
Chemosensitization Effect of Seabuckthorn (Hippophae rhamnoides L.) Pulp Oil via Autophagy and Senescence in NSCLC Cells



Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seabuckthorn Active Compounds | Medicinal Properties | References | |
|---|---|---|---|
| 1. | Tocopherols | Lipid-soluble antioxidant, inhibits oxidative stress, promising diet for Alzheimer’s disease (AD) prevention, may lower cholesterol levels | [7,26] |
| 2. | Carotenoids | Beneficial antioxidant, wound healing through stimulation of angiogenesis, collagen synthesis, epithelialization | [27] |
| 3. | Vitamin C | Antioxidant, acts as an enzymatic cofactor and maintains tissue integrity, and accelerates the formation of skin, epithelial, and endothelial barriers and collagen synthesis | [28,29] |
| 4. | Vitamin B complex (B1, B2, and B6) | Increases the regeneration of axons and neuronal survival, promotes cell repair | [16] |
| 5. | Polyphenolic compounds | Antioxidant, cytoprotective, hepatoprotective, wound healing | [13,14] |
| 6. | Phytosterols | Stimulates microcirculation in the skin, exerts anti-ulcer, anti-atherogenic, and antitumor effects, modulation of the inflammatory process | [11] |
| 7. | Polyunsaturated fatty acids (PUFA) | Immunomodulatory, neuroprotective, anticancer | [30] |
| 8. | Zinc | Capable of improving the blood circulation, anticancer effect, cofactor for a number of enzymes, enhances the utilization of vitamin A | [31] |
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. Preparation of Sea Buckthorn Pulp Oil
2.3. Cell Culture
2.4. Cell Cytotoxicity Assay
2.5. Evaluation of Cell Morphology
2.6. Drug Synergy
2.7. Cell Cycle Analysis
2.8. Intracellular Reactive Oxygen Species (ROS) Level Detection
2.9. Senescence-Associated-β-Galactosidase Staining
2.10. Immunocytochemistry
2.11. Western Blot
2.12. Liquid Chromatographic (LC)-Tandem Mass Spectrometric (MS/MS) Analysis
2.13. Statistical Analysis
3. Results
3.1. SBO Inhibited NSCLC Cells Viability
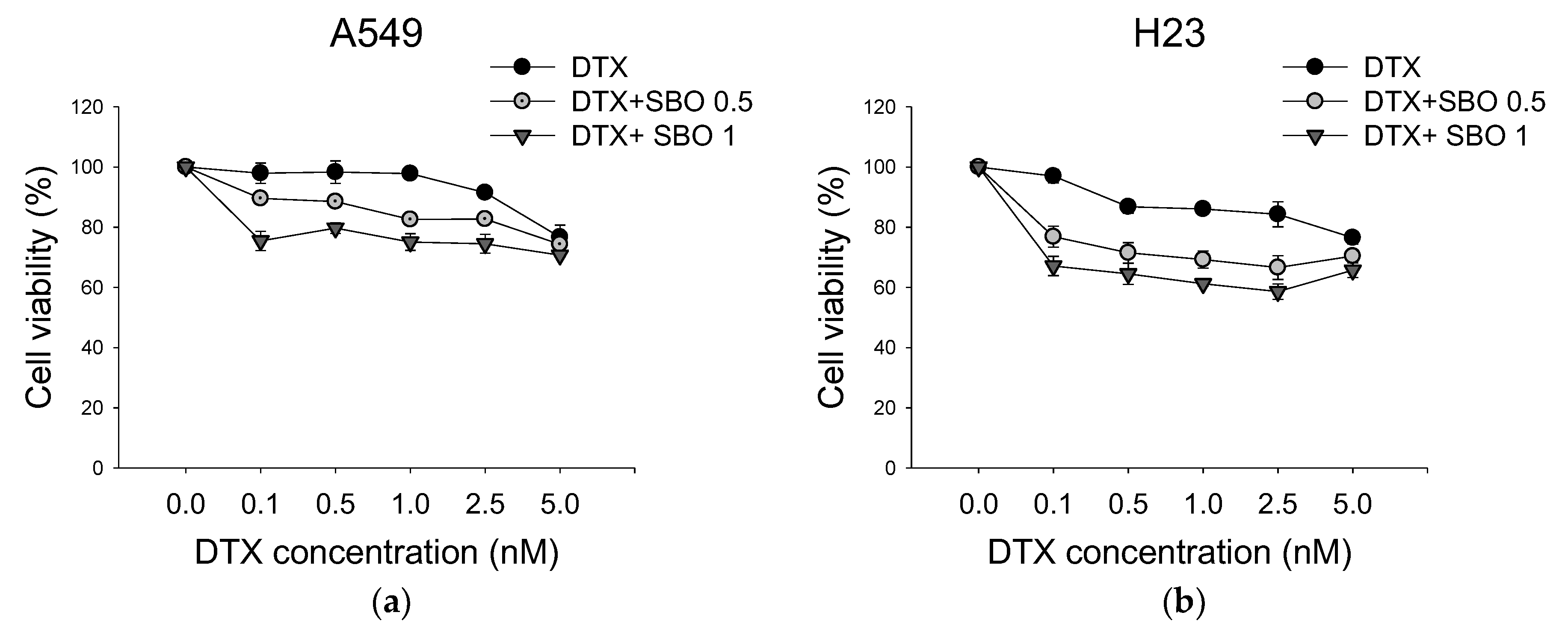
3.2. Synergistic Effect of SBO Combined with DTX in NSCLC
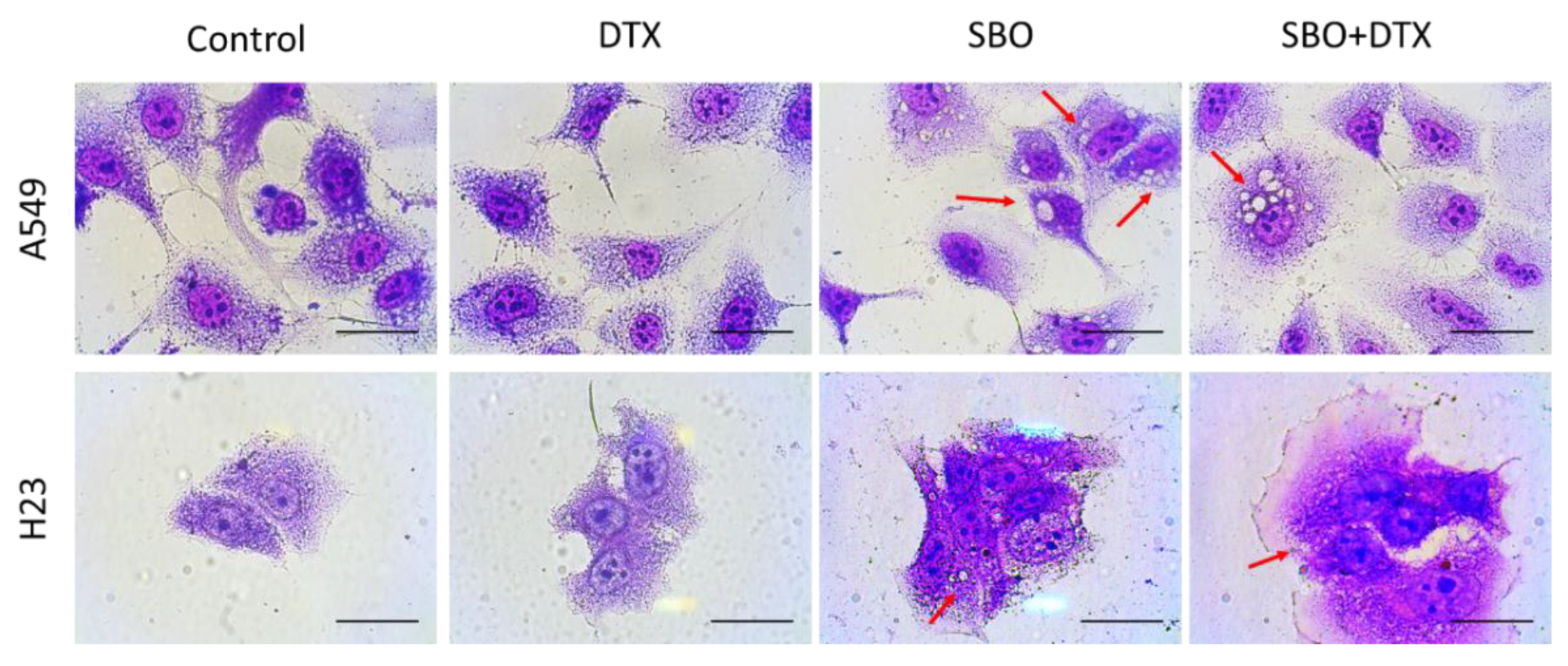
3.3. SBO Induces Morphological Changes in NSCLC Cells
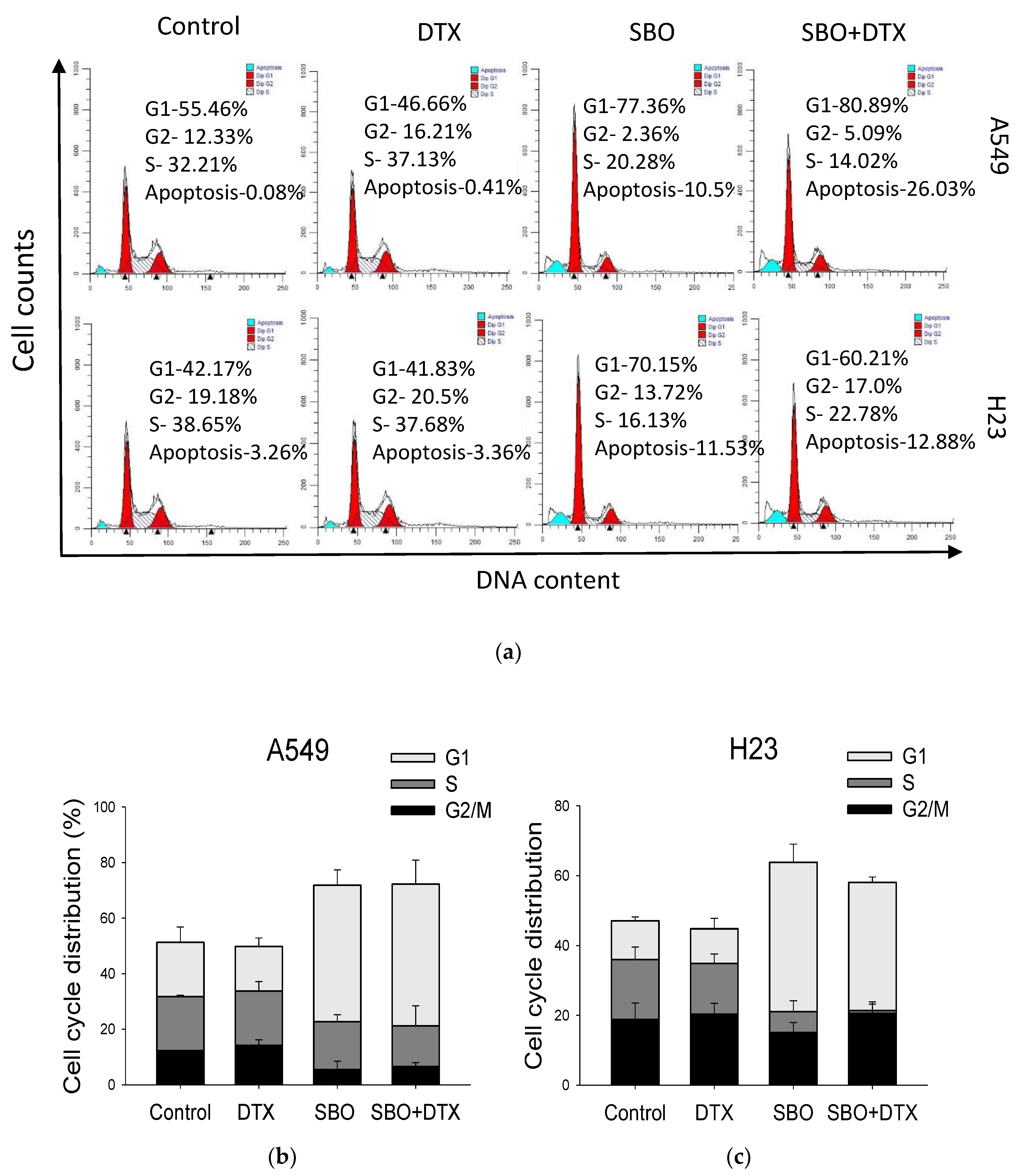
3.4. Synergizing Effect of SBO Induced by G1 Phase Cell Cycle Arrest and Senescence
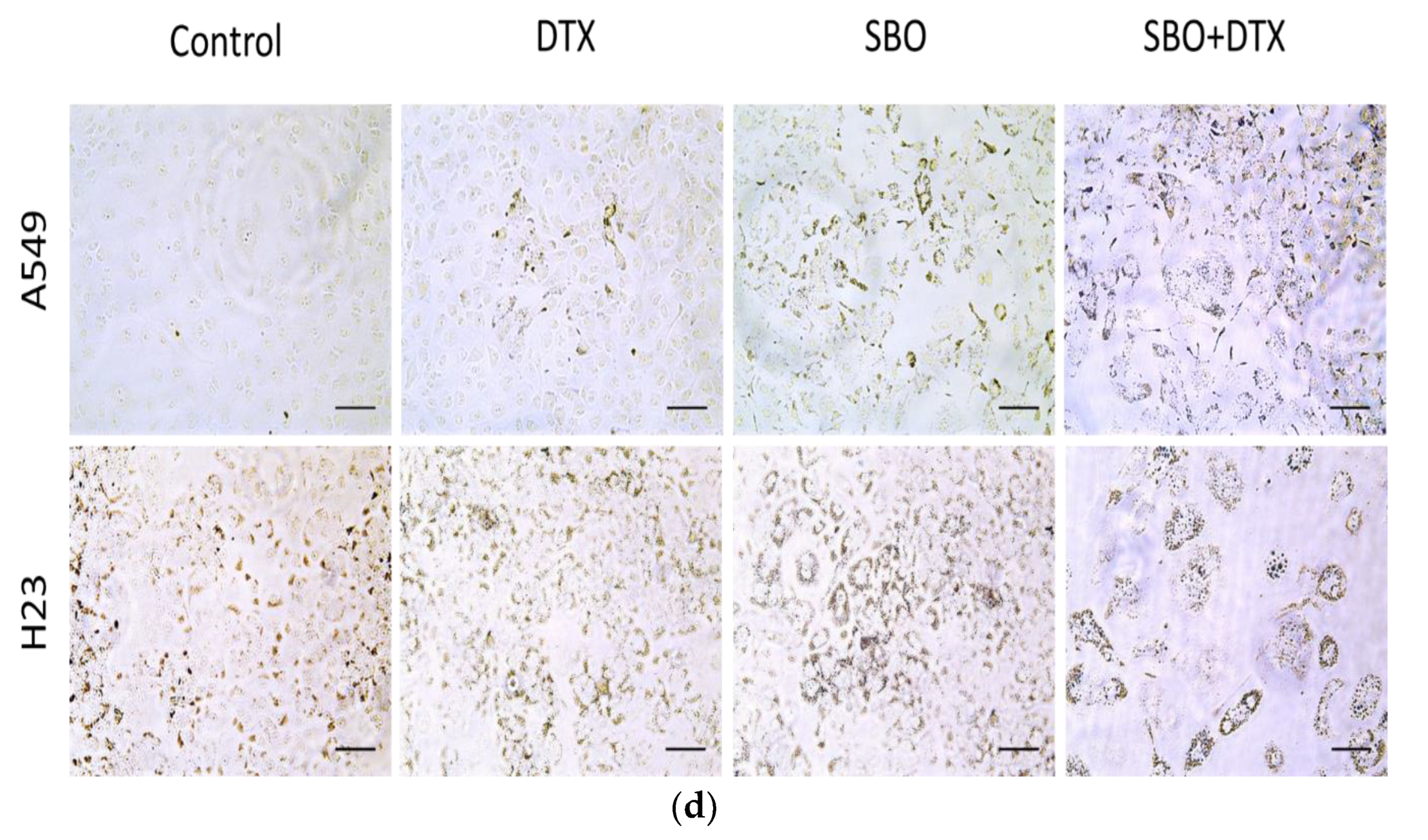
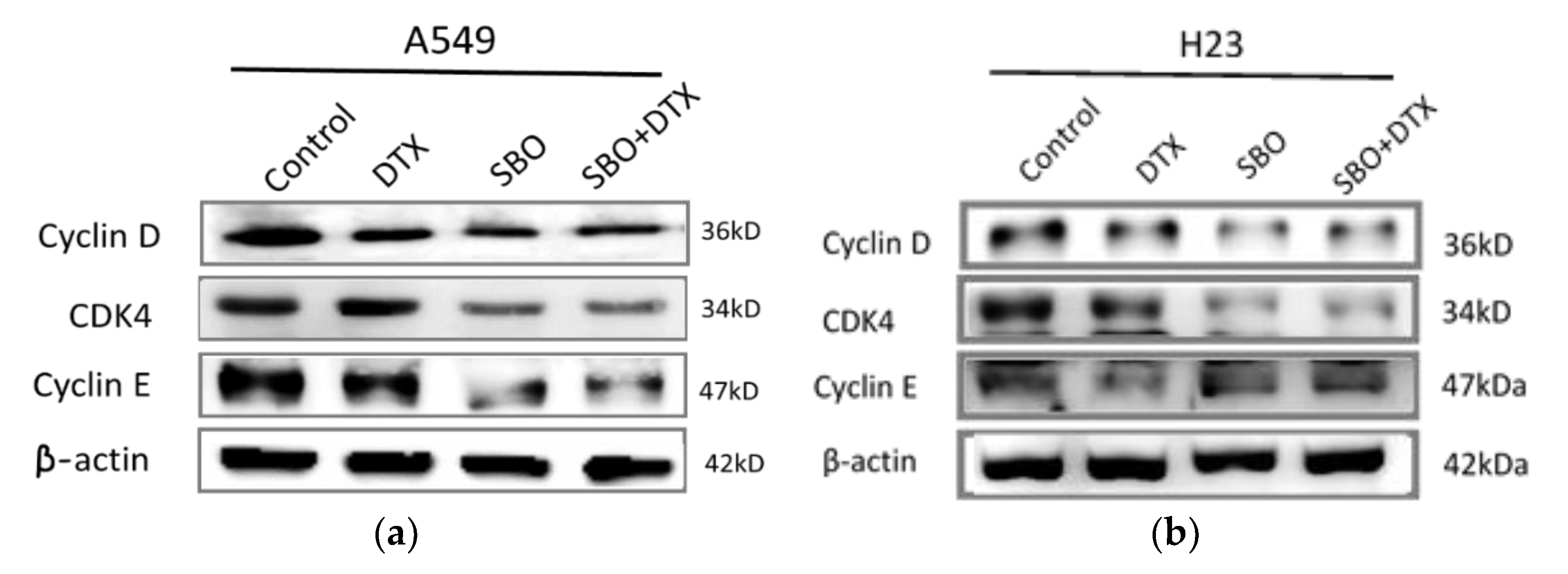
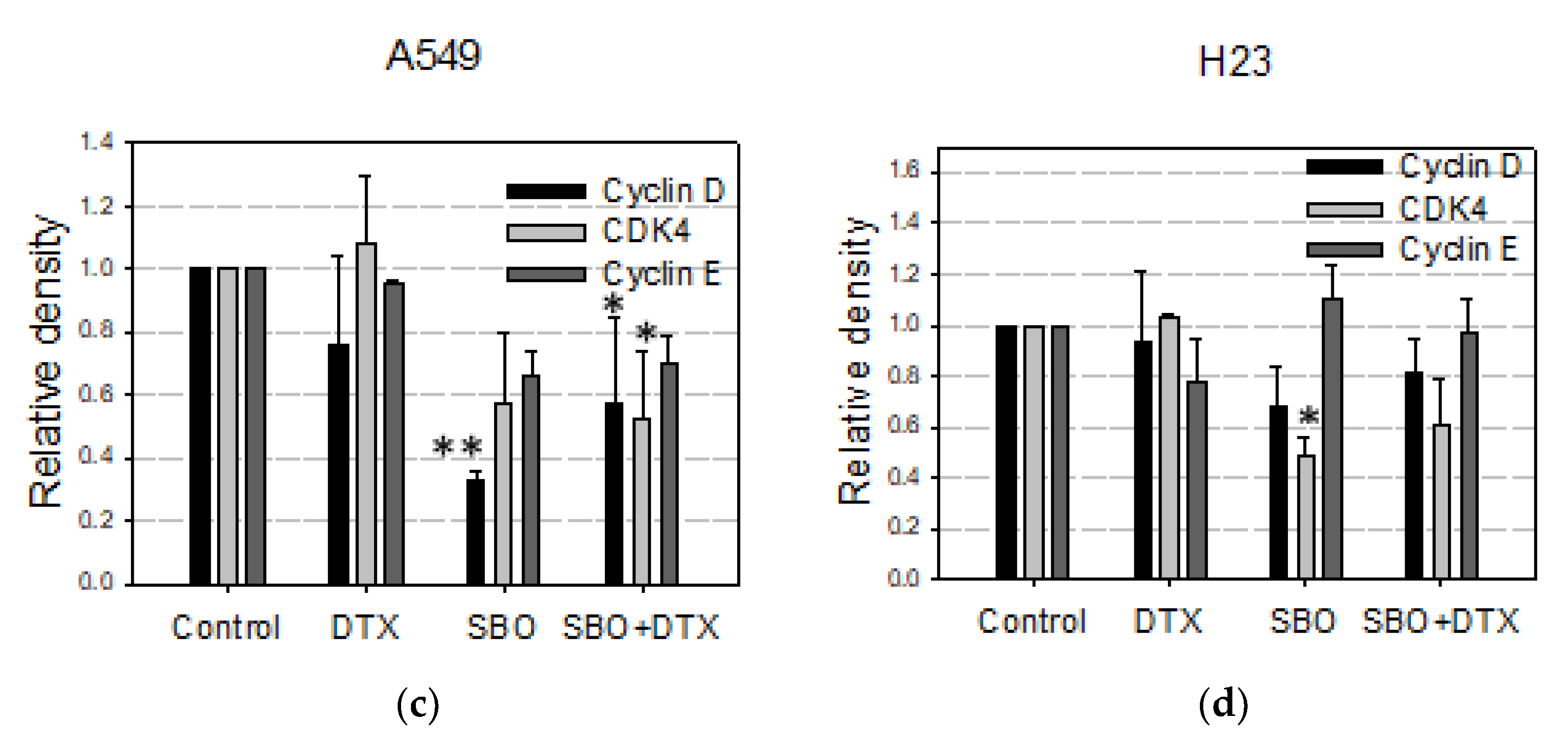
3.5. SBO Synergizes DTX Effect via Alteration in Proteins Associated with G1 Arrest and Senescence in NSCLC Cells
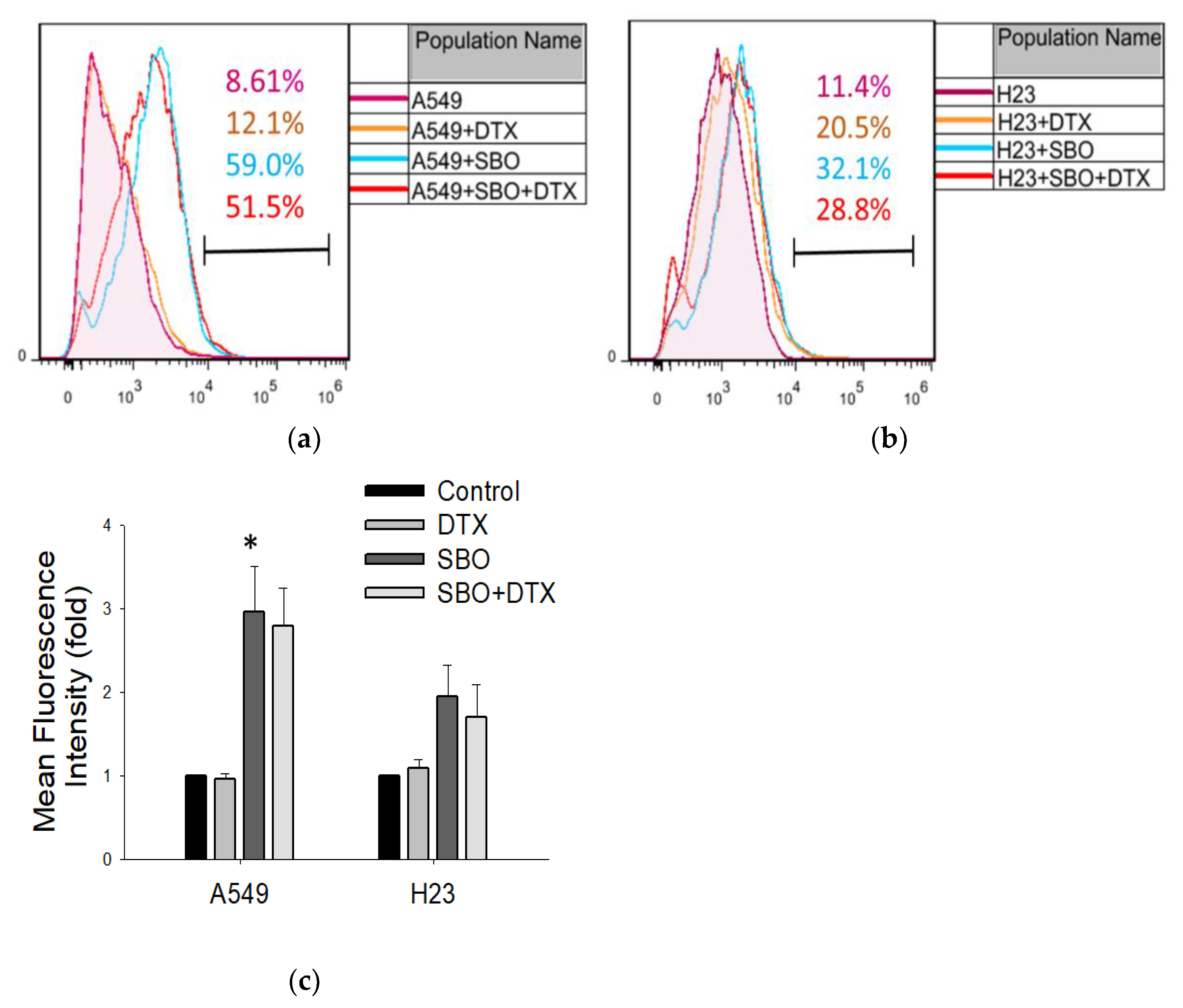
3.6. SBO Induced the Production of Reactive Oxygen Species (ROS) in NSCLC
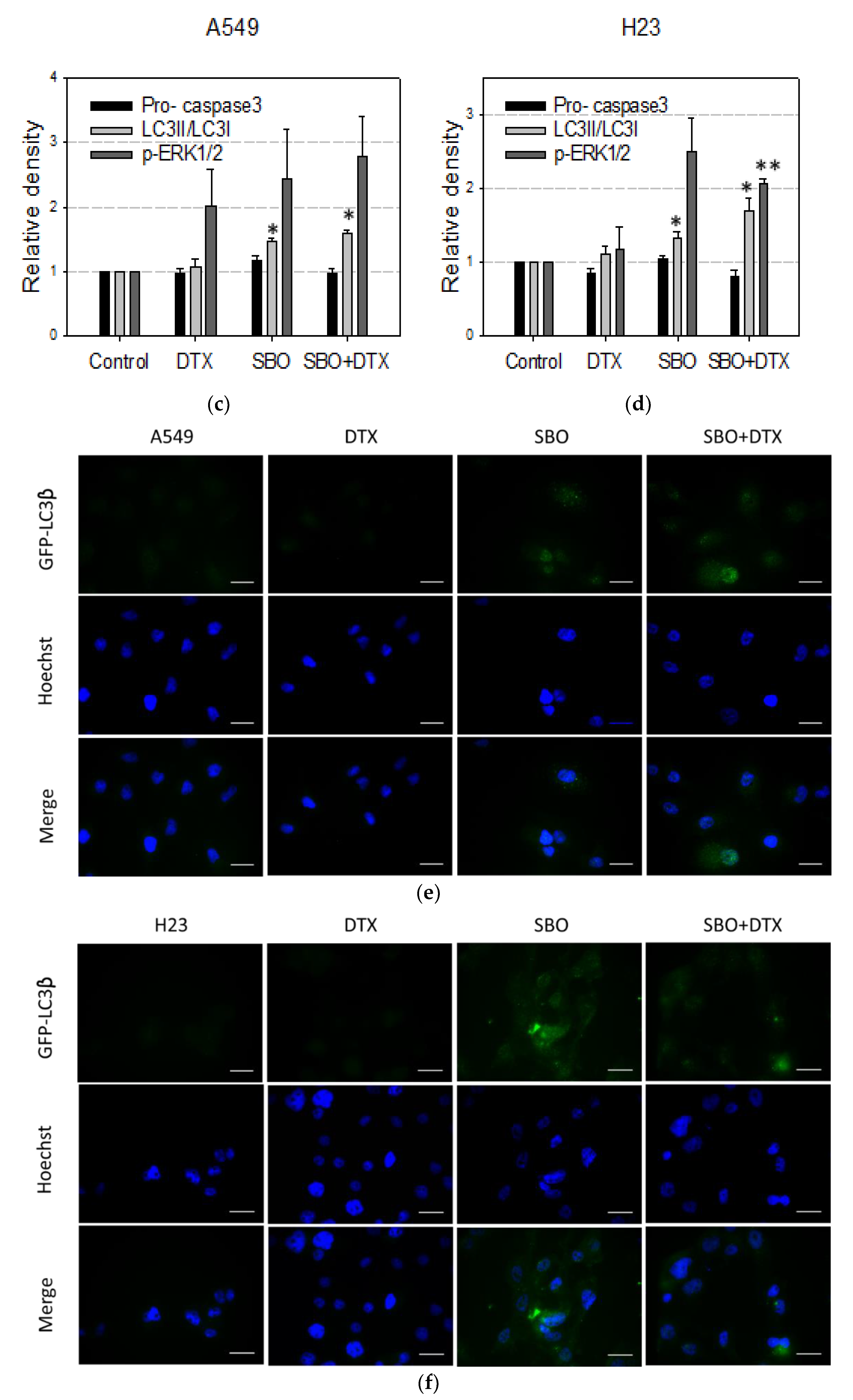
3.7. Non-Apoptotic Autophagy Is Involved in SBO Induced Synergizing Effect Possibly via Sustained ERK Phosphorylation
3.8. Chemical Profile of SBO
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Zarogoulidis, K.; Zarogoulidis, P.; Darwiche, K.; Boutsikou, E.; Machairiotis, N.; Tsakiridis, K.; Katsikogiannis, N.; Kougioumtzi, I.; Karapantzos, I.; Huang, H.; et al. Treatment of non-small cell lung cancer (NSCLC). J. Thorac. Dis 2013, 5 (Suppl. S4), S389–S396. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Bahadur, S.; Harwansh, R.K.; Biswas, S.; Banerjee, S. Paradigm shift in natural product research: Traditional medicine inspired approaches. Phytochem. Rev. 2017, 16, 803–826. [Google Scholar] [CrossRef]
- El-Readi, M.Z.; Al-Abd, A.M.; Althubiti, M.A.; Almaimani, R.A.; Al-Amoodi, H.S.; Ashour, M.L.; Wink, M.; Eid, S.Y. Multiple Molecular Mechanisms to Overcome Multidrug Resistance in Cancer by Natural Secondary Metabolites. Front. Pharmacol. 2021, 12, 658513. [Google Scholar] [CrossRef]
- Li, T.S.; Schroeder, W. Sea buckthorn (Hippophae rhamnoides L.): A multipurpose plant. HortTechnology 1996, 6, 370–380. [Google Scholar] [CrossRef]
- Christaki, E. Hippophae rhamnoides L. (Sea Buckthorn): A potential source of nutraceuticals. Food Public Health 2012, 2, 69–72. [Google Scholar] [CrossRef]
- Andersson, S.C.; Rumpunen, K.; Johansson, E.; Olsson, M.E. Tocopherols and tocotrienols in sea buckthorn (Hippophae rhamnoides L.) berries during ripening. J. Agric. Food Chem. 2008, 56, 6701–6706. [Google Scholar] [CrossRef]
- Guliyev, V.B.; Gul, M.; Yildirim, A. Hippophae rhamnoides L.: Chromatographic methods to determine chemical composition, use in traditional medicine and pharmacological effects. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2004, 812, 291–307. [Google Scholar] [CrossRef]
- Geetha, S.; Jayamurthy, P.; Pal, K.; Pandey, S.; Kumar, R.; Sawhney, R. Hepatoprotective effects of sea buckthorn (Hippophae rhamnoides L.) against carbon tetrachloride induced liver injury in rats. J. Sci. Food Agric. 2008, 88, 1592–1597. [Google Scholar] [CrossRef]
- Grey, C.; Widén, C.; Adlercreutz, P.; Rumpunen, K.; Duan, R.-D. Antiproliferative effects of sea buckthorn (Hippophae rhamnoides L.) extracts on human colon and liver cancer cell lines. Food Chem. 2010, 120, 1004–1010. [Google Scholar] [CrossRef]
- Li, T.S.; Beveridge, T.H.; Drover, J.C. Phytosterol content of sea buckthorn (Hippophae rhamnoides L.) seed oil: Extraction and identification. Food Chem. 2007, 101, 1633–1639. [Google Scholar] [CrossRef]
- Chaman, S.; Syed, N.I.; Danish, Z.; Khan, F.Z. Phytochemical analysis, antioxidant and antibacterial effects of sea buckthorn berries. Pak. J. Pharm. Sci. 2011, 24, 345–351. [Google Scholar]
- Gupta, A.; Kumar, R.; Pal, K.; Singh, V.; Banerjee, P.K.; Sawhney, R.C. Influence of sea buckthorn (Hippophae rhamnoides L.) flavone on dermal wound healing in rats. Mol. Cell Biochem. 2006, 290, 193–198. [Google Scholar] [CrossRef]
- Maheshwari, D.T.; Yogendra Kumar, M.S.; Verma, S.K.; Singh, V.K.; Singh, S.N. Antioxidant and hepatoprotective activities of phenolic rich fraction of Seabuckthorn (Hippophae rhamnoides L.) leaves. Food Chem. Toxicol. 2011, 49, 2422–2428. [Google Scholar] [CrossRef]
- Enkhtaivan, G.; Maria John, K.M.; Pandurangan, M.; Hur, J.H.; Leutou, A.S.; Kim, D.H. Extreme effects of Seabuckthorn extracts on influenza viruses and human cancer cells and correlation between flavonol glycosides and biological activities of extracts. Saudi J. Biol. Sci. 2017, 24, 1646–1656. [Google Scholar] [CrossRef] [Green Version]
- Olas, B. Sea buckthorn as a source of important bioactive compounds in cardiovascular diseases. Food Chem. Toxicol. 2016, 97, 199–204. [Google Scholar] [CrossRef]
- Davies, A.M.; Lara, P.N., Jr.; Mack, P.C.; Gandara, D.R. Docetaxel in non-small cell lung cancer: A review. Expert Opin. Pharm. 2003, 4, 553–565. [Google Scholar] [CrossRef]
- Fossella, F.V.; Lee, J.S.; Berille, J.; Hong, W.K. Summary of phase II data of docetaxel (Taxotere), an active agent in the first- and second-line treatment of advanced non-small cell lung cancer. Semin. Oncol. 1995, 22, 22–29. [Google Scholar]
- Georgoulias, V.; Ardavanis, A.; Agelidou, A.; Agelidou, M.; Chandrinos, V.; Tsaroucha, E.; Toumbis, M.; Kouroussis, C.; Syrigos, K.; Polyzos, A.; et al. Docetaxel versus docetaxel plus cisplatin as front-line treatment of patients with advanced non-small-cell lung cancer: A randomized, multicenter phase III trial. J. Clin. Oncol. 2004, 22, 2602–2609. [Google Scholar] [CrossRef] [Green Version]
- Seidman, R.; Gitelman, I.; Sagi, O.; Horwitz, S.B.; Wolfson, M. The role of ERK 1/2 and p38 MAP-kinase pathways in taxol-induced apoptosis in human ovarian carcinoma cells. Exp. Cell Res. 2001, 268, 84–92. [Google Scholar] [CrossRef]
- Boldt, S.; Weidle, U.H.; Kolch, W. The role of MAPK pathways in the action of chemotherapeutic drugs. Carcinogenesis 2002, 23, 1831–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Nigro, C.; Maffi, M.; Fischel, J.L.; Formento, P.; Milano, G.; Merlano, M. The combination of docetaxel and the somatostatin analogue lanreotide on androgen-independent docetaxel-resistant prostate cancer: Experimental data. BJU Int. 2008, 102, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Cagnol, S.; Chambard, J.C. ERK and cell death: Mechanisms of ERK-induced cell death--apoptosis, autophagy and senescence. FEBS J. 2010, 277, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, S.; Jain, K.; Basu, A. Regulation of autophagy by kinases. Cancers 2011, 3, 2630–2654. [Google Scholar] [CrossRef]
- Slobodnyuk, K.; Radic, N.; Ivanova, S.; Llado, A.; Trempolec, N.; Zorzano, A.; Nebreda, A.R. Autophagy-induced senescence is regulated by p38alpha signaling. Cell Death Dis. 2019, 10, 376. [Google Scholar] [CrossRef]
- Berman, K.; Brodaty, H. Tocopherol (vitamin E) in Alzheimer’s disease and other neurodegenerative disorders. CNS Drugs 2004, 18, 807–825. [Google Scholar] [CrossRef]
- Pop, R.M.; Weesepoel, Y.; Socaciu, C.; Pintea, A.; Vincken, J.P.; Gruppen, H. Carotenoid composition of berries and leaves from six Romanian sea buckthorn (Hippophae rhamnoides L.) varieties. Food Chem. 2014, 147, 1–9. [Google Scholar] [CrossRef]
- Gutzeit, D.; Baleanu, G.; Winterhalter, P.; Jerz, G. Vitamin C content in sea buckthorn berries (Hippophae rhamnoides L. ssp. rhamnoides) and related products: A kinetic study on storage stability and the determination of processing effects. J. Food Sci. 2008, 73, C615–C620. [Google Scholar] [CrossRef]
- DePhillipo, N.N.; Aman, Z.S.; Kennedy, M.I.; Begley, J.P.; Moatshe, G.; LaPrade, R.F. Efficacy of Vitamin C Supplementation on Collagen Synthesis and Oxidative Stress after Musculoskeletal Injuries: A Systematic Review. Orthop. J. Sports Med. 2018, 6, 2325967118804544. [Google Scholar] [CrossRef]
- Cakir, A. Essential oil and fatty acid composition of the fruits of Hippophae rhamnoides L. (Sea Buckthorn) and Myrtus communis L. from Turkey. Biochem. Syst. Ecol. 2004, 32, 809–816. [Google Scholar] [CrossRef]
- Suryakumar, G.; Gupta, A. Medicinal and therapeutic potential of Sea buckthorn (Hippophae rhamnoides L.). J. Ethnopharmacol. 2011, 138, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Houghton, P.; Fang, R.; Techatanawat, I.; Steventon, G.; Hylands, P.J.; Lee, C.C. The sulphorhodamine (SRB) assay and other approaches to testing plant extracts and derived compounds for activities related to reputed anticancer activity. Methods 2007, 42, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawless, C.; Wang, C.; Jurk, D.; Merz, A.; Zglinicki, T.; Passos, J.F. Quantitative assessment of markers for cell senescence. Exp. Gerontol 2010, 45, 772–778. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- El Hasasna, H.; Athamneh, K.; Al Samri, H.; Karuvantevida, N.; Al Dhaheri, Y.; Hisaindee, S.; Ramadan, G.; Al Tamimi, N.; AbuQamar, S.; Eid, A.; et al. Rhus coriaria induces senescence and autophagic cell death in breast cancer cells through a mechanism involving p38 and ERK1/2 activation. Sci. Rep. 2015, 5, 13013. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Lee, J.S. Cellular senescence: A promising strategy for cancer therapy. BMB Rep. 2019, 52, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Musgrove, E.A.; Caldon, C.E.; Barraclough, J.; Stone, A.; Sutherland, R.L. Cyclin D as a therapeutic target in cancer. Nat. Rev. Cancer 2011, 11, 558–572. [Google Scholar] [CrossRef]
- Azad, M.B.; Chen, Y.; Gibson, S.B. Regulation of autophagy by reactive oxygen species (ROS): Implications for cancer progression and treatment. Antioxid Redox Signal. 2009, 11, 777–790. [Google Scholar] [CrossRef]
- Lu, T.; Finkel, T. Free radicals and senescence. Exp. Cell Res. 2008, 314, 1918–1922. [Google Scholar] [CrossRef]
- Masoodi, K.Z.; Wani, W.; Dar, Z.A.; Mansoor, S.; Anam-ul-Haq, S.; Farooq, I.; Hussain, K.; Wani, S.A.; Nehvi, F.A.; Ahmed, N. Sea buckthorn (Hippophae rhamnoides L.) inhibits cellular proliferation, wound healing and decreases expression of prostate specific antigen in prostate cancer cells in vitro. J. Funct. Foods 2020, 73, 104102. [Google Scholar] [CrossRef]
- Scherz-Shouval, R.; Elazar, Z. ROS, mitochondria and the regulation of autophagy. Trends Cell Biol. 2007, 17, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.U.; Tan, K.B.; Lin, H.; Chiu, G.N. The role of reactive oxygen species and autophagy in safingol-induced cell death. Cell Death Dis. 2011, 2, e129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.J.; Kim, S.Y.; Kwon, E.B.; Jo, Y.H.; Lee, M.K.; Lee, H.S.; Moon, D.O.; Kim, M.O. Derrone induces autophagic cell death through induction of ROS and ERK in A549 cells. PLoS ONE 2019, 14, e0218659. [Google Scholar] [CrossRef] [PubMed]
- Olas, B.; Skalski, B.; Ulanowska, K. The Anticancer Activity of Sea Buckthorn [Elaeagnus rhamnoides (L.) A. Nelson]. Front. Pharmacol 2018, 9, 232. [Google Scholar] [CrossRef] [Green Version]
- Roesly, H.B.; Khan, M.R.; Chen, H.D.; Hill, K.A.; Narendran, N.; Watts, G.S.; Chen, X.; Dvorak, K. The decreased expression of Beclin-1 correlates with progression to esophageal adenocarcinoma: The role of deoxycholic acid. Am. J. Physiol. Gastrointest Liver Physiol. 2012, 302, G864–G872. [Google Scholar] [CrossRef]
- Ma, Y.; Wink, M. Lobeline, a piperidine alkaloid from Lobelia can reverse P-gp dependent multidrug resistance in tumor cells. Phytomedicine 2008, 15, 754–758. [Google Scholar] [CrossRef]
- Chen, J.; Shen, L.; Zhou, R.; Yao, W.; Zhong, M.; Zhu, Z.H.; Zeng, S. Reversal effect and mechanism of lobeline on the multidrug-resistance of human breast cancer cells MCF-7/ADM. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2009, 34, 738–743. [Google Scholar]
- Sharma, N.; Kumar, A.; Sharma, P.R.; Qayum, A.; Singh, S.K.; Dutt, P.; Paul, S.; Gupta, V.; Verma, M.K.; Satti, N.K.; et al. A new clerodane furano diterpene glycoside from Tinospora cordifolia triggers autophagy and apoptosis in HCT-116 colon cancer cells. J. Ethnopharmacol. 2018, 211, 295–310. [Google Scholar] [CrossRef]
- Yao, M.; Li, F.; Xu, L.; Ma, L.; Zhang, S. 24-Dehydrocholesterol Reductase alleviates oxidative damage-induced apoptosis in alveolar epithelial cells via regulating Phosphatidylinositol-3-Kinase/Protein Kinase B activation. Bioengineered 2022, 13, 155–163. [Google Scholar] [CrossRef]
- Fang, R.; Houghton, P.J.; Hylands, P.J. Cytotoxic effects of compounds from Iris tectorum on human cancer cell lines. J. Ethnopharmacol. 2008, 118, 257–263. [Google Scholar] [CrossRef]
- Sun, J.; Wang, D.; Zhang, Y.; Mu, Q.; Li, M.; Hou, X.; Chen, J.; Yu, X.; Wang, S.; Yu, W. Cell Cycle Arrest and Autophagy Induced by Compound Kushen Injection in sw620 and sw480 Colorectal Cancer Cells with p53 Mutation. 2019. [Google Scholar]
- Lee, H.S.; Kim, E.N.; Jeong, G.S. Lupenone Protects Neuroblastoma SH-SY5y Cells Against Methamphetamine-Induced Apoptotic Cell Death via PI3K/Akt/mTOR Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boivin, D.; Blanchette, M.; Barrette, S.; Moghrabi, A.; Beliveau, R. Inhibition of cancer cell proliferation and suppression of TNF-induced activation of NFkappaB by edible berry juice. Anticancer Res. 2007, 27, 937–948. [Google Scholar] [PubMed]
- Tsujimoto, Y.; Shimizu, S. Another way to die: Autophagic programmed cell death. Cell Death Differ. 2005, 12 (Suppl. S2), 1528–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 and Autophagy. Methods Mol. Biol. 2008, 445, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Ren, D.; Wang, J.; Liu, X.; Zhang, H.; Wu, M.; Yang, G. Bruceine D induces lung cancer cell apoptosis and autophagy via the ROS/MAPK signaling pathway in vitro and in vivo. Cell Death Dis. 2020, 11, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Whiteman, M.W.; Lian, H.; Wang, G.; Singh, A.; Huang, D.; Denmark, T. A non-canonical MEK/ERK signaling pathway regulates autophagy via regulating Beclin 1. J. Biol. Chem. 2009, 284, 21412–21424. [Google Scholar] [CrossRef] [Green Version]
- Olsson, M.E.; Gustavsson, K.E.; Andersson, S.; Nilsson, A.; Duan, R.D. Inhibition of cancer cell proliferation in vitro by fruit and berry extracts and correlations with antioxidant levels. J. Agric. Food Chem. 2004, 52, 7264–7271. [Google Scholar] [CrossRef] [PubMed]










| Cell Line | SBO (mg/mL) | DTX 0.0 nM | DTX 0.1 nM | DTX 0.5 nM | DTX 1 nM |
|---|---|---|---|---|---|
| A549 | 0.0 | 2.1 ± 3.4 | 1.7 ± 2.9 | 6.0 ± 6.3 | |
| 0.5 | 10.9 ± 1.5 | 10.4 ± 1.6 (1.06) | 16.2 ± 1.0 (0.64) | 17.4 ± 1.2 (0.6) | |
| 1 | 19.8 ± 2.5 | 24.6 ± 3.2 (0.7) | 22.4 ± 4.6 (0.86) | 24.9 ± 2.8 (0.8) | |
| H23 | 0.0 | 3.0 ± 2.8 | 9.2 ± 1.6 | 14.0 ± 2.1 | |
| 0.5 | 22.8 ± 2.7 | 23.2 ± 4.3 (1.0) | 27.7 ± 1.4 (0.6) | 30.8 ± 3.6 (0.5) | |
| 1 | 26.5 ± 7.7 | 32.9 ± 4.0 (0.37) | 35.2 ± 2.9 (0.4) | 38.8 ± 1.1 (0.3) |
| No. | Compound Name | Compound Formula | Observed RT * (min) | Observed m/z | Mass Error (mDa) | References |
|---|---|---|---|---|---|---|
| 1. | Deoxycholic acid | C24H40O4 | 17.46 | 393.2979 | −5.2 | [46] |
| 2. | Lobeline alkaloid | C22H27NO2 | 14.42 | 338.2116 | 0.3 | [47,48] |
| 3. | Poricoic acid A | C32H48O5 | 6.57 | 513.3618 | 8.4 | [49] |
| 4. | 24-Dehydrocholesterol | C27H44O | 9.74 | 385.3494 | 7.7 | [50] |
| 5. | Blestrianol B | C37H32O7 | 18.72 | 589.222 | −0.2 | - |
| 6. | Iridobelamal A | C30H50O4 | 8.5 | 475.3809 | 5.8 | [51] |
| 7. | Esculentoside O | C35H54O10 | 6.37 | 635.3841 | 8.1 | - |
| 8. | Prosapogenin 1 | C47H76O18 | 18.03 | 707.3985 | −2.2 | - |
| 9. | Lupenone | C30H48O | 9.85 | 425.3797 | 4.5 | [52,53] |
| 10. | Campesterol-β-D-glucoside | C34H58O6 | 15.62 | 563.4281 | −4.4 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batbold, U.; Liu, J.-J. Chemosensitization Effect of Seabuckthorn (Hippophae rhamnoides L.) Pulp Oil via Autophagy and Senescence in NSCLC Cells. Foods 2022, 11, 1517. https://doi.org/10.3390/foods11101517
Batbold U, Liu J-J. Chemosensitization Effect of Seabuckthorn (Hippophae rhamnoides L.) Pulp Oil via Autophagy and Senescence in NSCLC Cells. Foods. 2022; 11(10):1517. https://doi.org/10.3390/foods11101517
Chicago/Turabian StyleBatbold, Uyanga, and Jun-Jen Liu. 2022. "Chemosensitization Effect of Seabuckthorn (Hippophae rhamnoides L.) Pulp Oil via Autophagy and Senescence in NSCLC Cells" Foods 11, no. 10: 1517. https://doi.org/10.3390/foods11101517