scholar] [CrossRef]
- Lonvaud-Funel, A. Lactic acid bacteria in the quality improvement and depreciation of wine. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 1999, 76, 317–331. [Google Scholar] [CrossRef]
- Guo, L.; Luo, Y.; Zhou, Y.; Bianba, C.; Guo, H.; Zhao, Y.; Fu, H. Exploring microbial dynamics associated with flavours production during highland barley wine fermentation. Food Res. Int. 2020, 130, 108971. [Google Scholar] [CrossRef]
- Mota, R.V.D.; Peregrino, I.; Rivera, S.P.T.; Hassimotto, N.M.A.; Souza, A.L.D.; Souza, C.R.D. Characterization of Brazilian Syrah winter wines at bottling and after ageing. Sci. Agric. 2021, 78, e20190233. [Google Scholar] [CrossRef]
- Baneras, L.; Trias, R.; Godayol, A.; Cerdan, L.; Nawrath, T.; Schulz, S.; Antico, E. Mass spectrometry identification of alkyl-substituted pyrazines produced by Pseudomonas spp. isolates obtained from wine corks. Food Chem. 2013, 138, 2382–2389. [Google Scholar] [CrossRef] [PubMed]
- West, T.P. Effect of temperature on bacterial gellan production. World J. Microbiol. Biotechnol. 2003, 19, 649–652. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among Wine Grape Microbiome, Metabolome, and Fermentation Behavior Suggest Microbial Contribution to Regional Wine Characteristics. Mbio 2016, 7, e00631-16. [Google Scholar] [CrossRef] [Green Version]
- Andorra, I.; Berradre, M.; Rozes, N.; Mas, A.; Guillamon, J.M.; Esteve-Zarzoso, B. Effect of pure and mixed cultures of the main wine yeast species on grape must fermentations. Eur. Food Res. Technol. 2010, 231, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Ciani, M.; Comitini, F. Yeast interactions in multi-starter wine fermentation. Curr. Opin. Food Sci. 2015, 1, 1–6. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Time | CECA | CEC01 | F15 | 796 |
|---|---|---|---|---|
| Day 0 | C0d | E0d | F0d | S0d |
| Day 2 | C2d | E2d | F2d | S2d |
| Day 4 | C4d | E4d | F4d | S4d |
| Day 6 | C6d | E6d | F6d | S6d |
| Starters | Fermentation Days | Reducing Sugar (g/L) | Ethanol (% vol) | pH | Total Acid (g/L) |
|---|---|---|---|---|---|
| CECA | Day 0 | 221.33 ± 0.93 a | 0 | 3.71 ± 0.01 a | 6.23 ± 0.06 a |
| Day 2 | 177.23 ± 2.24 b | 2.01 ± 0.04 b | 3.80 ± 0.01 a | 4.00 ± 0.01 b | |
| Day 4 | 44.00 ± 0.56 c | 10.47 ± 0.02 a | 3.89 ± 0.006 a | 4.13 ± 0.06 b | |
| Day 6 | 0.70 ± 0.20 d | 12.37 ± 0.04 a | 3.82 ± 0.006 a | 4.20 ± 0.02 b | |
| CEC01 | Day 0 | 224.77 ± 0.06 a | 0 | 3.67 ± 0.006 a | 6.57 ± 0.06 a |
| Day 2 | 132.73 ± 1.16 b | 5.21 ± 0.03 b | 3.72 ± 0.006 a | 4.63 ± 0.06 b | |
| Day 4 | 27.50 ± 0.44 c | 11.74 ± 0.04 a | 3.79 ± 0.006 a | 4.50 ± 0.01 b | |
| Day 6 | 1.00 ± 0.26 d | 12.67 ± 0.01 a | 3.75 ± 0.006 a | 4.30 ± 0.01 b | |
| F15 | Day 0 | 225.40 ± 0.20 a | 0 | 3.66 ± 0.01 c | 6.56 ± 0.06 a |
| Day 2 | 212.50 ± 1.36 b | 0.036 ± 0.02 d | 3.68 ± 0.1 b | 3.60 ± 0.02 d | |
| Day 4 | 64.63 ± 1.36 c | 9.17 ± 0.01 c | 3.68 ± 0.006 b | 4.56 ± 0.06 b | |
| Day 6 | 0.63 ± 0.12 d | 12.66 ± 0.02 b | 3.72 ± 0.002 a | 4.53 ± 0.06 b | |
| 796 | Day 0 | 222.00 ± 0.89 a | 0 | 3.69 ± 0.06 b | 6.93 ± 0.06 a |
| Day 2 | 170.43 ± 2.00 b | 2.19 ± 0.09 d | 3.72 ± 0.01 a | 4.53 ± 0.06 d | |
| Day 4 | 51.03 ± 0.93 c | 9.64 ± 0.01 c | 3.73 ± 0.06 a | 5.57 ± 0.06 c | |
| Day 6 | 1.17 ± 0.06 d | 12.41 ± 0.02 b | 3.74 ± 0.06 a | 5.73 ± 0.06 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Wang, J.; Chen, J.; Philipp, C.; Zhao, X.; Wang, J.; Liu, Y.; Suo, R. Effect of Commercial Yeast Starter Cultures on Cabernet Sauvignon Wine Aroma Compounds and Microbiota. Foods 2022, 11, 1725. https://doi.org/10.3390/foods11121725
Wang M, Wang J, Chen J, Philipp C, Zhao X, Wang J, Liu Y, Suo R. Effect of Commercial Yeast Starter Cultures on Cabernet Sauvignon Wine Aroma Compounds and Microbiota. Foods. 2022; 11(12):1725. https://doi.org/10.3390/foods11121725
Chicago/Turabian StyleWang, Meiqi, Jiarong Wang, Jiawei Chen, Christian Philipp, Xiaoning Zhao, Jie Wang, Yaqiong Liu, and Ran Suo. 2022. "Effect of Commercial Yeast Starter Cultures on Cabernet Sauvignon Wine Aroma Compounds and Microbiota" Foods 11, no. 12: 1725. https://doi.org/10.3390/foods11121725