
Exopolysaccharides from the Energy Microalga Strain Botryococcus braunii: Purification, Characterization, and Antioxidant Activity

 ,
, 


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae and Culture Conditions
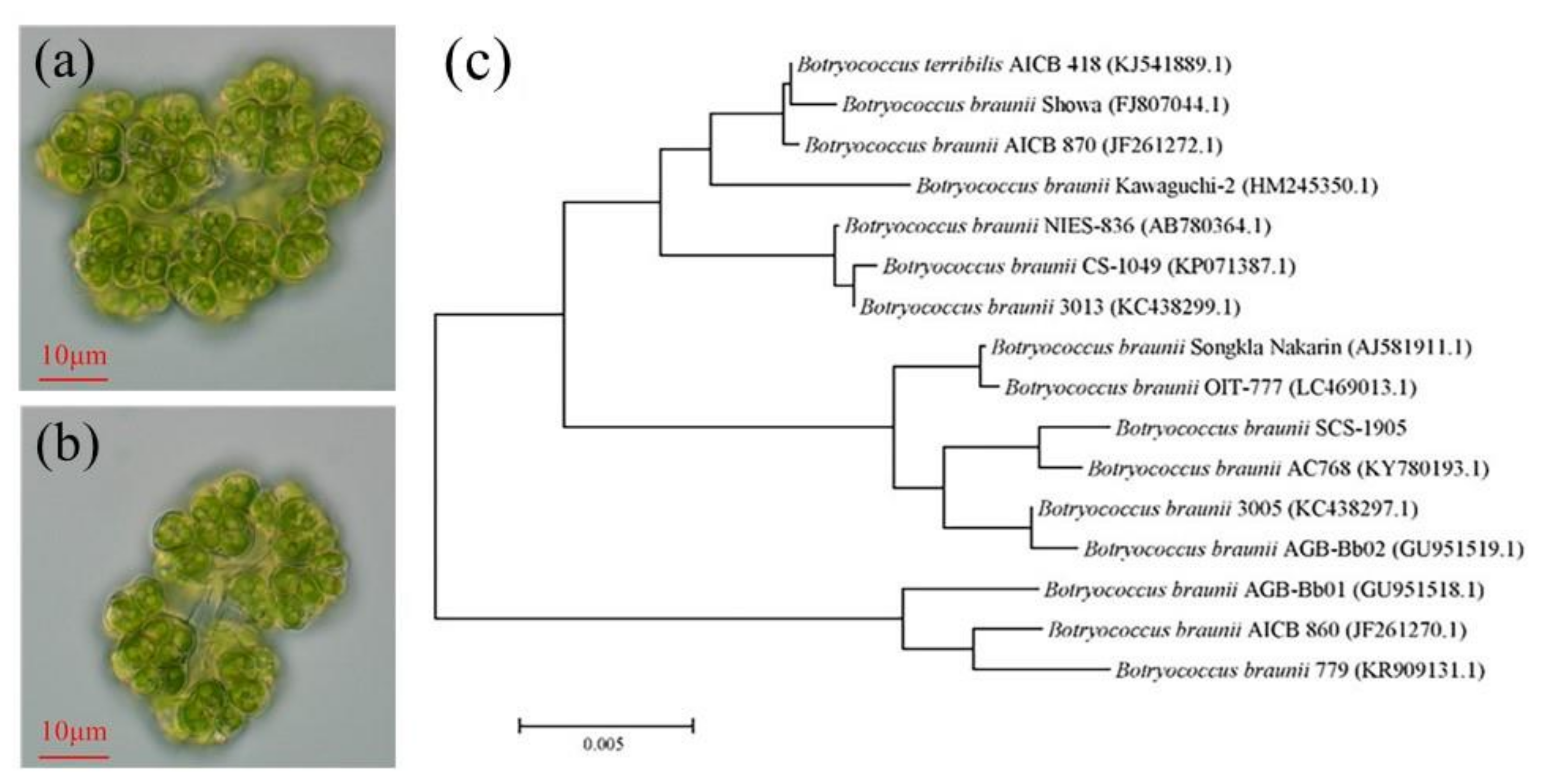
2.2. Morphological Observation and Phylogenetic Tree Construction
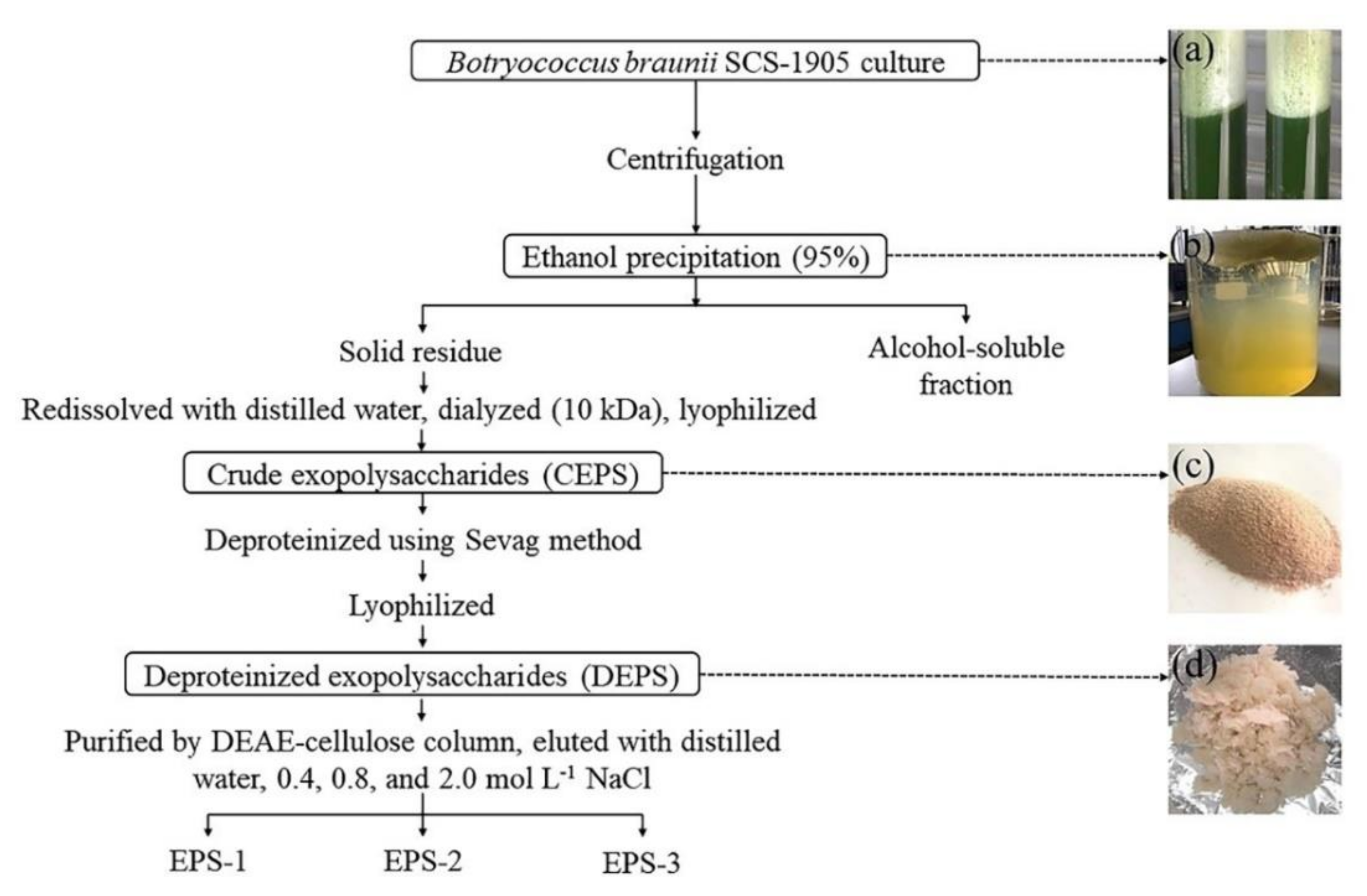
2.3. Preparation of Crude Exopolysaccharides (CEPS) and Deproteinized Exopolysaccharides (DEPS)
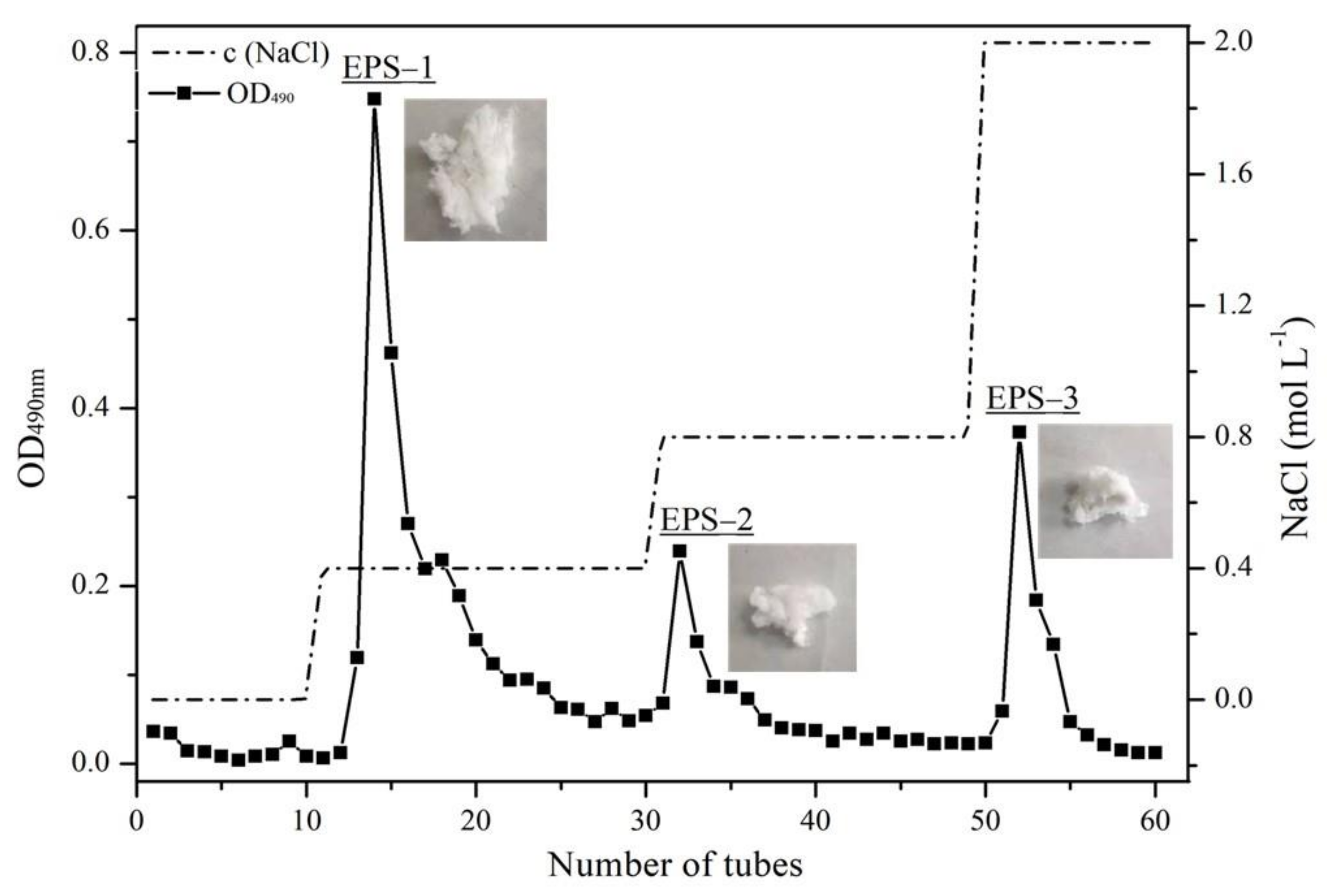
2.4. Purification by Ion Exchange Chromatography of Exopolysaccharides (EPS)
2.5. Determination of EPS Composition
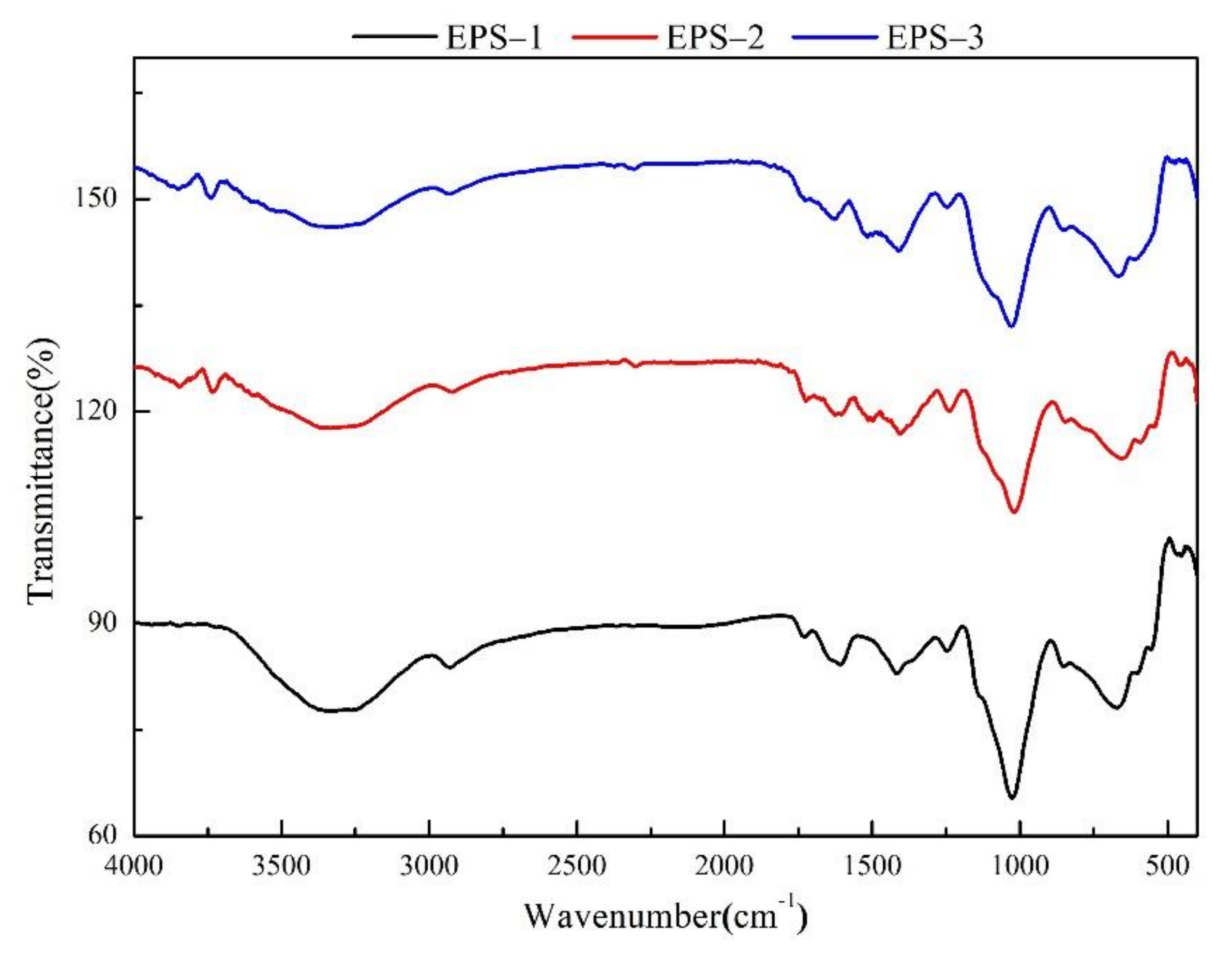
2.6. Fourier Transform Infrared Spectroscopy (FT-IR) Analysis
2.7. In Vitro Antioxidant Activity Assay
2.7.1. ABTS (2,2′-Azinobis (3-Ethylbenzothiazoline-6-sulfonic Acid)) Radical-Scavenging Ability
2.7.2. Hydroxyl Radical-Scavenging Ability
2.7.3. DPPH (2,2-Diphenyl-1-picrylhydrazyl) Radical-Scavenging Ability
2.7.4. Superoxide Anion Radical-Scavenging Ability
2.8. Statistical Analysis
3. Results
3.1. Morphological and Molecular Identification of Microalgal Strain
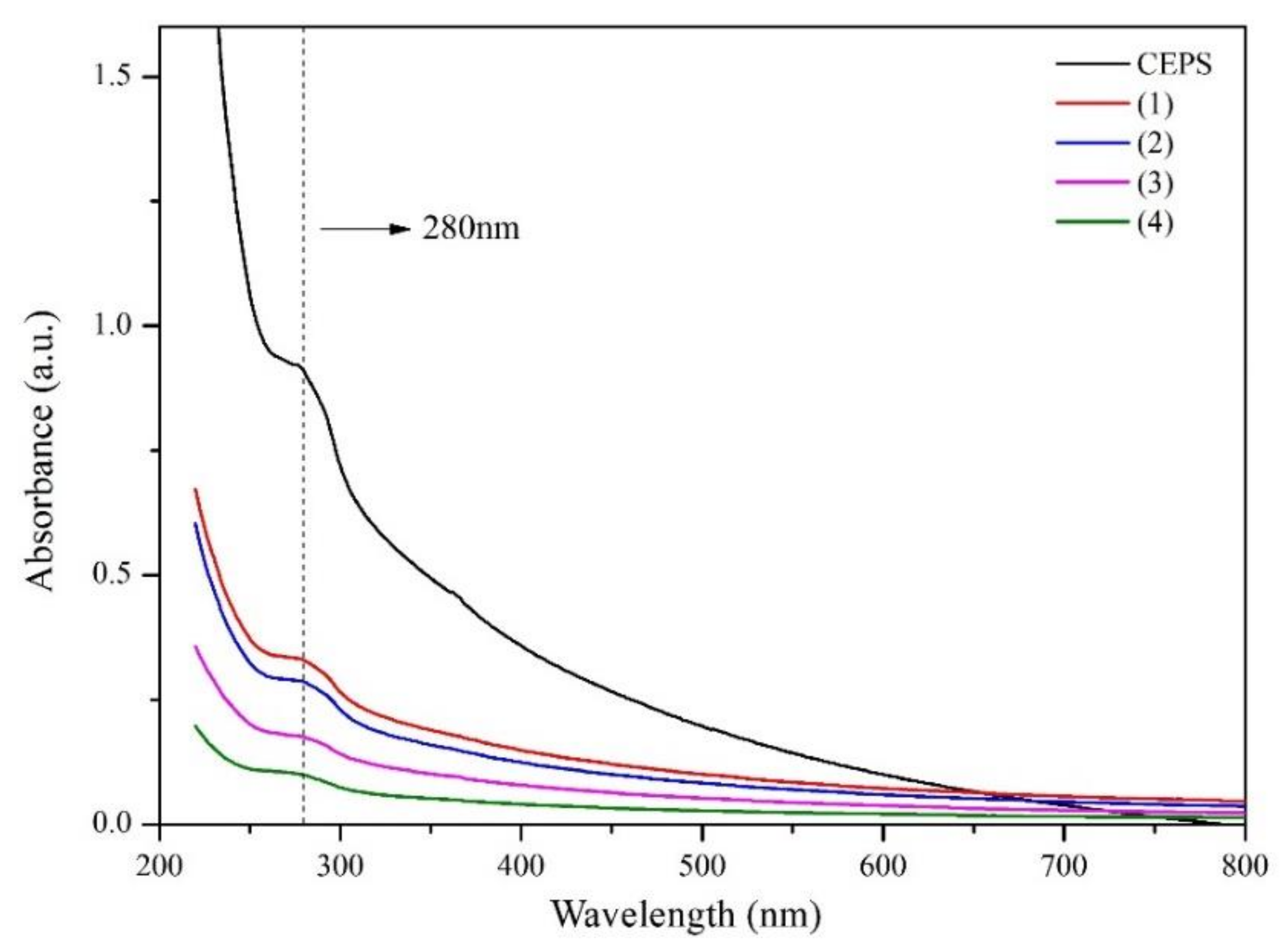
3.2. Preparation and Chemical Composition of EPS
3.3. Purification and Chemical Composition of Purified Fractions
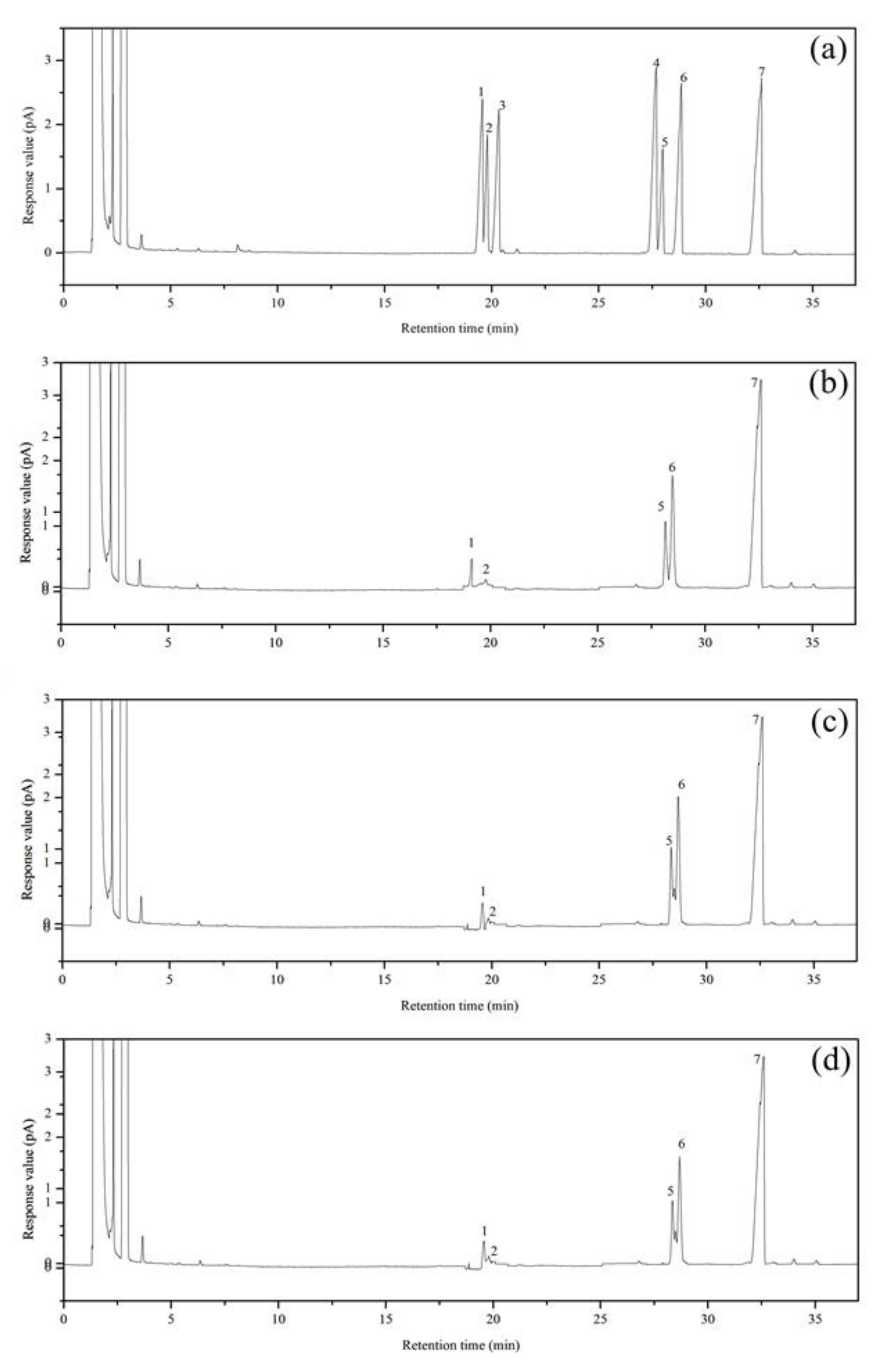
3.4. Primary Structures Characterization of Purified Fractions
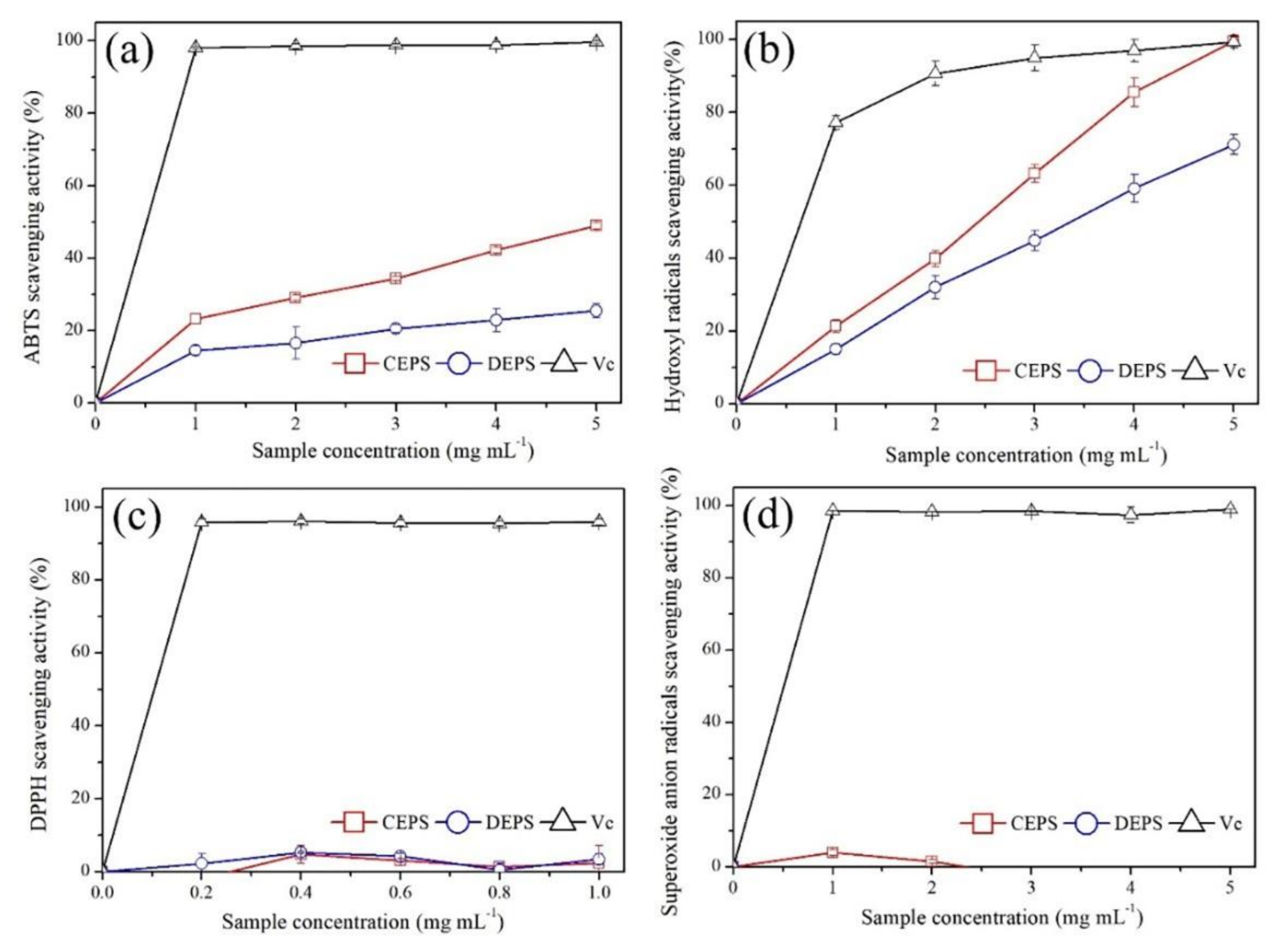
3.5. Evaluation of Antioxidant Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef] [PubMed]
- De Philippis, R.; Sili, C.; Paperi, R.; Vincenzini, M. Exopolysaccharide-producing cyanobacteria and their possible exploitation: A review. J. Appl. Phycol. 2001, 13, 293–299. [Google Scholar] [CrossRef]
- Mann, E.E.; Wozniak, D.J. Pseudomonas biofilm matrix composition and niche biology. FEMS Microbiol. Rev. 2012, 36, 893–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yalcin, I.; Hicsasmaz, Z.; Boz, B.; Bozoglu, F. Characterization of the extracellular polysaccharide from freshwater microalgae Chlorella sp. LWT-Food Sci. Technol. 1994, 27, 158–165. [Google Scholar] [CrossRef]
- Arad, S.; Rapoport, L.; Moshkovich, A.; van Moppes, D.; Karpasas, M.; Golan, R.; Golan, Y. Superior biolubricant from a species of red microalga. Langmuir 2006, 22, 7313–7317. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, A.M.; de Morais, R.M. Emergent sources of prebiotics: Seaweeds and microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef]
- Sun, L.; Wang, C.; Shi, Q.; Ma, C. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; de Morais, A.M.; de Morais, R.M. Influence of sulphate on the composition and antibacterial and antiviral properties of the exopolysaccharide from Porphyridium cruentum. Life Sci. 2014, 101, 56–63. [Google Scholar] [CrossRef]
- Guzman, S.; Gato, A.; Lamela, M.; Freire-Garabal, M.; Calleja, J. Anti-inflammatory and immunomodulatory activities of polysaccharide from Chlorella stigmatophora and Phaeodactylum tricornutum. Phytother. Res. 2003, 17, 665–670. [Google Scholar] [CrossRef]
- Zheng, W.; Chen, C.; Cheng, Q.; Wang, Y.; Chu, C. Oral administration of exopolysaccharide from Aphanothece halophytica (Chroococcales) significantly inhibits influenza virus (H1N1)-induced pneumonia in mice. Int. Immunopharmacol. 2006, 6, 1093–1099. [Google Scholar] [CrossRef]
- Senousy, H.H.; Beakes, G.W.; Hack, E. Phylogenetic placement of Botryococcus braunii (Trebouxiophyceae) and Botryococcus sudeticus isolate UTEX 2629 (Chlorophyceae). J. Appl. Phycol. 2004, 40, 412–423. [Google Scholar] [CrossRef]
- Metzger, P.; Berkaloff, C.; Casadevall, E.; Coute, A. Alkadiene-and botryococcene-producing races of wild strains of Botryococcus braunii. Phytochemistry 1985, 24, 2305–2312. [Google Scholar] [CrossRef]
- Metzger, P.; Allard, B.; Casadevall, E.; Berkaloff, C.; Couté, A. Structure and chemistry of a new chemical race of Botryococcus braunii (Chlorophyceae) that produces lycopadiene, a tetraterpenoid hydrocarbon. J. Appl. Phycol. 1990, 26, 258–266. [Google Scholar] [CrossRef]
- Kawachi, M.; Tanoi, T.; Demura, M.; Kaya, K.; Watanabe, M.M. Relationship between hydrocarbons and molecular phylogeny of Botryococcus braunii. Algal Res. 2012, 1, 114–119. [Google Scholar] [CrossRef]
- Casadevall, E.; Dif, D.; Largeau, C.; Gudin, C.; Chaumont, D.; Desanti, O. Studies on batch and continuous cultures of Botryococcus braunii: Hydrocarbon production in relation to physiological state, cell ultrastructure, and phosphate nutrition. Biotechnol. Bioeng. 1985, 27, 286–295. [Google Scholar] [CrossRef]
- Fernandes, H.; Tomé, M.; Lupi, F.; Fialho, A.; Sá-Correia, I.; Novais, J. Biosynthesis of high concentrations of an exopolysaccharide during the cultivation of the microalga Botryococcus braunii. Biotechnol. Lett. 1989, 11, 433–436. [Google Scholar] [CrossRef]
- Allard, B.; Casadevall, E. Carbohydrate composition and characterization of sugars from the green microalga Botryococcus braunii. Phytochemistry 1990, 29, 1875–1878. [Google Scholar] [CrossRef]
- Li, T.; Xu, J.; Gao, B.; Xiang, W.; Li, A.; Zhang, C. Morphology, growth, biochemical composition and photosynthetic performance of Chlorella vulgaris (Trebouxiophyceae) under low and high nitrogen supplies. Algal Res. 2016, 16, 481–491. [Google Scholar] [CrossRef]
- Chen, Z.; Lei, X.; Zhang, B.; Yang, L.; Zhang, H.; Zhang, J.; Li, Y.; Zheng, W.; Tian, Y.; Liu, J. First report of Pseudobodo sp., a new pathogen for a potential energy-producing algae: Chlorella vulgaris cultures. PLoS ONE 2014, 9, e89571. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Chem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Reim, R.E. Total sulfite in cellulosics by ion exclusion chromatography with electrochemical detection. J. Food. Sci. 1991, 56, 1079–1090. [Google Scholar] [CrossRef]
- Luo, J.; Liu, J.; Sun, Y.; Ye, H.; Zhou, C.; Zeng, X. Medium optimization, preliminary characterization and antioxidant activity in vivo of mycelial polysaccharide from Phellinus baumii Pilát. Carbohydr. Polym. 2010, 81, 533–540. [Google Scholar] [CrossRef]
- Li, X.; Lin, J.; Gao, Y.; Han, W.; Chen, D. Antioxidant activity and mechanism of Rhizoma Cimicifugae. Chem. Cent. J. 2012, 6, 140. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jiang, B.; Zhang, T.; Mu, W.; Liu, J. Antioxidant and free radical-scavenging activities of chickpea protein hydrolysate (CPH). Food Chem. 2008, 106, 444–450. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, X.; Wu, L.; Tong, A.; Zhao, L.; Liu, B.; Zhao, C. Physicochemical characterization of polysaccharides from Chlorella pyrenoidosa and its anti-ageing effects in Drosophila melanogaster. Carbohydr. Polym. 2018, 185, 120–126. [Google Scholar] [CrossRef]
- Wu, M.; Feng, H.; Song, J.; Chen, L.; Xu, Z.; Xia, W.; Zhang, W. Structural elucidation and immunomodulatory activity of a neutral polysaccharide from the Kushui Rose (Rosa setate x Rosa rugosa) waste. Carbohydr. Polym. 2020, 232, 115804. [Google Scholar] [CrossRef]
- Wang, B. Chemical characterization and ameliorating effect of polysaccharide from Chinese jujube on intestine oxidative injury by ischemia and reperfusion. Int. J. Biol. Macromol. 2011, 48, 386–391. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; Nikšić, M.; Vrvić, M.M.; Todorović, N.; Jakovljević, D.; Van Griensven, L.J. Antioxidative activities and chemical characterization of polysaccharide extracts from the widely used mushrooms Ganoderma applanatum, Ganoderma lucidum, Lentinus edodes and Trametes versicolor. J. Food Compost. Anal. 2012, 26, 144–153. [Google Scholar] [CrossRef]
- Kacurakova, M.; Capek, P.; Sasinkova, V.; Wellner, N.; Ebringerova, A. FT-IR study of plant cell wall model compounds: Pectic polysaccharides and hemicelluloses. Carbohydr. Polym. 2000, 43, 195–203. [Google Scholar] [CrossRef]
- Liu, C.H.; Wang, C.H.; Xu, Z.L.; Wang, Y. Isolation, chemical characterization and antioxidant activities of two polysaccharides from the gel and the skin of Aloe barbadensis Miller irrigated with sea water. Process Biochem. 2007, 42, 961–970. [Google Scholar]
- Gouveia, J.D.; Ruiz, J.; van den Broek, L.A.; Hesselink, T.; Peters, S.; Kleinegris, D.M.; Smith, A.G.; van der Veen, D.; Barbosa, M.J.; Wijffels, R.H. Botryococcus braunii strains compared for biomass productivity, hydrocarbon and carbohydrate content. J. Biotechnol. 2017, 248, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Metzger, P.; Largeau, C. Botryococcus braunii: A rich source for hydrocarbons and related ether lipids. Appl. Microbiol. Biotechnol. 2005, 66, 486–496. [Google Scholar] [CrossRef]
- Lupi, F.M.; Fernandes, H.M.; Sá-Correia, I.; Novais, J.M. Temperature profiles of cellular growth and exopolysaccharide synthesis by Botryococus braunii Kütz. UC 58. J. Appl. Phycol. 1991, 3, 35–42. [Google Scholar] [CrossRef]
- Chiovitti, A.; Harper, R.E.; Willis, A.; Bacic, A.; Mulvaney, P.; Wetherbee, R. Variations in the substituted 3-linked mannans closely associated with the silicified walls of diatoms. J. Appl. Phycol. 2005, 41, 1154–1161. [Google Scholar] [CrossRef]
- Wang, W.N.; Li, Y.; Zhang, Y.; Xiang, W.Z.; Li, A.F.; Li, T. Comparison on characterization and antioxidant activity of exopolysaccharides from two Porphyridium strains. Appl. Phycol. 2021, 33, 2983–2994. [Google Scholar] [CrossRef]
- Heiss, C.; Black, I.; Ishihara, M.; Tatli, M.; Devarenne, T.P.; Azadi, P. Structure of the polysaccharide sheath from the B race of the green microalga Botryococcus braunii. Algal Res. 2021, 55, 102252. [Google Scholar] [CrossRef]
- Weiss, T.L.; Roth, R.; Goodson, C.; Vitha, S.; Black, I.; Azadi, P.; Rusch, J.; Holzenburg, A.; Devarenne, T.P.; Goodenough, U. Colony organization in the green alga Botryococcus braunii (Race B) is specified by a complex extracellular matrix. Eukaryot. Cell 2012, 11, 1424–1440. [Google Scholar] [CrossRef] [Green Version]
- Tatli, M.; Ishihara, M.; Heiss, C.; Browne, D.R.; Dangott, L.J.; Vitha, S.; Azadi, P.; Devarenne, T.P. Polysaccharide associated protein (PSAP) from the green microalga Botryococcus braunii is a unique extracellular matrix hydroxyproline-rich glycoprotein. Algal Res. 2018, 29, 92–103. [Google Scholar] [CrossRef]
- Banerjee, A.; Sharma, R.; Chisti, Y.; Banerjee, U. Botryococcus braunii: A renewable source of hydrocarbons and other chemicals. Crit. Rev. Biotechnol. 2002, 22, 245–279. [Google Scholar] [CrossRef]
- Kaplan, D.; Christiaen, D.; Arad, S.M. Chelating properties of extracellular polysaccharides from Chlorella spp. Appl. Environ. Microbiol. 1987, 53, 2953–2956. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.S.; Hong, M.E.; Jung, S.C.; Ha, S.J.; Yu, B.J.; Koo, H.M.; Park, S.M.; Seo, J.H.; Kweon, D.H.; Park, J.C. Improved galactose fermentation of Saccharomyces cerevisiae through inverse metabolic engineering. Biotechnol. Bioeng. 2011, 108, 621–631. [Google Scholar] [CrossRef]
- García-Cubero, R.; Cabanelas, I.T.; Sijtsma, L.; Kleinegris, D.M.; Barbosa, M.J. Production of exopolysaccharide by Botryococcus braunii CCALA 778 under laboratory simulated Mediterranean climate conditions. Algal Res. 2018, 29, 330–336. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, L.; Chen, F. Production and characterization of exopolysaccharides from Chlorella zofingiensis and Chlorella vulgaris with anti-colorectal cancer activity. Int. J. Biol. Macromol. 2019, 134, 976–983. [Google Scholar] [CrossRef]
- Fritzsche, S.; Blenk, P.; Christian, J.; Castiglione, K.; Becker, A.M. Inhibitory properties of crude microalgal extracts on the in vitro replication of cyprinid herpesvirus 3. Sci. Rep. 2021, 11, 23134. [Google Scholar] [CrossRef]
- Guzman-Murillo, M.; Ascencio, F. Anti-adhesive activity of sulphated exopolysaccharides of microalgae on attachment of red sore disease-associated bacteria and Helicobacter pylori to tissue culture cells. Lett. Appl. Microbiol. 2000, 30, 473–478. [Google Scholar] [CrossRef]
- Fujitani, N.; Sakaki, S.; Yamaguchi, Y.; Takenaka, H. Inhibitory effects of microalgae on the activation of hyaluronidase. J. Appl. Phycol. 2001, 13, 489–492. [Google Scholar] [CrossRef]
- Jin, J.-O.; Chauhan, P.S.; Arukha, A.P.; Chavda, V.; Dubey, A.; Yadav, D. The therapeutic potential of the anticancer activity of fucoidan: Current advances and hurdles. Mar. Drugs 2021, 19, 265. [Google Scholar] [CrossRef]
- Fernandez, O.; Béthencourt, L.; Quero, A.; Sangwan, R.S.; Clément, C. Trehalose and plant stress responses: Friend or foe? Trends Plant Sci. 2010, 15, 409–417. [Google Scholar] [CrossRef]
- Shen, S.; Jia, S.; Wu, Y.; Yan, R.; Lin, Y.; Zhao, D.; Han, P. Effect of culture conditions on the physicochemical properties and antioxidant activities of polysaccharides from Nostoc flagelliforme. Carbohydr. Polym. 2018, 198, 426–433. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compost. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Dudonne, S.; Vitrac, X.; Coutiere, P.; Woillez, M.; Mérillon, J.M. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD, and ORAC assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, B.; Ibrahim, S.; Gao, S.S.; Yang, H.; Huang, W. Purification, characterization and antioxidant activity of polysaccharides from Flammulina velutipes residue. Carbohydr. Polym. 2016, 145, 71–77. [Google Scholar] [CrossRef]
- Bayona, K.C.; Garcés, L.A. Effect of different media on exopolysaccharide and biomass production by the green microalga Botryococcus braunii. J. Appl. Phycol. 2014, 26, 2087–2095. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, P.; Li, Z.; Zhang, H.; Xu, Z.; Li, P. Antioxidant activities of sulfated polysaccharide fractions from Porphyra haitanesis. J. Appl. Phycol. 2003, 15, 305–310. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Composition (%DW) | Sample | |
|---|---|---|
| CEPS | DEPS | |
| Total carbohydrate | 42.39 ± 0.15 | 58.88 ± 1.23 |
| Protein | 4.04 ± 0.14 | 2.30 ± 0.08 |
| Uronic acid | 7.43 ± 0.47 | 8.83 ± 0.99 |
| Sulfate | 1.95 ± 0.01 | 1.52 ± 0.04 |
| Chemical Composition (%DW) | Sample | ||
|---|---|---|---|
| EPS-1 | EPS-2 | EPS-3 | |
| Total carbohydrate | 53.27 ± 0.09 a1 | 42.36 ± 0.18 b1 | 39.15 ± 0.17 c1 |
| Protein | 0.95 ± 0.01 a2 | 0.80 ± 0.02 c2 | 0.86 ± 0.01 b2 |
| Uronic acid | 7.04 ± 0.50 a3 | 6.82 ± 0.42 a3 | 6.56 ± 0.39 a3 |
| Percentage (%Total Carbohydrate) | Sample | ||
|---|---|---|---|
| EPS-1 | EPS-2 | EPS-3 | |
| Galactose | 54.12 ± 0.19 a1 | 53.82 ± 0.80 a1 | 52.34 ± 0.61 b1 |
| Glucose | 34.60 ± 0.02 | 34.65 ± 0.86 | 35.53 ± 0.20 |
| Arabinose | 9.41 ± 0.09 b2 | 9.54 ± 0.24 b2 | 10.32 ± 0.16 a2 |
| Fucose | 1.87 ± 0.25 | 1.99 ± 0.18 | 1.80 ± 0.25 |
| IC50 (mg·mL−1) | Sample | ||
|---|---|---|---|
| CEPS | DEPS | Vc | |
| ABTS | 5.13 ± 0.11 b1 | 12.38 ± 1.06 a1 | 0.003 ± 0.001 c1 |
| OH· | 1.67 ± 0.06 b2 | 3.04 ± 0.30 a2 | 0.240 ± 0.001 c2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.-N.; Li, T.; Li, Y.; Zhang, Y.; Wu, H.-L.; Xiang, W.-Z.; Li, A.-F. Exopolysaccharides from the Energy Microalga Strain Botryococcus braunii: Purification, Characterization, and Antioxidant Activity. Foods 2022, 11, 110. https://doi.org/10.3390/foods11010110
Wang W-N, Li T, Li Y, Zhang Y, Wu H-L, Xiang W-Z, Li A-F. Exopolysaccharides from the Energy Microalga Strain Botryococcus braunii: Purification, Characterization, and Antioxidant Activity. Foods. 2022; 11(1):110. https://doi.org/10.3390/foods11010110
Chicago/Turabian StyleWang, Wei-Nan, Tao Li, Yi Li, Ying Zhang, Hua-Lian Wu, Wen-Zhou Xiang, and Ai-Fen Li. 2022. "Exopolysaccharides from the Energy Microalga Strain Botryococcus braunii: Purification, Characterization, and Antioxidant Activity" Foods 11, no. 1: 110. https://doi.org/10.3390/foods11010110