
Preventive Effects of Thinned Apple Extracts on TNF-α-Induced Intestinal Tight Junction Dysfunction in Caco-2 Cells through Myosin Light Chain Kinase Suppression


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Sample Preparation
2.3. Cell Culture and Treatment
2.4. Determination of Transepithelial Electrical Resistance and Paracellular Permeability
2.5. RNA Isolation and Reverse Transcription
2.6. Gene Expression by Quantitative RT-PCR Analysis
2.7. Preparation of Cell Lysate and Immunoblot Analysis
2.8. Immunoprecipitation
2.9. Polyphenol Contents by High-Performance Liquid Chromatography
2.10. Statistics
3. Results
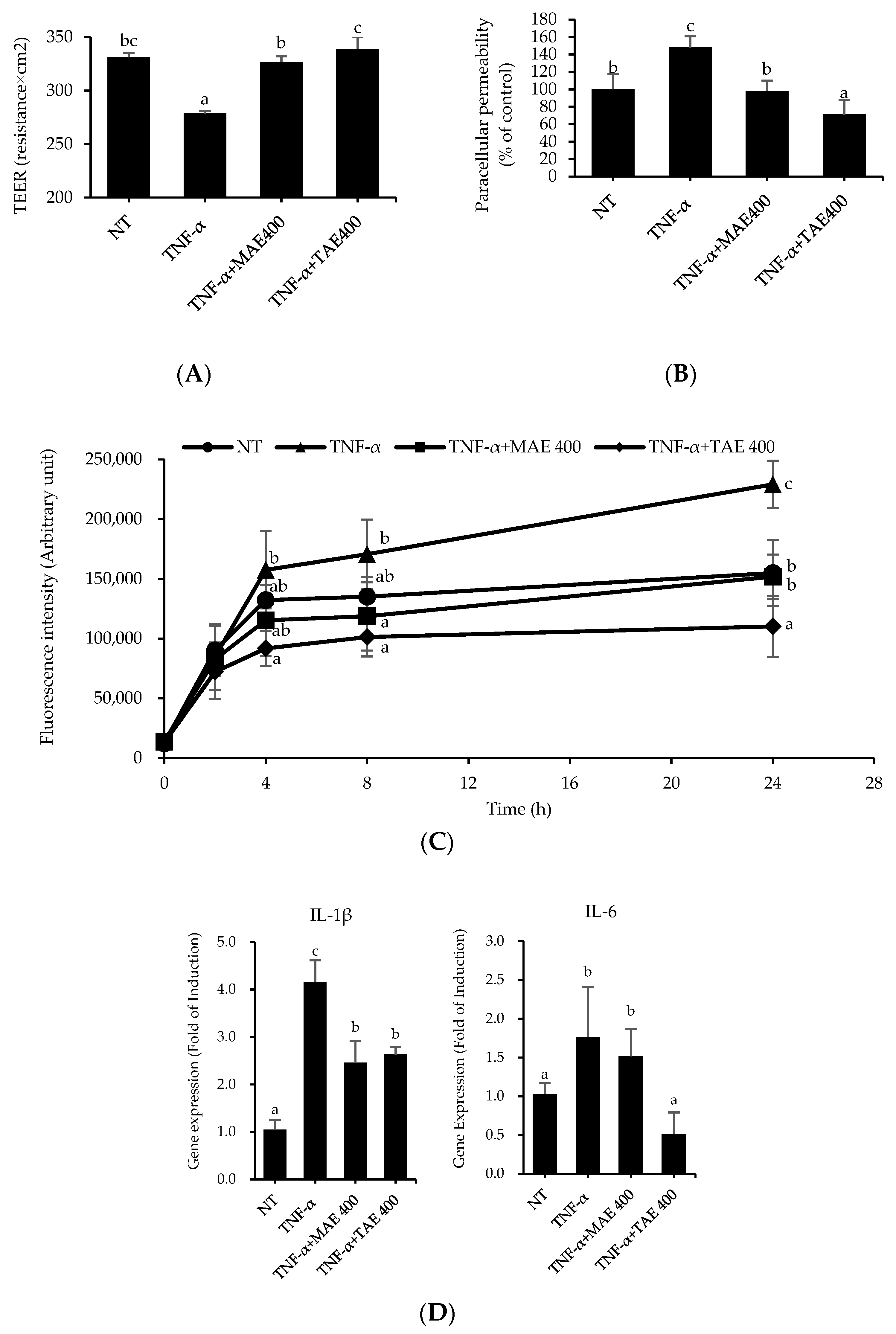
3.1. The Thinned Apple Extract Is Superior to Mature Apple Extract in the Prevention of TNF-α-Mediated Deterioration of TJ Integrity
3.2. Quantification of Polyphenolic Compounds Present in the Mature and Thinned Apple Extracts by HPLC Analysis
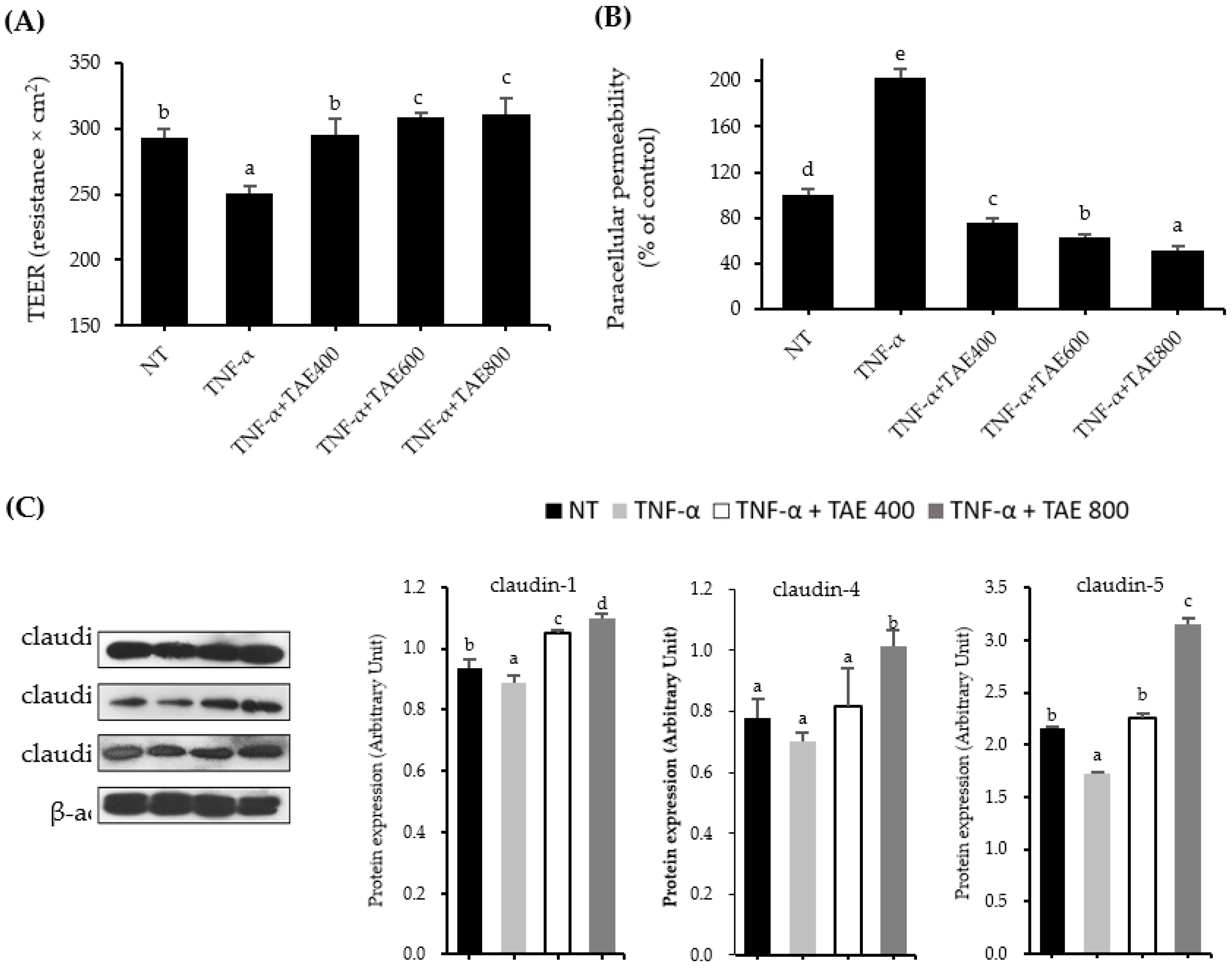
3.3. Thinned Apple Extracts Prevent TNF-α-Mediated TJ Alteration by Regulating TJ Protein Expression
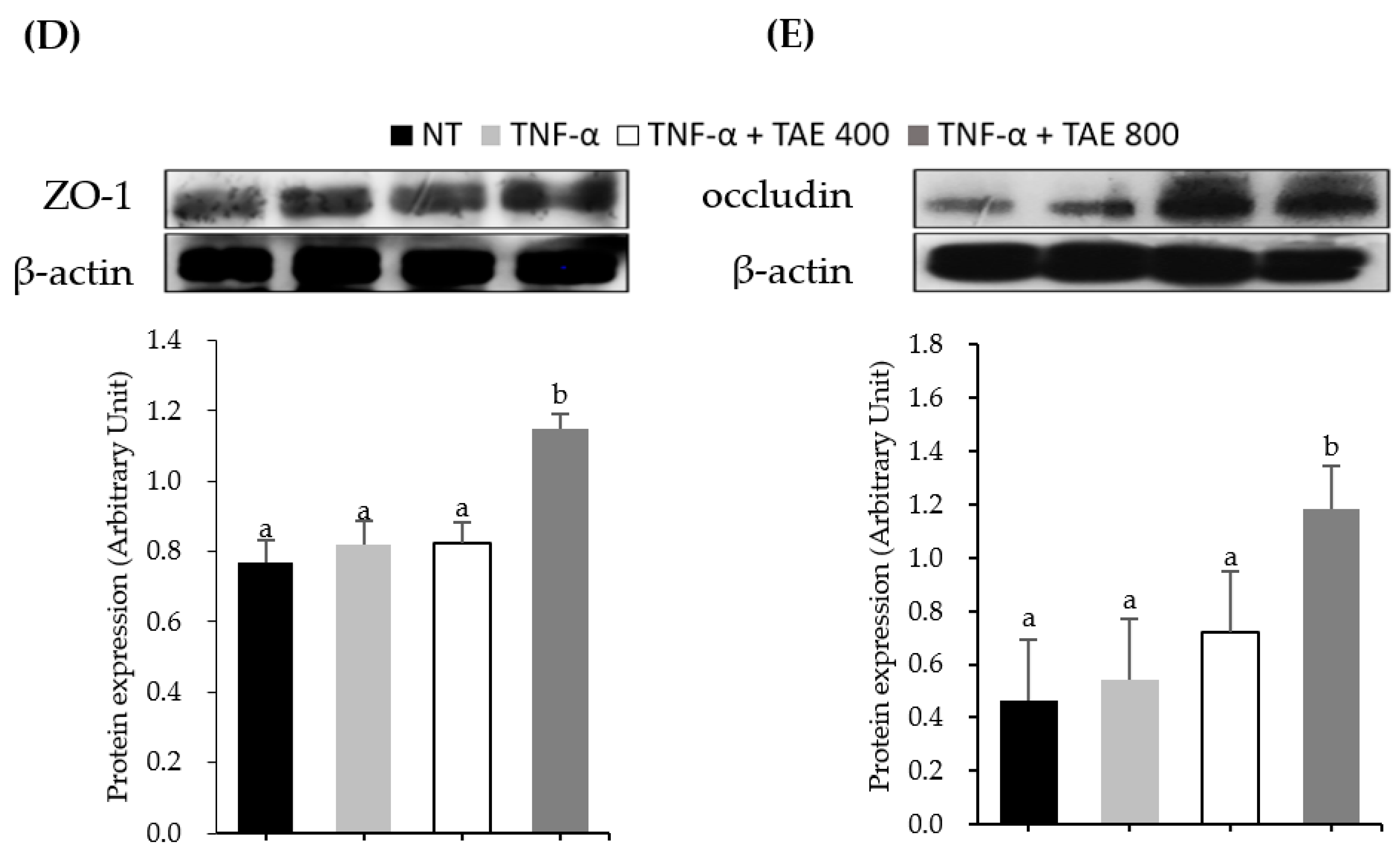
3.4. Thinned Apple Extract Prevents TJ Disruption by Modulating the Interaction between ZO-1-Occludin and Myosin Light Chain Kinase Protein
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, C.G.; Lee, S.Y.; Joo, S.Y.; Cho, L.H.; Park, S.Y.; Lee, S.H.; Oh, K.C.; Kim, D.H. A Study on Agricultural by-products for Biomass-to-energy Conversion and Korean Collecting Model. New Renew. Energy 2017, 13, 27–35. [Google Scholar] [CrossRef]
- Bhatia, L.; Johri, S.; Ahmad, R. An economic and ecological perspective of ethanol production from renewable agro waste: A review. AMB Express 2012, 2, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, S.; Bhushan, S. Apple phenolics as nutraceuticals: Assessment, analysis and application. J. Food Sci. Technol. 2016, 53, 1727–1738. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Akazome, Y.; Kanda, T.; Ikeda, M. The toxicology and safety of apple polyphenol extract. Food Chem. Toxicol. 2004, 42, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerhauser, C. Cancer Chemopreventive Potential of Apples, Apple Juice, and Apple Components. Planta Med. 2008, 74, 1608–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef]
- Schneeberger, E.E.; Lynch, R.D. The tight junction: A multifunctional complex. Am. J. Physiol. Physiol. 2004, 286, C1213–C1228. [Google Scholar] [CrossRef]
- Lee, B.; Moon, K.M.; Kim, C.Y. Tight Junction in the Intestinal Epithelium: Its Association with Diseases and Regulation by Phytochemicals. J. Immunol. Res. 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Baumgart, D.C.; Carding, S. Inflammatory bowel disease: Cause and immunobiology. Lancet 2007, 369, 1627–1640. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Boivin, M.; Ma, T. Mechanism of cytokine modulation of epithelial tight junction barrier. Front. Biosci. 2009, 14, 2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.Y. Inhibition of interleukin-1α-induced intestinal epithelial tight junction permeability by curcumin treatment in Caco-2 Cells in Caco-2 cells. J. Life Sci 2016, 26, 1082–1087. [Google Scholar] [CrossRef]
- Cao, M.; Wang, P.; Sun, C.; He, W.; Wang, F. Amelioration of IFN-γ and TNF-α-induced intestinal epithelial barrier dysfunction by berberine via suppression of MLCK-MLC phosphorylation signaling pathway. PLoS ONE 2013, 8, e61944. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Gu, L.; Qu, L.; Gong, J.; Li, Q.; Zhu, W.; Li, J. Berberine attenuates pro-inflammatory cytokine-induced tight junction disruption in an in vitro model of intestinal epithelial cells. Eur. J. Pharm. Sci. 2010, 40, 1–8. [Google Scholar] [CrossRef]
- Suzuki, T.; Tanabe, S.; Hara, H. Kaempferol Enhances Intestinal Barrier Function through the Cytoskeletal Association and Expression of Tight Junction Proteins in Caco-2 Cells. J. Nutr. 2011, 141, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Hara, H. Quercetin Enhances Intestinal Barrier Function through the Assembly of Zonnula Occludens-2, Occludin, and Claudin-1 and the Expression of Claudin-4 in Caco-2 Cells. J. Nutr. 2009, 139, 965–974. [Google Scholar] [CrossRef]
- Günzel, D.; Yu, A.S.L. Claudins and the Modulation of Tight Junction Permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, B.T.; Wang, F.; Shen, L.; Clayburgh, D.; Su, L.; Wang, Y.; Fu, Y.-X.; Turner, J.R. LIGHT Signals Directly to Intestinal Epithelia to Cause Barrier Dysfunction via Cytoskeletal and Endocytic Mechanisms. Gastroenterology 2007, 132. [Google Scholar] [CrossRef] [Green Version]
- Amasheh, M.; Fromm, A.; Krug, S.M.; Amasheh, S.; Andres, S.; Zeitz, M.; Fromm, M.; Schulzke, J.D. TNFα-induced and berberine-antagonized tight junction barrier impairment via tyrosine kinase, Akt and NFκB signaling. J. Cell Sci. 2010, 123, 4145–4155. [Google Scholar] [CrossRef] [Green Version]
- Vreeburg, R.A.; E van Wezel, E.; Ocaña-Calahorro, F.; Mes, J.J. Apple extract induces increased epithelial resistance and claudin 4 expression in Caco-2 cells. J. Sci. Food Agric. 2012, 92, 439–444. [Google Scholar] [CrossRef]
- Finotti, E.; Gezzi, R.; Nobili, F.; Garaguso, I.; Friedman, M. Effect of apple, baobab, red-chicory, and pear extracts on cellular energy expenditure and morphology of a Caco-2 cells using transepithelial electrical resistance (TEER) and scanning electron microscopy (SEM). RSC Adv. 2015, 5, 22490–22498. [Google Scholar] [CrossRef]
- Wong, X.; Carrasco-Pozo, C.; Escobar, E.; Navarrete, P.; Blachier, F.; Andriamihaja, M.; Lan, A.; Tome, A.; Cires, M.J.; Pastene, E.; et al. Deleterious effect of p-cresol on human colonic epithelial cells prevented by proanthocyanidin-containing polyphenol extracts from fruits and proanthocyanidin bacterial metabolites. J. Agric. Food Chem. 2016, 64, 3574–3583. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, Q.; Zang, Y.; Zhao, Y.; Liu, N.; Wang, Y.; Xu, X.; Liu, L.; Mei, Q. Apple Polysaccharide inhibits microbial dysbiosis and chronic inflammation and modulates gut permeability in HFD-fed rats. Int. J. Biol. Macromol. 2017, 99, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Lauren, D.R.; Smith, W.A.; Adaim, A.; Cooney, J.M.; Wibisono, R.; Jensen, D.J.; Zhang, J.; Skinner, M.A. Chemical composition and in vitro anti-inflammatory activity of apple phenolic extracts and of their sub-fractions. Int J Food Sci Nutr. 2009, 60, 188–205. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Akiyama, H.; Nakano, M.; Shoji, T.; Kanda, T.; Ohtake, Y.; Takita, T.; Matsuda, R.; Maitani, T. Orally administered apple procyanidins protect against experimental inflammatory bowel disease in mice. Int. Immunopharmacol. 2008, 8, 1802–1807. [Google Scholar] [CrossRef]
- Skyberg, J.A.; Robison, A.; Golden, S.; Rollins, M.F.; Callis, G.; Huarte, E.; Kochetkova, I.; Jutila, M.A.; Pascual, D.W. Apple polyphenols require T cells to ameliorate dextran sulfate sodium-induced colitis and dampen proinflammatory cytokine expression. J. Leukoc. Biol. 2011, 90, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.-Z.; Kim, Y.-I.; Chung, S.-K. A profile of physicochemical and antioxidant changes during fruit growth for the utilisation of unripe apples. Food Chem. 2012, 131, 106–110. [Google Scholar] [CrossRef]
- Zhang, P.; Jiao, H.; Wang, C.; Lin, Y.; You, S. Chlorogenic Acid Ameliorates Colitis and Alters Colonic Microbiota in a Mouse Model of Dextran Sulfate Sodium-Induced Colitis. Front. Physiol. 2019, 17, 107–114. [Google Scholar] [CrossRef]
- Akazome, Y.; Kametani, N.; Kanda, T.; Shimasaki, H.; Kobayashi, S. Evaluation of Safety of Excessive Intake and Efficacy of Long-term Intake of Beverages Containing Apple Polyphenols. J. Oleo Sci. 2010, 59, 321–338. [Google Scholar] [CrossRef] [Green Version]
- Miura, D.; Miura, Y.; Yagasaki, K. Effect of Apple Polyphenol Extract on Hepatoma Proliferation and Invasion in Culture and on Tumor Growth, Metastasis, and Abnormal Lipoprotein Profiles in Hepatoma-Bearing Rats. Biosci. Biotechnol. Biochem. 2007, 71, 2743–2750. [Google Scholar] [CrossRef] [Green Version]
- Barmeyer, C.; Erko, I.; Awad, K.; Fromm, A.; Bojarski, C.; Meissner, S.; Loddenkemper, C.; Kerick, M.; Siegmund, B.; Fromm, M.; et al. Epithelial barrier dysfunction in lymphocytic colitis through cytokine-dependent internalization of claudin-5 and -8. J. Gastroenterol. 2017, 52, 1090–1100. [Google Scholar] [CrossRef] [PubMed]
- Watari, A.; Sakamoto, Y.; Hisaie, K.; Iwamoto, K.; Fueta, M.; Yagi, K.; Kondoh, M. Rebeccamycin Attenuates TNF-α-Induced Intestinal Epithelial Barrier Dysfunction by Inhibiting Myosin Light Chain Kinase Production. Cell. Physiol. Biochem. 2017, 41, 1924–1934. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ding, X.; Miao, C.; Chen, J. Propofol attenuated TNF-α-modulated occludin expression by inhibiting Hif-1α/ VEGF/ VEGFR-2/ ERK signaling pathway in hCMEC/D3 cells. BMC Anesthesiol. 2019, 19, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Mariscal, L.; Tapia, R.; Chamorro, D. Crosstalk of tight junction components with signaling pathways. Biochim. Biophys. Acta (BBA)-Biomembr. 2008, 1778, 729–756. [Google Scholar] [CrossRef] [Green Version]
- Kawauchiya, T.; Takumi, T.; Kudo, Y.; Takamori, A.; Sasagawa, T.; Takahashi, K.; Kikuchi, H. Correlation between the destruction of tight junction by patulin treatment and increase of phosphorylation of ZO-1 in Caco-2 human colon cancer cells. Toxicol. Lett. 2011, 205, 196–202. [Google Scholar] [CrossRef]
- Piegholdt, S.; Pallauf, K.; Esatbeyoglu, T.; Speck, N.; Reiss, K.; Ruddigkeit, L.; Stocker, A.; Huebbe, P.; Rimbach, G. Biochanin A and prunetin improve epithelial barrier function in intestinal CaCo-2 cells via downregulation of ERK, NF-κB, and tyrosine phosphorylation. Free Radic. Biol. Med. 2014, 70, 255–264. [Google Scholar] [CrossRef]
- Ma, T.Y.; Boivin, M.A.; Ye, D.; Pedram, A.; Said, H.M. Mechanism of TNF-α modulation of Caco-2 intestinal epithelial tight junction barrier: Role of myosin light-chain kinase protein expression. Am. J. Physiol. Liver Physiol. 2005, 288, G422–G430. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Sun, G.; Wang, Y.; Yu, P.; Wang, X.; Zhou, B.; Zhu, H. Glabridin attenuates endothelial dysfunction and permeability, possibly via the MLCK/p MLC signaling pathway. Exp. Ther. Med. 2019, 17, 107–114. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Orientation | Primers Sequence (5′→3′) | NCBI Accession NO. |
|---|---|---|---|
| COX-2 | Forward | CAAATCCTTGCTGT TCCCACCCAT | NM_000963.4 |
| Reverse | GTGCACTGTGTTTGGAGTGGGTTT | ||
| IL-6 | Forward | GGTACATCCTCGACGGCATCT | NM_001371096.1 |
| Reverse | GTGCCTCTTTGCTGCTTTCAC | ||
| TNF-α | Forward | CCCAGGCAGTCAGATCATCTTC | NM_000594.4 |
| Reverse | AGCTGCCCCTCAGCTTGA | ||
| IL-1β | Forward | TGGCAATGAGGATGACTTGTTC | NM_000576.3 |
| Reverse | CTGTAGTGGTGGTCGGAGATT | ||
| Claudin-3 | Forward | AAGGTGTACGACTCGCTGCT | NM_001306.4 |
| Reverse | AGTCCCGGATAATGGTGTTG | ||
| Claudin-5 | Forward | CTCTGCTGGTTCGCCAACAT | NM_003277.4 |
| Reverse | CACAGACGGGTCGTAAAACTC | ||
| 18S rRNA | Forward | GATGGAAAATACAGCCAGGTCCTA | NM_022551.3 |
| Reverse | TTCTTCAGTCGCTCCAGGTCTT |
| Total Polyphenols | Thinned Apple Extract (µg/g) | Mature Apple Extract (µg/g) |
|---|---|---|
| Chlorogenic acid | 6172.03 | 965.02 *** |
| Epicatechin | 2027.13 | 171.18 *** |
| Catechin | 1646.39 | 380.81 *** |
| Phloridizin | 1081.61 | 260.99 *** |
| Procyanidin B1 | 792.32 | 320.41 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-Y.; Kim, C.Y. Preventive Effects of Thinned Apple Extracts on TNF-α-Induced Intestinal Tight Junction Dysfunction in Caco-2 Cells through Myosin Light Chain Kinase Suppression. Foods 2022, 11, 1714. https://doi.org/10.3390/foods11121714
Lee J-Y, Kim CY. Preventive Effects of Thinned Apple Extracts on TNF-α-Induced Intestinal Tight Junction Dysfunction in Caco-2 Cells through Myosin Light Chain Kinase Suppression. Foods. 2022; 11(12):1714. https://doi.org/10.3390/foods11121714
Chicago/Turabian StyleLee, Joo-Yeon, and Choon Young Kim. 2022. "Preventive Effects of Thinned Apple Extracts on TNF-α-Induced Intestinal Tight Junction Dysfunction in Caco-2 Cells through Myosin Light Chain Kinase Suppression" Foods 11, no. 12: 1714. https://doi.org/10.3390/foods11121714