
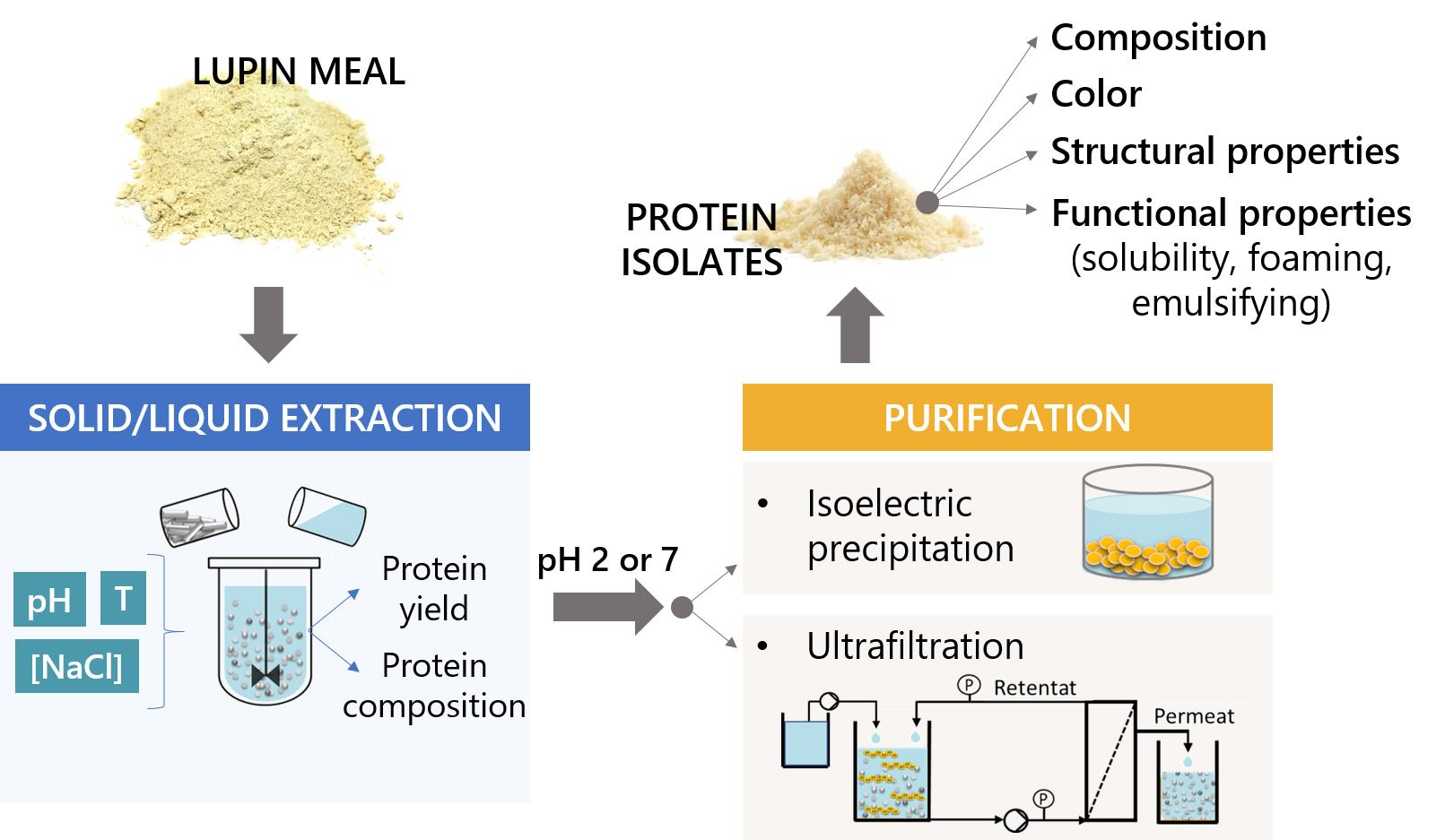
Combined Effect of Extraction and Purification Conditions on Yield, Composition and Functional and Structural Properties of Lupin Proteins


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Effect of Extraction Conditions
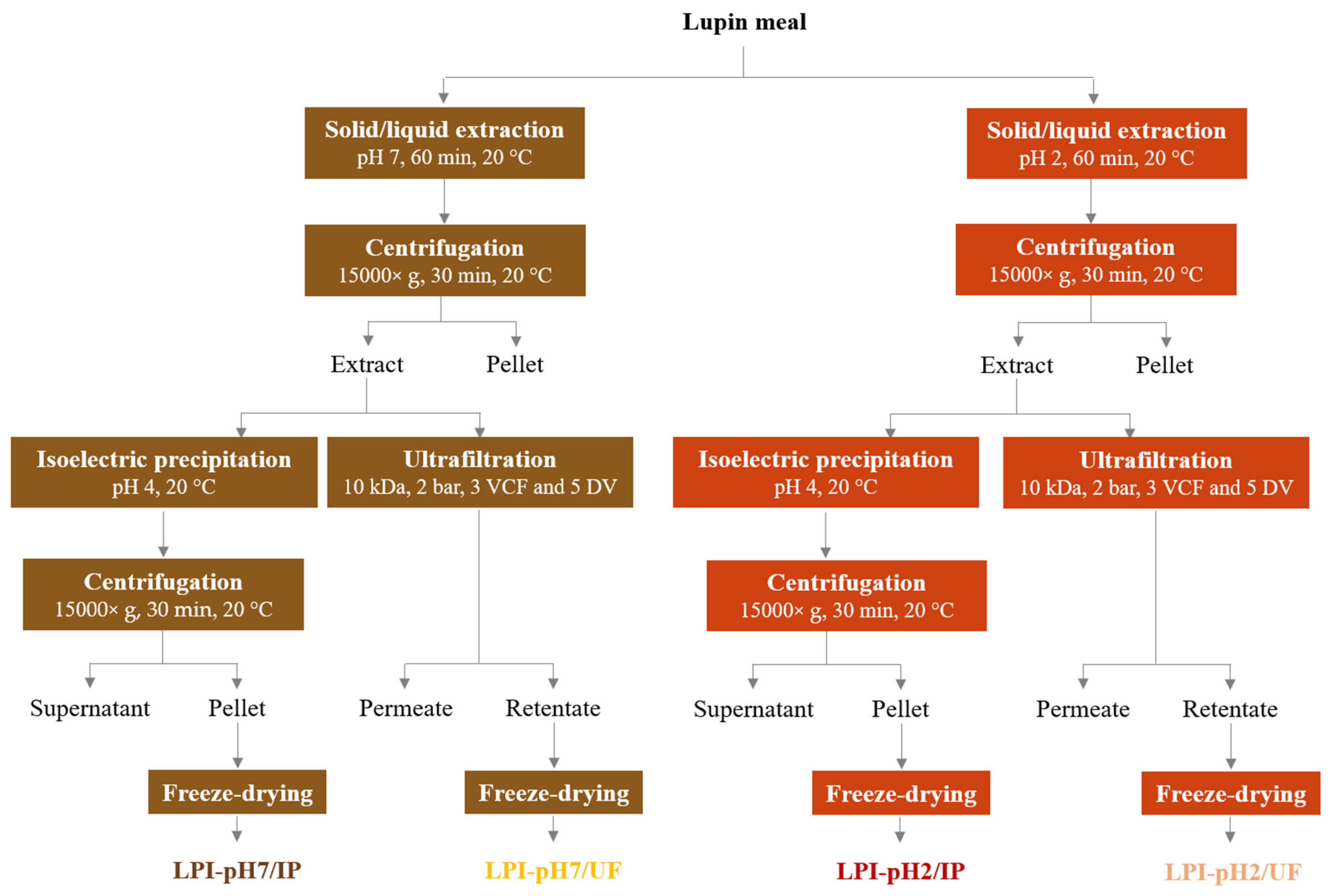
2.3. Preparation of Lupin Protein Isolates
2.3.1. Purification by Isoelectric Precipitation
2.3.2. Purification by Ultrafiltration
2.4. Analytical Methods
2.4.1. Determination of Protein Content and Protein Process Yields
2.4.2. Electrophoresis on Gel
2.5. Characterization of Isolated Proteins
2.5.1. Protein Purity and Solubility
2.5.2. Measurement of Colour
2.5.3. Functional Properties
2.5.4. Structural Properties
- Circular Dichroism
- Differential Scanning Calorimetry
- Dynamic Light Scattering
2.6. Statistical Analysis
3. Results and Discussion
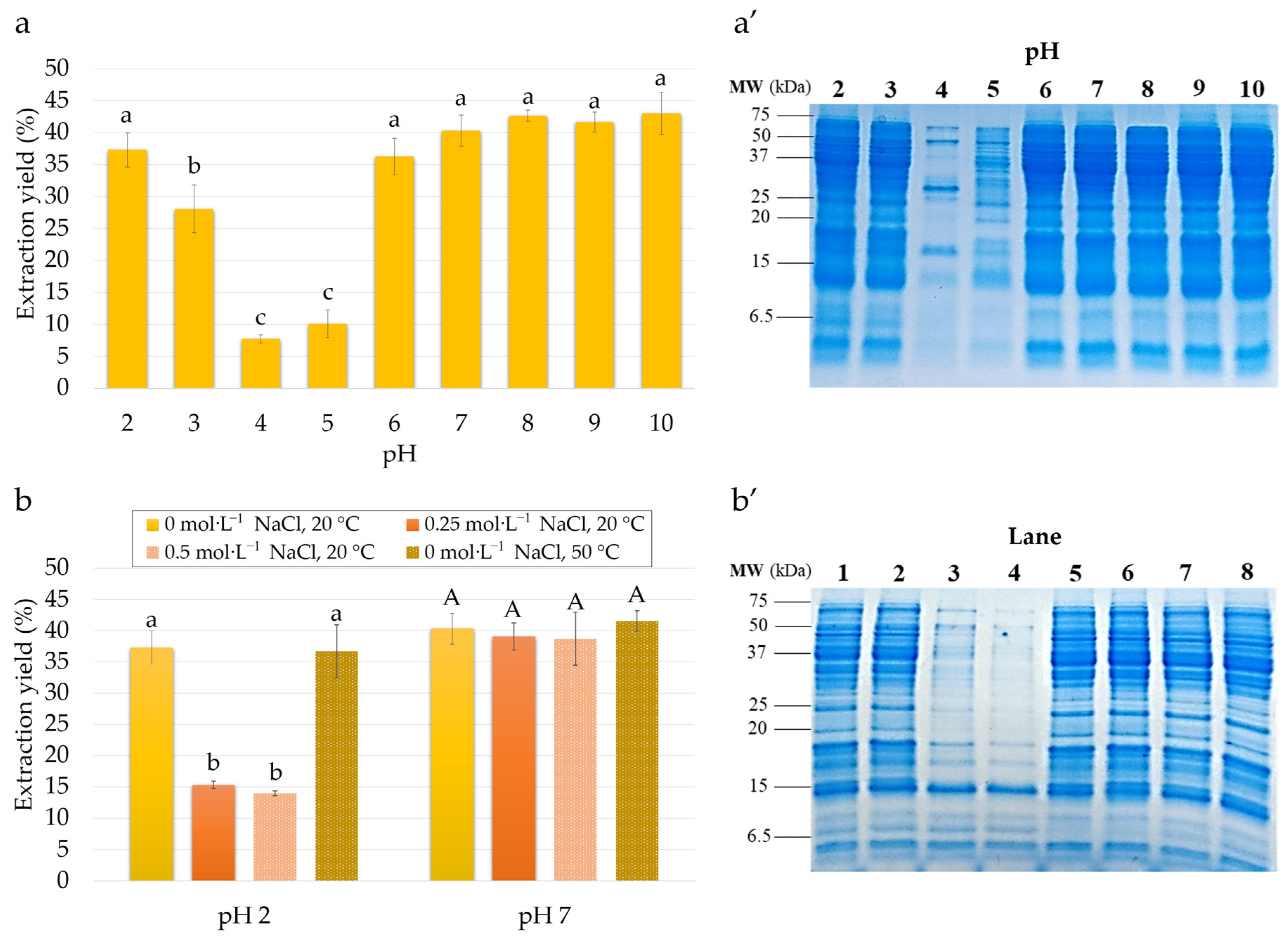
3.1. Effect of Extraction Conditions on Extraction Yield and Protein Composition
3.1.1. Effect of pH
3.1.2. Effect of NaCl Concentration
3.1.3. Effect of Temperature
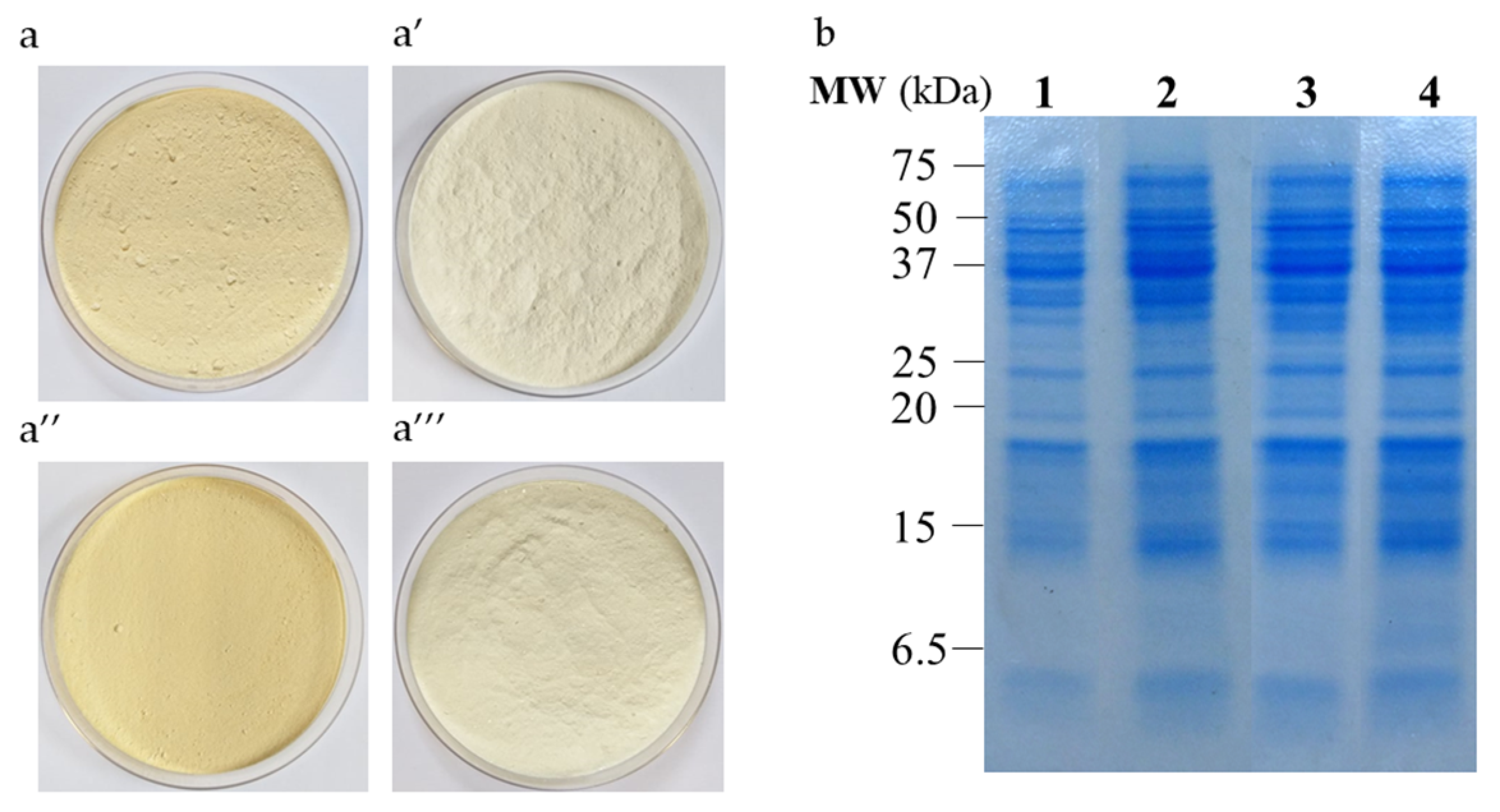
3.2. Combined Effect of Extraction pH and Purification Process on Protein Recovery, Composition and Color of Lupin Protein Isolates
3.3. Effect of Extraction Conditions on Yield and Protein Properties
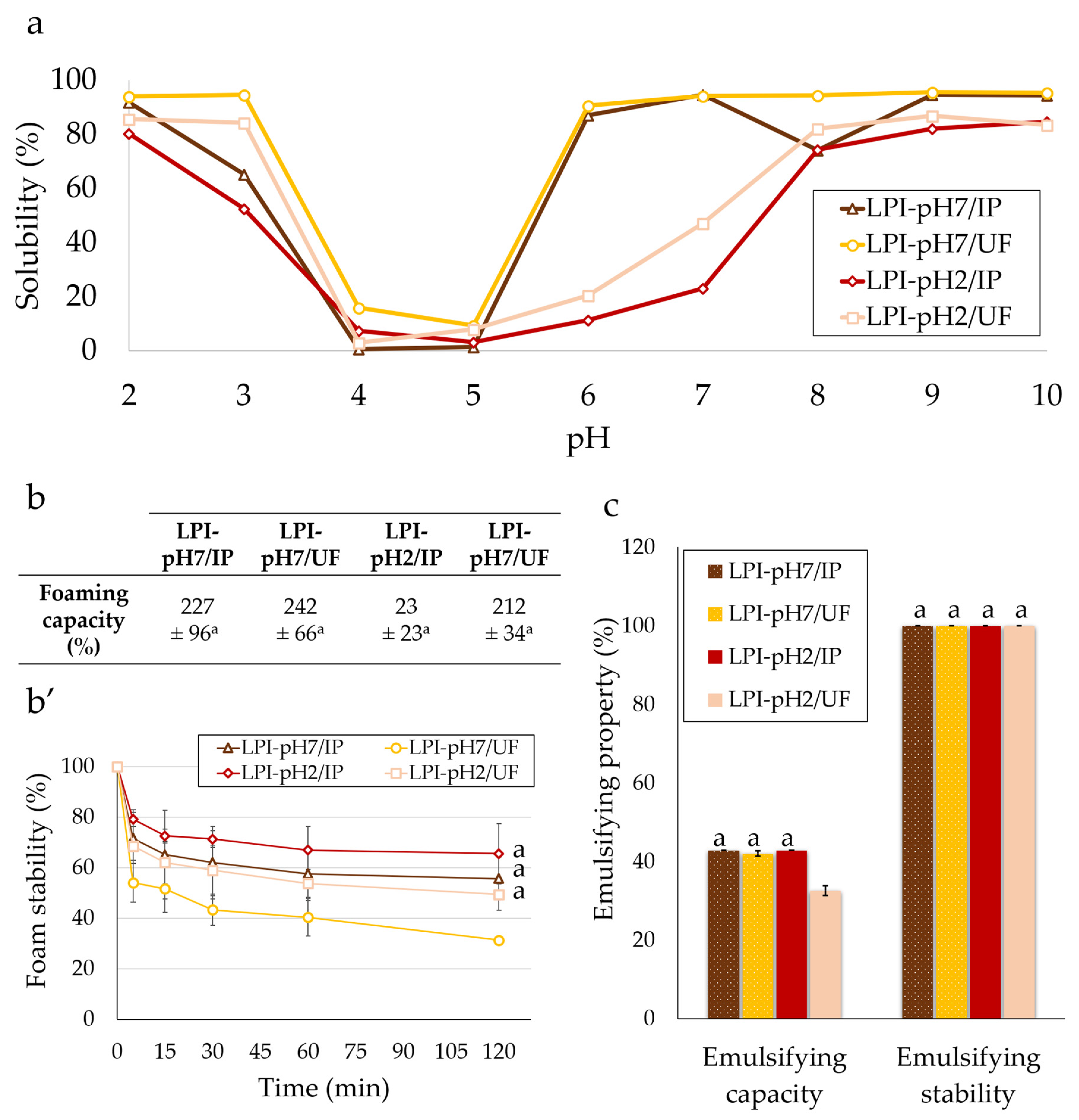
3.3.1. Solubility
3.3.2. Foaming
3.3.3. Emulsifying
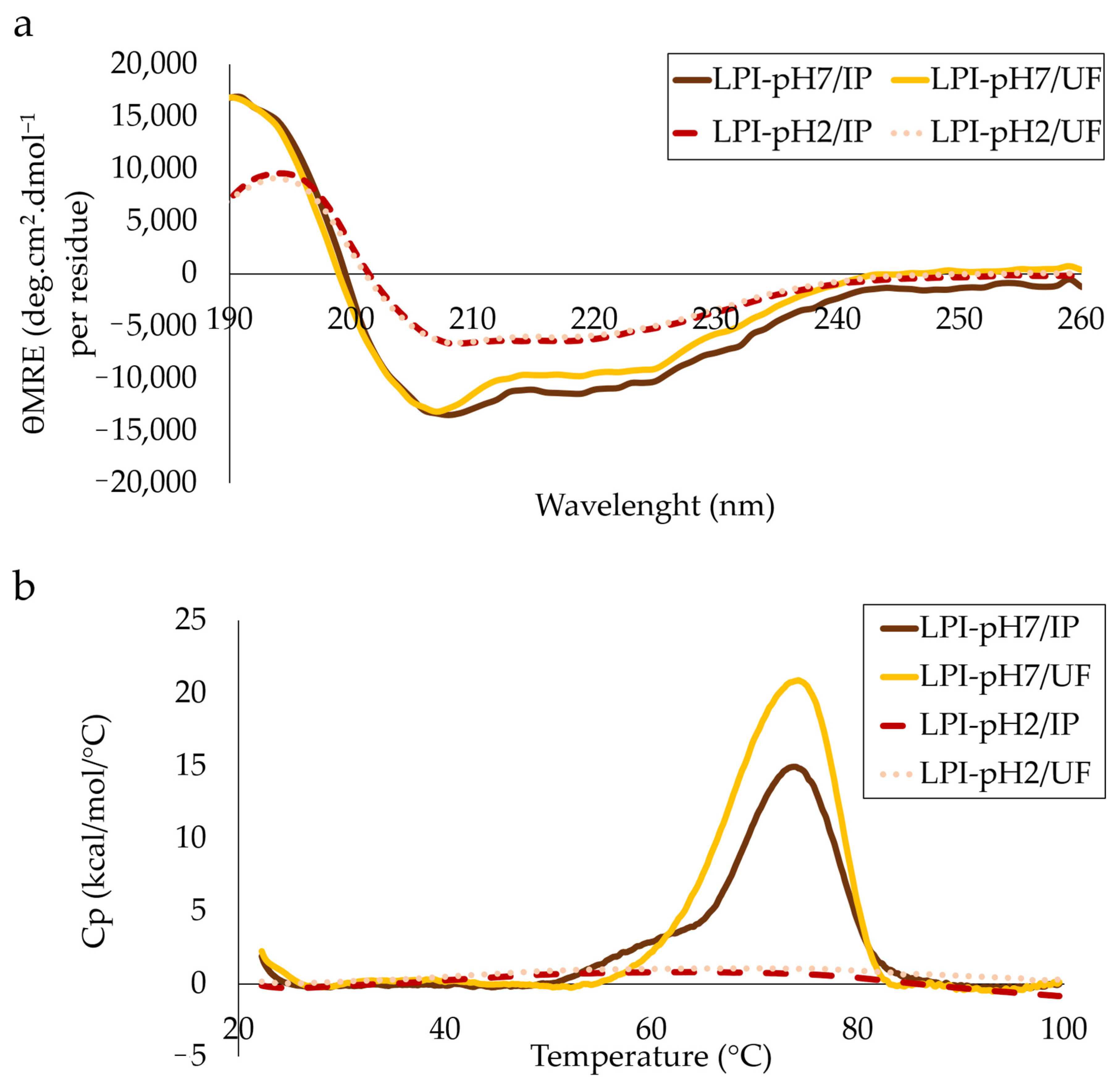
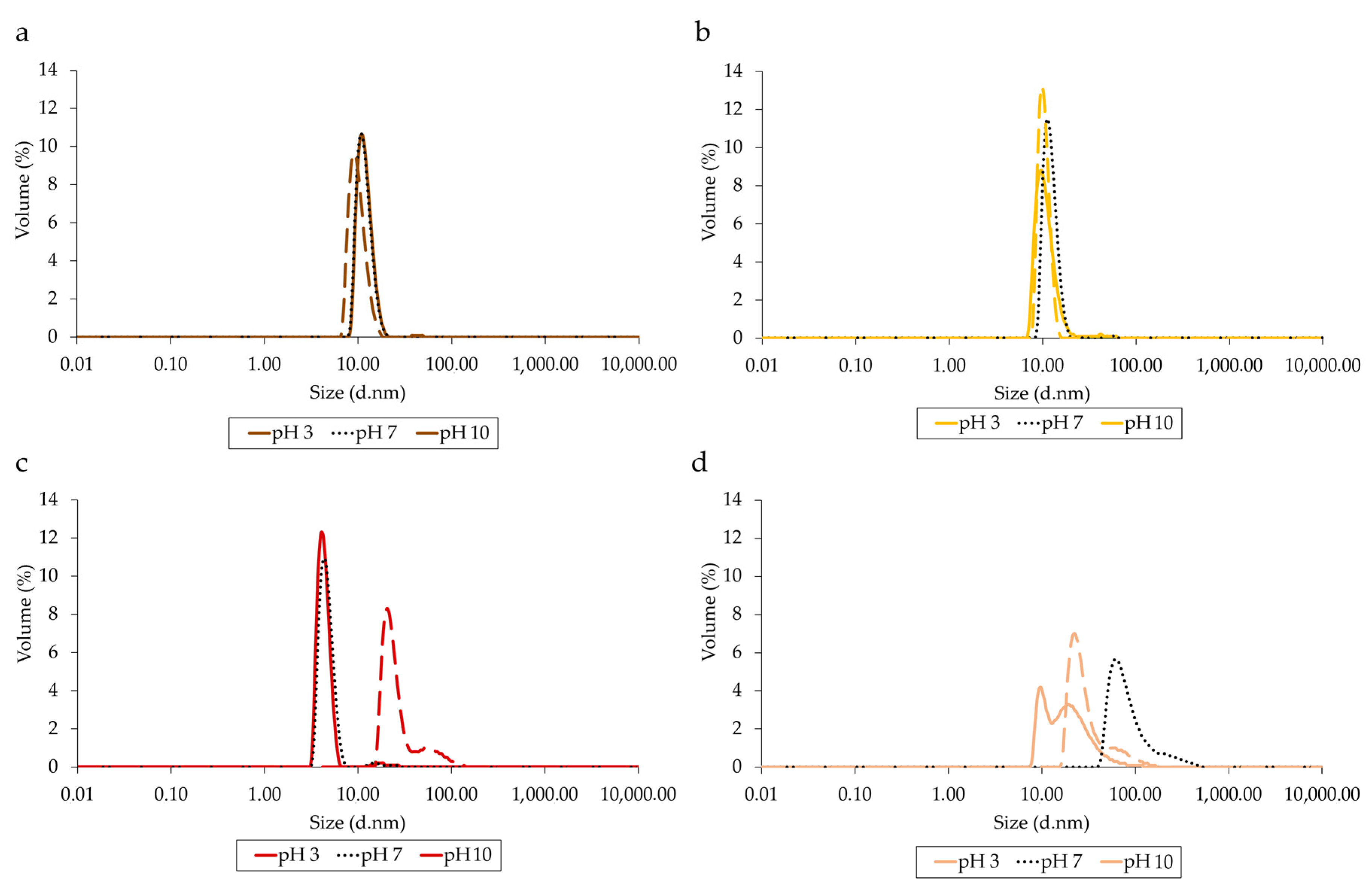
3.4. Structural Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sim, S.Y.J.; Srv, A.; Chiang, J.H.; Henry, C.J. Plant proteins for future foods: A roadmap. Foods 2021, 10, 1967. [Google Scholar] [CrossRef] [PubMed]
- Gueguen, J.; Walrand, S.; Bourgeois, O. Plant proteins: Context and potentialities for human food. Cah. Nutr. Diet. 2016, 51, 177–185. [Google Scholar] [CrossRef]
- Sa, A.G.A.; Moreno, Y.M.F.; Carciofi, B.A.M. Plant proteins as high-quality nutritional source for human diet. Trends Food Sci. Technol. 2020, 97, 170–184. [Google Scholar] [CrossRef]
- GEPV—Bilan de Referencement. 2019. Available online: http://www.lesproteinesvegetales.fr/wp-content/uploads/2020/06/20190627095904_Le_communique_de_presse__Bilan_de_referencement_2019.pdf (accessed on 13 May 2022).
- FAO. FAOSTAT-Production. FAO Statistics Division 2009. 2019. Available online: https://www.fao.org/faostat/fr/#data/QCL (accessed on 10 November 2021).
- Duranti, M.; Consonni, A.; Magni, C.; Sessa, F.; Scarafoni, A. The major proteins of lupin seed: Characterisation and molecular properties for use as functional and nutraceutical ingredients. Trends Food Sci. Technol. 2008, 19, 624–633. [Google Scholar] [CrossRef]
- Jayasena, V.; Chih, H.J.; Nasar-Abbas, S. In Efficient isolation of lupin protein. Food Aust. 2011, 63, 306–309. [Google Scholar]
- Hojilla-Evangelista, M.P.; Sessa, D.J.; Mohamed, A. Functional properties of soybean and lupin protein concentrates produced by ultrafiltration-diafiltration. J. Am. Oil Chem. Soc. 2004, 81, 1153–1157. [Google Scholar] [CrossRef]
- Lo, B.; Kasapis, S.; Farahnaky, A. Lupin protein: Isolation and techno-functional properties, a review. Food Hydrocoll. 2021, 112, 106318. [Google Scholar] [CrossRef]
- Hudson, B.J. (Ed.) New and Developing Sources of Food Proteins; Springer Science & Business Media: London, UK, 1994; pp. 145–194. [Google Scholar]
- Muranyi, I.S.; Volke, D.; Hoffmann, R.; Eisner, P.; Herfellner, T.; Brunnbauer, M.; Koehler, P.; Schweiggert-Weisz, U. Protein distribution in lupin protein isolates from Lupinus angustifolius L. prepared by various isolation techniques. Food Chem. 2016, 207, 6–15. [Google Scholar] [CrossRef]
- Nadal, P.; Canela, N.; Katakis, I.; O’Sullivan, C.K. Extraction, isolation, and characterization of globulin proteins from Lupinus albus. J. Agric. Food Chem. 2011, 59, 2752–2758. [Google Scholar] [CrossRef]
- WHO & UN University. Protein and amino acid requirements in human nutrition. In WHO Technical Report Series; WHO & UN University: Geneva, Switzerland, 2007; p. 935. [Google Scholar]
- Chew, P.G.; Casey, A.J.; Johnson, S.K. Protein quality and physico-functionality of Australian sweet lupin (Lupinus angustifolius cv. Gungurru) protein concentrates prepared by isoelectric precipitation or ultrafiltration. Food Chem. 2003, 83, 575–583. [Google Scholar] [CrossRef]
- El-Adawy, T.A.; Rahma, E.H.; El-Bedawey, A.A.; Gafar, A.F. Nutritional potential and functional properties of sweet and bitter lupin seed protein isolates. Food Chem. 2001, 74, 455–462. [Google Scholar] [CrossRef]
- Mariotti, F.; Pueyo, M.E.; Tomz, D.; Mahe, S. The bioavailability and postprandial utilisation of sweet lupin (Lupinus albus)-flour protein is similar to that of purified soyabean protein in human subjects: A study using intrinsically 15 N-labelled proteins. Br. J. Nutr. 2002, 87, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Agostina, A.; Antonioni, C.; Resta, D.; Arnoldi, A.; Bez, J.; Knauf, U.; Wasche, A. Optimization of a pilot-scale process for producing lupin protein isolates with valuable technological properties and minimum thermal damage. J. Agric. Food Chem. 2006, 54, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Lucas, M.M.; Stoddard, F.L.; Annicchiarico, P.; Frias, J.; Martinez-Villaluenga, C.; Sussmann, D.; Duranti, M.; Seger, A.; Zander, P.M.; Pueyo, J.J. The future of lupin as a protein crop in Europe. Front. Plant Sci. 2015, 6, 705. [Google Scholar] [CrossRef]
- Loveday, S.M. Plant protein ingredients with food functionality potential. Nutr. Bull. 2020, 45, 321–327. [Google Scholar] [CrossRef]
- Fontanari, G.G.; Martins, J.M.; Kobelnik, M.; Pastre, I.A.; Arêas, J.A.G.; Batistuti, J.P.; Fertonani, F.L. Thermal studies on protein isolates of white lupin seeds (Lupinus albus). J. Therm. Anal. Calorim. 2012, 108, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Muranyi, I.S.; Otto, C.; Pickardt, C.; Osen, R.; Koehler, P.; Schweiggert-Weisz, U. Influence of the Isolation Method on the Technofunctional Properties of Protein Isolates from Lupinus angustifolius L. J. Food Sci. 2016, 81, C2656–C2663. [Google Scholar] [CrossRef]
- Wong, A.; Pitts, K.; Jayasena, V.; Johnson, S. Isolation and foaming functionality of acid-soluble protein from lupin (Lupinus angustifolius) kernels. J. Sci. Food Agric. 2013, 93, 3755–3762. [Google Scholar] [CrossRef]
- Muranyi, I.S.; Otto, C.; Pickardt, C.; Koehler, P.; Schweiggert-Weisz, U. Microscopic characterisation and composition of proteins from lupin seed (Lupinus angustifolius L.) as affected by the isolation procedure. Food Res. Int. 2013, 54, 1419–1429. [Google Scholar] [CrossRef]
- Sussmann, D.; Pickardt, C.; Schweiggert, U.; Eisner, P. Influence of different processing parameters on the isolation of lupin (Lupinus angustifolius L.) protein isolates: A preliminary study. J. Food Process Eng. 2013, 36, 18–28. [Google Scholar] [CrossRef]
- AOAC. Method 991.20. Nitrogen (Total) in Milk. In Official Methods of Analysis of the Association of Official Analytical Chemists; Association of Official Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Sussmann, D.; Halter, T.; Pickardt, C.; Schweiggert-Weisz, U.; Eisner, P. An optimization approach for the production of fatlike protein isolates from different leguminous seeds using response surface methodology. J. Food Process Eng. 2013, 36, 715–730. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680. [Google Scholar] [CrossRef] [PubMed]
- Albe-Slabi, S.; Mathe, C.; Basselin, M.; Fournier, F.; Aymes, A.; Bianeis, M.; Galet, O.; Kapel, R. Optimization of sunflower albumin extraction from oleaginous meal and characterization of their structure and properties. Food Hydrocoll. 2020, 99, 105335. [Google Scholar] [CrossRef]
- Vioque, J.; Sanchez-Vioque, R.; Clemente, A.; Pedroche, J.; Millan, F. Partially hydrolyzed rapeseed protein isolates with improved functional properties. J. Am. Oil Chem. Soc. 2000, 77, 447–450. [Google Scholar] [CrossRef]
- Albe-Slabi, S.; Mathe, C.; Basselin, M.; Framboisier, X.; Ndiaye, M.; Galet, O.; Kapel, R. Multi-objective optimization of solid/liquid extraction of total sunflower proteins from cold press meal. Food Chem. 2020, 317, 126423. [Google Scholar] [CrossRef]
- Albe-Slabi, S.; Defaix, C.; Beaubier, S.; Galet, O.; Kapel, R. Selective extraction of napins: Process optimization and impact on structural and functional properties. Food Hydrocoll. 2022, 122, 107105. [Google Scholar] [CrossRef]
- Nioi, C.; Kapel, R.; Rondags, E.; Marc, I. Selective extraction, structural characterization and antifungal activity assessment of napins from an industrial rapeseed meal. Food Chem. 2012, 134, 2149–2155. [Google Scholar] [CrossRef]
- Sironi, E.; Sessa, F.; Duranti, M. A simple procedure of lupin seed protein fractionation for selective food applications. Eur. Food Res. Technol. 2005, 221, 145–150. [Google Scholar] [CrossRef]
- Wanasundara, J.P.D.; McIntosh, T.C. Process of Aqueous Protein Extraction from Brassicaceae Oilseeds. U.S. Patent No. 8,557,963, 15 October 2013. [Google Scholar]
- Gonzalez-Perez, S.; Vereijken, J.M.; Merck, K.B.; van Koningsveld, G.A.; Gruppen, H.; Voragen, A.G. Conformational states of sunflower (Helianthus annuus) helianthinin: Effect of heat and pH. J. Agric. Food Chem. 2004, 52, 6770–6778. [Google Scholar] [CrossRef]
- Berghout, J.A.M.; Boom, R.M.; Van der Goot, A.J. The potential of aqueous fractionation of lupin seeds for high-protein foods. Food Chem. 2014, 159, 64–70. [Google Scholar] [CrossRef]
- Berghout, J.A.M.; Venema, P.; Boom, R.M.; Van der Goot, A.J. Comparing functional properties of concentrated protein isolates with freeze-dried protein isolates from lupin seeds. Food Hydrocoll. 2015, 51, 346–354. [Google Scholar] [CrossRef]
- Boye, J.I.; Aksay, S.; Roufik, S.; Ribereau, S.; Mondor, M.; Farnworth, E.; Rajamohamed, S.H. Comparison of the functional properties of pea, chickpea and lentil protein concentrates processed using ultrafiltration and isoelectric precipitation techniques. Food Res. Int. 2010, 43, 537–546. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Ereifej, K.; Gammoh, S.; Kubow, S.; Tawalbeh, D. Preparation of mayonnaise from extracted plant protein isolates of chickpea, broad bean and lupin flour: Chemical, physiochemical, nutritional and therapeutic properties. J. Food Sci. Technol. 2017, 54, 1395–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozani, S.; Doxastakis, G.; Kiosseoglou, V. Functionality of lupin seed protein isolate in relation to its interfacial behaviour. Food Hydrocoll. 2002, 16, 241–247. [Google Scholar] [CrossRef]
- Makri, E.; Papalamprou, E.; Doxastakis, G. Study of functional properties of seed storage proteins from indigenous European legume crops (lupin, pea, broad bean) in admixture with polysaccharides. Food Hydrocoll. 2005, 19, 583–594. [Google Scholar] [CrossRef]
- Lilley, G.G. The subunit structure and stability of conglutin δ, a sulphurrich protein from the seeds of Lupinus angustifolius L. J. Sci. Food Agric. 1986, 37, 895–907. [Google Scholar] [CrossRef]
- Alonso-Miravalles, L.; Jeske, S.; Bez, J.; Detzel, A.; Busch, M.; Krueger, M.; Lrissa Wriessnegger, C.; O’Mahony, J.A.; Zannini, E.; Arendt, E.K. Membrane filtration and isoelectric precipitation technological approaches for the preparation of novel, functional and sustainable protein isolate from lentils. Eur. Food Res. Technol. 2019, 245, 1855–1869. [Google Scholar] [CrossRef]
- Czubinski, J. Insight into thermally induced structural changes of lupin seed γ-conglutin. Food Chem. 2021, 354, 129480. [Google Scholar] [CrossRef]
- Sirtori, E.; Resta, D.; Brambilla, F.; Zacherl, C.; Arnoldi, A. The effects of various processing conditions on a protein isolate from Lupinus angustifolius. Food Chem. 2010, 120, 496–504. [Google Scholar] [CrossRef]
- Sousa, I.M.; Mitchell, J.R.; Ledward, D.A.; Hill, S.E.; da Costa, M.L.B. Differential scanning calorimetry of lupin and soy proteins. Z. Fur Lebensm.-Unters. Und Forsch. 1995, 201, 566–569. [Google Scholar] [CrossRef]
- Aguilar-Acosta, L.A.; Serna-Saldivar, S.O.; Rodríguez-Rodríguez, J.; Escalante-Aburto, A.; Chuck-Hernández, C. Effect of ultrasound application on protein yield and fate of alkaloids during lupin alkaline extraction process. Biomolecules 2020, 10, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Múzquiz, M.; Fernández, E.G.; Juana, C.B.; Pacual, H.; Cabellos, B.; Hoyo, C.C.; Pedrosa, M.M. Chemical composition of a new Lupinus species found in Spain, Lupinus mariae-josephi H. Pascual (Fabaceae). Span. J. Agric. Res. 2011, 9, 1233–1244. [Google Scholar] [CrossRef]
- González-Pérez, S.; Vereijken, J.M. Sunflower proteins: Overview of their physicochemical, structural and functional properties. J. Sci. Food Agric. 2007, 87, 2173–2191. [Google Scholar] [CrossRef]
- Folawiyo, Y.L.; Apenten, R.O. The effect of heat-and acid-treatment on the structure of rapeseed albumin (napin). Food Chem. 1997, 58, 237–243. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Membrane Cut-Off (kDa) | Retention Rate of Proteins | Flux (mL/min/cm2) |
|---|---|---|
| 10 | 1.00 | 0.09 |
| 30 | 0.99 | 0.09 |
| 100 | 0.99 | 0.09 |
| 300 | 0.97 | 0.11 |
| Process Yields | Lupin Protein Isolate | |||
|---|---|---|---|---|
| LPI-pH7/IP | LPI-pH7/UF | LPI-pH2/IP | LPI-pH2/UF | |
| Extraction yield (%) | 40.4 | 40.4 | 37.1 | 37.1 |
| Purification yield (%) | 66.5 | 72.5 | 75.8 | 76.1 |
| Protein recovery (%) | 30.2 | 32.2 | 28.1 | 31.3 |
| Lupin Protein Isolate | Parameters of CIE L*a*b* Scale | ||
|---|---|---|---|
| L* | a* | b* | |
| LP-pH7/IP | 100.8 | 1.4 | 9.6 |
| LP-pH7/UF | 101.8 | 3.1 | 5.2 |
| LP-pH2/IP | 98.0 | -0.2 | 8.1 |
| LP-pH2/UF | 101.0 | 2.8 | 2.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albe-Slabi, S.; Mesieres, O.; Mathé, C.; Ndiaye, M.; Galet, O.; Kapel, R. Combined Effect of Extraction and Purification Conditions on Yield, Composition and Functional and Structural Properties of Lupin Proteins. Foods 2022, 11, 1646. https://doi.org/10.3390/foods11111646
Albe-Slabi S, Mesieres O, Mathé C, Ndiaye M, Galet O, Kapel R. Combined Effect of Extraction and Purification Conditions on Yield, Composition and Functional and Structural Properties of Lupin Proteins. Foods. 2022; 11(11):1646. https://doi.org/10.3390/foods11111646
Chicago/Turabian StyleAlbe-Slabi, Sara, Odile Mesieres, Christelle Mathé, Mbalo Ndiaye, Olivier Galet, and Romain Kapel. 2022. "Combined Effect of Extraction and Purification Conditions on Yield, Composition and Functional and Structural Properties of Lupin Proteins" Foods 11, no. 11: 1646. https://doi.org/10.3390/foods11111646