
Food Metabolites as Tools for Authentication, Processing, and Nutritive Value Assessment

 , ,
, ,  ,
,  ,
,  and
and
Abstract
:1. Metabolomic Studies
1.1. Which “omic” Is It?
1.2. Applications of Metabolomics
2. Metabolomic Studies in Foods
2.1. General Considerations
2.2. Primary and Secondary Metabolites
3. Platforms for Metabolomic Studies: Analytical Methods and Data Processing
3.1. General Aspects of Metabolomic Workflow
3.2. Separation Techniques
3.2.1. Gas Chromatography (GC)
3.2.2. Liquid Chromatography (LC)
3.2.3. Capillary Electrophoresis (CE)
3.2.4. Ion Mobility–Mass Spectrometry (IM–MS)
3.3. Detection Techniques
3.3.1. Nuclear Magnetic Resonance (NMR)
3.3.2. Mass Spectrometry (MS)
3.4. Data Processing
4. Application in Foods
4.1. Food Safety
4.1.1. Quality Control for Foods
4.1.2. Authentication for Foods
4.1.3. Food Toxins
4.2. Nutritional Value Assessment of Foods
4.3. Food Processing
5. Main Challenges and Difficulties
6. Trends and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Coulier, L.; Wopereis, S.; Rubingh, C.; Hendriks, H.; Radonjic´, M.; Jellema, R.H. Systems Biology. In An Introduction to Key Concepts in Medicinal Chemistry; Elsevier B.V.: Amsterdam, The Netherlands, 2009; pp. 279–312. [Google Scholar]
- Benson, D.A.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Rapp, B.A.; Wheeler, D.L. GenBank. Nucleic Acids Res. 2002, 30, 17–20. [Google Scholar] [CrossRef]
- Fanos, V.; Antonucci, R.; Barberini, L.; Atzori, L. Urinary Metabolomics in Newborns and Infants. Adv. Clin. Chem. 2012, 58, 193–223. [Google Scholar] [CrossRef]
- Park, S.T.; Kim, J. Trends in next-generation sequencing and a new era for whole genome sequencing. Int. Neurourol. J. 2016, 20, 76–83. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Ji, L.; Chen, J. Metabolomic Profiling of Neoplastic Lesions in Mice. In Cell-Wide Metabolic Alterations Associated with Malignancy, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; Volume 543. [Google Scholar] [CrossRef]
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. Mass spectrometry-based metabolomics. Mass Spectrom. Rev. 2006, 26, 51–78. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Yang, Z. Single cell metabolomics using mass spectrometry: Techniques and data analysis. Anal. Chim. Acta 2021, 1143, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, G.R.; Sisodia, B.S.; Shikha. Secondary Metabolites: Metabolomics for Secondary Metabolites. In New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Secondary Metabolites Biochesmistry and Applications; Gupta, V.K., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 333–344. [Google Scholar]
- Hall, R.D. Plant metabolomics: From holistic hope, to hype, to hot topic. New Phytol. 2006, 169, 453–468. [Google Scholar] [CrossRef]
- De Castro, M.D.L.; Priego-Capote, F. The analytical process to search for metabolomics biomarkers. J. Pharm. Biomed. Anal. 2018, 147, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Howlett, K.; Patterson, K.; Wang, B. Introduction of a new method for two-dimensional NMR quantitative analysis in metabolomics studies. Anal. Biochem. 2020, 597, 113692. [Google Scholar] [CrossRef]
- Rocchetti, G.; O’Callaghan, T.F. Application of metabolomics to assess milk quality and traceability. Curr. Opin. Food Sci. 2021, 40, 168–178. [Google Scholar] [CrossRef]
- Castro-Puyana, M.; Herrero, M. Metabolomics approaches based on mass spectrometry for food safety, quality and traceability. Trends Anal. Chem. 2013, 52, 74–87. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, C. Metabolomics: Bridging Chemistry and Biology in Drug Discovery and Development. Curr. Pharmacol. Rep. 2017, 3, 16–25. [Google Scholar] [CrossRef]
- Campillo, J.A.; Sevilla, A.; González-Fernández, C.; Bellas, J.; Bernal, C.; Cánovas, M.; Albentosa, M. Metabolomic responses of mussel Mytilus galloprovincialis to fluoranthene exposure under different nutritive conditions. Mar. Environ. Res. 2019, 144, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, W.X. NMR-based metabolomic studies on the toxicological effects of cadmium and copper on green mussels Perna viridis. Aquat. Toxicol. 2010, 100, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, X.; You, L.; Zhou, D.; Wang, Q.; Li, F.; Cong, M.; Li, L.; Zhao, J.; Liu, D.; et al. Benzo(a)pyrene-induced metabolic responses in Manila clam Ruditapes philippinarum by proton nuclear magnetic resonance (1H NMR) based metabolomics. Environ. Toxicol. Pharmacol. 2011, 32, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Murithi, J.M.; Owen, E.S.; Istvan, E.S.; Lee, M.C.S.; Ottilie, S.; Chibale, K.; Goldberg, D.E.; Winzeler, E.A.; Llinás, M.; Fidock, D.A.; et al. Combining Stage Specificity and Metabolomic Profiling to Advance Antimalarial Drug Discovery. Cell Chem. Biol. 2020, 27, 158–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klont, F.; Kremer, D.; Gomes Neto, A.W.; Berger, S.P.; Touw, D.J.; Hak, E.; Bonner, R.; Bakker, S.J.L.; Hopfgartner, G. Metabolomics data complemented drug use information in epidemiological databases: Pilot study of potential kidney donors. J. Clin. Epidemiol. 2021, 135, 10–16. [Google Scholar] [CrossRef]
- Cramer, G.R.; Urano, K.; Delrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Almuhayawi, M.S.; Hassan, A.H.A.; Al, S.K. Influence of elevated CO 2 on nutritive value and health-promoting prospective of three genotypes of Alfalfa sprouts (Medicago Sativa). Food Chem. 2021, 340, 128147. [Google Scholar] [CrossRef]
- Cozzolino, D. Foodomics and infrared spectroscopy: From compounds to functionality. Curr. Opin. Food Sci. 2015, 4, 39–43. [Google Scholar] [CrossRef]
- Jacobs, D.M.; van den Berg, M.A.; Hall, R.D. Towards superior plant-based foods using metabolomics. Curr. Opin. Biotechnol. 2021, 70, 23–28. [Google Scholar] [CrossRef]
- Kim, S.; Kim, J.; Yun, E.J.; Kim, K.H. Food metabolomics: From farm to human. Curr. Opin. Biotechnol. 2016, 37, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Pavagadhi, S.; Swarup, S. Metabolomics for evaluating flavor-associated metabolites in plant-based products. Metabolites 2020, 10, 197. [Google Scholar] [CrossRef]
- Wu, B.; Wei, F.; Xu, S.; Xie, Y.; Lv, X.; Chen, H.; Huang, F. Mass spectrometry-based lipidomics as a powerful platform in foodomics research. Trends Food Sci. Technol. 2021, 107, 358–376. [Google Scholar] [CrossRef]
- Cifuentes, A. Food analysis and foodomics. J. Chromatogr. A 2009, 1216, 7109. [Google Scholar] [CrossRef] [Green Version]
- Skov, T.; Honoré, A.H.; Jensen, H.M.; Næs, T.; Engelsen, S.B. Chemometrics in foodomics: Handling data structures from multiple analytical platforms. Trends Anal. Chem. 2014, 60, 71–79. [Google Scholar] [CrossRef]
- Steuer, A.E.; Brockbals, L.; Kraemer, T. Metabolomic strategies in biomarker research-new approach for indirect identification of drug consumption and sample manipulation in clinical and forensic toxicology? Front. Chem. 2019, 7, 319. [Google Scholar] [CrossRef]
- Sanchez, S.; Demain, A.L. Metabolic regulation and overproduction of primary metabolites. Microb. Biotechnol. 2008, 1, 283–319. [Google Scholar] [CrossRef]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root exudation of primary metabolites: Mechanisms and their roles in plant responses to environmental stimuli. Front. Plant Sci. 2019, 10, 157. [Google Scholar] [CrossRef] [Green Version]
- Campos, M.R.S. (Ed.) Bioactive Compounds: Health Benefits and Potential Applications; Woodhead Publishing: Sawtson, UK, 2019. [Google Scholar]
- Aharoni, A.; Galili, G. Metabolic engineering of the plant primary-secondary metabolism interface. Curr. Opin. Biotechnol. 2011, 22, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Tyc, O.; Song, C.; Dickschat, J.S.; Vos, M.; Garbeva, P. The Ecological Role of Volatile and Soluble Secondary Metabolites Produced by Soil Bacteria. Trends Microbiol. 2017, 25, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Bills, G.F.; Gloer, J.B. Biologically active secondary metabolites from the fungi. In The Fungal Kingdom; Wiley: Hoboken, NJ, USA, 2017; pp. 1087–1119. [Google Scholar] [CrossRef]
- Rastegari, A.A.; Yadav, A.N.; Yadav, N. Genetic Manipulation of Secondary Metabolites Producers. In New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Secondary Metabolites Biochesmistry and Applications; Gupta, V.K., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 13–29. [Google Scholar] [CrossRef]
- Gokulan, K.; Khare, S.; Cerniglia, C. Production of Secondary Metabolites of Bacteria. In Encyclopedia of Food Microbiology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 561–569. [Google Scholar]
- Kumar, A.; Naraian, R. Producers of Bioactive Compounds. In New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Secondary Metabolites Biochesmistry and Applications; Gupta, V.K., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 205–221. [Google Scholar]
- Toffano, R.B.D.; Hillesheim, E.; Mathias, M.G.; Coelho-Landell, C.A.; Salomão, R.G.; Almada, M.O.R.V.; Camarneiro, J.M.; Barros, T.T.; Camelo-Junior, J.S.; Rezzi, S.; et al. Validation of the brazilian healthy eating index-revised using biomarkers in children and adolescents. Nutrients 2018, 10, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Sanchez, P.; De Vos, R.C.H.; Jonker, H.H.; Mumm, R.; Hall, R.D.; Bialek, L.; Leenman, R.; Strassburg, K.; Vreeken, R.; Hankemeier, T.; et al. Comprehensive metabolomics to evaluate the impact of industrial processing on the phytochemical composition of vegetable purees. Food Chem. 2015, 168, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Capuano, E.; Boerrigter-Eenling, R.; van der Veer, G.; van Ruth, S.M. Analytical authentication of organic products: An overview of markers. J. Sci. Food Agric. 2013, 93, 12–28. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, X.; Han, L.; Jia, L.; Liu, E.; Li, Z.; Yu, H.; Wang, Y.; Gao, X.; Yang, W. Integration of multicomponent characterization, untargeted metabolomics and mass spectrometry imaging to unveil the holistic chemical transformations and key markers associated with wine steaming of Ligustri Lucidi Fructus. J. Chromatogr. A 2020, 1624, 461228. [Google Scholar] [CrossRef]
- Amargianitaki, M.; Spyros, A. NMR-based metabolomics in wine quality control and authentication. Chem. Biol. Technol. Agric. 2017, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Giacometti, J.; Tomljanović, A.B.; Josić, D. Application of proteomics and metabolomics for investigation of food toxins. Food Res. Int. 2013, 54, 1042–1051. [Google Scholar] [CrossRef]
- Oyedeji, A.B.; Green, E.; Adebiyi, J.A.; Ogundele, O.M.; Gbashi, S.; Adefisoye, M.A.; Oyeyinka, S.A.; Adebo, O.A. Metabolomic Approaches for the Determination of Metabolites from Pathogenic Microorganisms: A Review. Food Res. Int. 2021, 140, 110042. [Google Scholar] [CrossRef]
- Rawat, J.M.; Bhandari, A.; Raturi, M.; Rawat, B. Agrobacterium rhizogenes Mediated Hairy Root Cultures: A Promising Approach for Production of Useful Metabolites. In New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Secondary Metabolites Biochesmistry and Applications; Gupta, V.K., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 103–118. [Google Scholar]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Zhong, J.-J.; Xiao, J.-H. Secondary Metabolites from Higher Fungi: Discovery, Bioactivity, and Bioproduction. In Biotechnology in China I. From Bioreaction to Bioseparation and Bioremediation; Zhong, J.-J., Bai, F.-W., Zhang, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 79–150. [Google Scholar]
- Andryukov, B.; Mikhailov, V.; Besednova, N. The biotechnological potential of secondary metabolites from marine bacteria. J. Mar. Sci. Eng. 2019, 7, 176. [Google Scholar] [CrossRef] [Green Version]
- Mohan, C.D.; Rangappa, S.; Nayak, S.C.; Jadimurthy, R.; Wang, L.; Sethi, G.; Garg, M.; Rangappa, K.S. Bacteria as a treasure house of secondary metabolites with anticancer potential. Semin. Cancer Biol. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Hamacher, M.; Malisch, C.S.; Reinsch, T.; Taube, F.; Loges, R. Evaluation of yield formation and nutritive value of forage legumes and herbs with potential for diverse grasslands due to their concentration in plant specialized metabolites. Eur. J. Agron. 2021, 128, 126307. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, J.; Teng, Z.; Luo, Y.; Yu, L.; Simko, I.; Chen, P. Identification of marker compounds for predicting browning of fresh-cut lettuce using untargeted UHPLC-HRMS metabolomics. Postharvest Biol. Technol. 2021, 180, 111626. [Google Scholar] [CrossRef]
- Pereira, G.E.; Padhi, E.M.T.; Sudarshana, M.R.; Fialho, F.B.; Medina-Plaza, C.; Girardello, R.C.; Tseng, D.; Bruce, R.C.; Erdmann, J.N.; Slupsky, C.M.; et al. Impact of grapevine red blotch disease on primary and secondary metabolites in ‘Cabernet Sauvignon’ grape tissues. Food Chem. 2021, 342, 128312. [Google Scholar] [CrossRef]
- Zhan, J.; Yu, X.J.; Zhong, Y.Y.; Zhang, Z.T.; Cui, X.M.; Peng, J.F.; Feng, R.; Liu, X.T.; Zhu, Y. Generic and rapid determination of veterinary drug residues and other contaminants in raw milk by ultra performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 906, 48–57. [Google Scholar] [CrossRef]
- Hanson, J.R. Natural Products: The Secondary Metabolites. In Tutorial Chemistry Texts; Royal Society of Chemistry: London, UK, 2003. [Google Scholar]
- Bajkacz, S.; Kycia-Słocka, E. Liquid chromatography in food analysis. In Chemical Analysis of Food—Techniques and Applications, 2nd ed.; Pico, Y., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 391–455. [Google Scholar]
- Cubero-Leon, E.; Peñalver, R.; Maquet, A. Review on metabolomics for food authentication. Food Res. Int. 2014, 60, 95–107. [Google Scholar] [CrossRef]
- Choi, M.Y.; Choi, W.; Park, J.H.; Lim, J.; Kwon, S.W. Determination of coffee origins by integrated metabolomic approach of combining multiple analytical data. Food Chem. 2010, 121, 1260–1268. [Google Scholar] [CrossRef]
- Cevallos-cevallos, J.M.; Etxeberria, E.; Danyluk, M.D.; Rodrick, G.E. Metabolomic analysis in food science: A review. Trends Food Sci. Technol. 2009, 20, 557–566. [Google Scholar] [CrossRef]
- Oms-Oliu, G.; Odriozola-Serrano, I.; Martín-Belloso, O. Metabolomics for assessing safety and quality of plant-derived food. Food Res. Int. J. 2013, 54, 1172–1183. [Google Scholar] [CrossRef]
- Mamat, S.F.; Azizan, K.A.; Baharum, S.N.; Noor, N.M.; Aizat, W.M. GC-MS and LC-MS analyses reveal the distribution of primary and secondary metabolites in mangosteen (Garcinia mangostana Linn.) fruit during ripening. Sci. Hortic. 2020, 262, 109004. [Google Scholar] [CrossRef]
- Sørensen, K.M.; Khakimov, B.; Engelsen, S.B. The use of rapid spectroscopic screening methods to detect adulteration of food raw materials and ingredients. Curr. Opin. Food Sci. 2016, 10, 45–51. [Google Scholar] [CrossRef]
- Mairinger, T.; Causon, T.J.; Hann, S. The potential of ion mobility–mass spectrometry for non-targeted metabolomics. Curr. Opin. Chem. Biol. 2018, 42, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Lebanov, L.; Ghiasvand, A.; Paull, B. Data handling and data analysis in metabolomic studies of essential oils using GC-MS. J. Chromatogr. A 2021, 1640, 461896. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hu, Q.; Yu, Q.; Chen, Y.; Zhao, Y.; Qie, M. Metabolomics Analysis in Food Authentication. In Comprehensive Foodomics; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar] [CrossRef]
- Vu Dang, H.; Marini, F. Editorial: Chemometrics-based spectroscopy for pharmaceutical and biomedical analysis. Front. Chem. 2019, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Oliveri, P.; Malegori, C.; Casale, M. Chemometrics: Multivariate analysis of chemical data. In Chemical Analysis of Food—Techniques and Applications, 2nd ed.; Pico, Y., Ed.; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Sparkman, O.D.; Penton, Z.E.; Kitson, F.G. Gas Chromatography and Mass Spectrometry: A Practical Guide, 2nd ed.; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- McNair, H.M.; Miller, J.M.; Snow, N.H. Basic Gas Chromatography, 3rd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019. [Google Scholar]
- Engewald, W.; Dettmer-Wilde, K. Practical Gas Chromatography: A Comprehensive Reference; Dettmer-Wilde, K., Engewald, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; p. 902. [Google Scholar]
- Jennings, W.; Mittlefehldt, E.; Stremple, P. Analytical Gas Chromatography, 2nd ed.; Academic Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Mihailova, A.; Kelly, S.D.; Chevallier, O.P.; Elliott, C.T.; Maestroni, B.M.; Cannavan, A. High-resolution mass spectrometry-based metabolomics for the discrimination between organic and conventional crops: A review. Trends Food Sci. Technol. 2021, 110, 142–154. [Google Scholar] [CrossRef]
- Tan, K.; Ipcho, S.V.S.; Trengove, R.D.; Oliver, R.P.; Solomon, P.S. Challenges for molecular plant pathology over the next ten years: Assessing the impact of transcriptomics, proteomics and metabolomics on fungal phytopathology. Mol. Plant Pathol. 2009, 10, 703–715. [Google Scholar] [CrossRef]
- Beleggia, R.; Platani, C.; Papa, R.; Di Chio, A.; Barros, E.; Mashaba, C.; Wirth, J.; Fammartino, A.; Sautter, C.; Conner, S.; et al. Metabolomics and food processing: From semolina to pasta. J. Agric. Food Chem. 2011, 59, 9366–9377. [Google Scholar] [CrossRef]
- Palamareva, M.D. Liquid Chromatography: Overview. In Encyclopedia of Analytical Science, 2nd ed.; Worsfold, P., Townshend, A., Poole, C., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2005; pp. 102–106. [Google Scholar]
- Swartz, M. Liquid chromatography: Mobile phase selection. In Encyclopedia of Analytical Science, 2nd ed.; Worsfold, P., Townshend, A., Poole, C., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2005; pp. 135–142. [Google Scholar]
- Meyer, V.R. Practical High-Performance Liquid Chromatography, 5th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010. [Google Scholar]
- Cubero-Leon, E.; De Rudder, O.; Maquet, A. Metabolomics for organic food authentication: Results from a long-term field study in carrots. Food Chem. 2018, 239, 760–770. [Google Scholar] [CrossRef]
- do Lago, C.L.; Daniel, D.; Lopes, F.S.; Cieslarová, Z. Electrophoresis. In Chemical Analysis of Food—Techniques and Applications; Yolanda, P., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 499–522. [Google Scholar]
- Stringer, R. Electrophoresis: Overview. In Encyclopedia of Analytical Science, 2nd ed.; Worsfold, P., Townshend, A., Poole, C., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2005; pp. 356–363. [Google Scholar]
- Bowser, M. Capillary Electrophoresis: Overview. In Encyclopedia of Analytical Science, 2nd ed.; Worsfold, P., Townshend, A., Poole, C., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2005; pp. 334–343. [Google Scholar]
- Snyder, L.R.; Kirkland, J.J.; Dolan, J.W. Introduction to Modern Liquid Chromatography. J. Am. Soc. Mass Spectrom. 2010, 22, 196. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, C.; Simó, C.; García-Cañas, V.; Cifuentes, A.; Castro-Puyana, M. Metabolomics, peptidomics and proteomics applications of capillary electrophoresis-mass spectrometry in Foodomics: A review. Anal. Chim. Acta 2013, 802, 1–13. [Google Scholar] [CrossRef]
- García-Villalba, R.; León, C.; Dinelli, G.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Garcia-Cañas, V.; Cifuentes, A. Comparative metabolomic study of transgenic versus conventional soybean using capillary electrophoresis-time-of-flight mass spectrometry. J. Chromatogr. A 2008, 1195, 164–173. [Google Scholar] [CrossRef]
- Levandi, T.; Leon, C.; Kaljurand, M.; Garcia-Cañas, V.; Cifuentes, A. Capillary electrophoresis time-of-flight mass spectrometry for comparative metabolomics of transgenic versus conventional maize. Anal. Chem. 2008, 80, 6329–6335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Quinn, K.; Cruickshank-Quinn, C.; Reisdorph, R.; Reisdorph, N. The application of ion mobility mass spectrometry to metabolomics. Curr. Opin. Chem. Biol. 2018, 42, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Feuerstein, M.L.; Kurulugama, R.T.; Hann, S.; Causon, T. Novel acquisition strategies for metabolomics using drift tube ion mobility-quadrupole resolved all ions time-of-flight mass spectrometry (IM-QRAI-TOFMS). Anal. Chim. Acta 2021, 1163, 338508. [Google Scholar] [CrossRef]
- Wishart, D.S. Perspectives in Magnetic Resonance NMR metabolomics: A look ahead. J. Magn. Reson. 2019, 306, 155–161. [Google Scholar] [CrossRef]
- Hatzakis, E. Nuclear Magnetic Resonance (NMR) Spectroscopy in Food Science: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 189–220. [Google Scholar] [CrossRef] [Green Version]
- Janovick, J.; Spyros, A.; Dais, P.; Hatzakis, E. Nuclear magnetic resonance. In Chemical Analysis of Food—Techniques and Applications; Pico, Y., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 135–175. [Google Scholar]
- Emwas, A.; Roy, R.; Mckay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.A.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR Spectroscopy for Metabolomics Research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Laghi, L.; Picone, G.; Capozzi, F. Nuclear magnetic resonance for foodomics beyond food analysis. Trends Anal. Chem. 2014, 59, 93–102. [Google Scholar] [CrossRef]
- Kaklamanos, G.; Aprea, E.; Theodoridis, G. Mass spectrometry: Principles and instrumentation. In Chemical Analysis of Food—Techniques and Applications, 2nd ed.; Pico, Y., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 525–552. [Google Scholar]
- Dunn, W.B. Current trends and future requirements for the mass spectrometric investigation of microbial, mammalian and plant metabolomes. Phys. Biol. 2008, 5, 011001. [Google Scholar] [CrossRef]
- Lee, D.K.; Yoon, M.H.; Kang, Y.P.; Yu, J.; Park, J.H.; Lee, J.; Kwon, S.W. Comparison of primary and secondary metabolites for suitability to discriminate the origins of Schisandra chinensis by GC/MS and LC/MS. Food Chem. 2013, 141, 3931–3937. [Google Scholar] [CrossRef]
- Okada, T.; Mochamad Afendi, F.; Altaf-Ul-Amin, M.; Takahashi, H.; Nakamura, K.; Kanaya, S. Metabolomics of Medicinal Plants: The Importance of Multivariate Analysis of Analytical Chemistry Data. Curr. Comput. Aided Drug Des. 2010, 6, 179–196. [Google Scholar] [CrossRef]
- Society, M. Metabolomic Database. 2014. Available online: http://metabolomicssociety.org/resources/metabolomics-databases (accessed on 22 July 2021).
- Reisdorph, N.; Reisdorph, R.; Quinn, K.; Doenges, K. Metabolomics Mass Spectrometry Data Processing: Applications in Food Analysis. In Comprehensive Foodomics; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Reid, L.M.; O’Donnell, C.P.; Downey, G. Potential of SPME-GC and Chemometrics to Detect Adulteration of Soft Fruit Purées. J. Agric. Food Chem. 2004, 52, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Pramai, P.; Abdul Hamid, N.A.; Mediani, A.; Maulidiani, M.; Abas, F.; Jiamyangyuen, S. Metabolite profiling, antioxidant, and α-glucosidase inhibitory activities of germinated rice: Nuclear-magnetic-resonance-based metabolomics study. J. Food Drug Anal. 2018, 26, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, F.; Wu, B. Use of Lipidomics for Food Quality Assurance and Authentication. In Comprehensive Foodomics; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar] [CrossRef]
- European Parliament; Council of the European Union. Regulation (EU) No 1169/2011 of the European Parliament and of the Council of 25 October 2011 on the provision of food information to consumers, amending Regulations (EC) No 1924/2006 and (EC) No 1925/2006 of the European Parliament and of the Council. Off. J. Eur. Union 2011, L304, 18–63. [Google Scholar]
- European Comission. The EU Food Fraud Network. Food Safety; European Comisison: Brussels, Belgium, 2021; Available online: https://ec.europa.eu/food/food/agri-food-fraud/eu-food-fraud-.network_en#meeting_reports (accessed on 5 September 2021).
- European Comission. Quality Schemes Explained; European Comission: Brussels, Belgium, 2021; Available online: https://ec.europa.eu/info/food-farming-fisheries/food-safety-and-quality/certification/quality-labels/quality-schemes-explained_en#aims (accessed on 5 September 2021).
- FAO/WHO. Nutrition and Labelling. In Codex Alimentarius International Food Standards; FAO: Rome, Italy, 2021; Available online: http://www.fao.org/fao-who-codexalimentarius/thematic-areas/nutrition-labelling/en/#c452837 (accessed on 5 September 2021).
- Cavanna, D.; Catellani, D.; Dall’asta, C.; Suman, M. Egg product freshness evaluation: A metabolomic approach. J. Mass Spectrom. 2018, 53, 849–861. [Google Scholar] [CrossRef] [Green Version]
- Solanky, K.S.; Bailey, N.J.C.; Beckwith-Hall, B.M.; Davis, A.; Bingham, S.; Holmes, E.; Nicholson, J.K.; Cassidy, A. Application of biofluid 1H nuclear magnetic resonance-based metabonomic techniques for the analysis of the biochemical effects of dietary isoflavones on human plasma profile. Anal. Biochem. 2003, 323, 197–204. [Google Scholar] [CrossRef]
- Wang, H.; Hua, J.; Yu, Q.; Li, J.; Wang, J.; Deng, Y.; Yuan, H. Widely targeted metabolomic analysis reveals dynamic changes in non-volatile and volatile metabolites during green tea processing. Food Chem. 2021, 363, 130131. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, T.; Kanaya, S.; Yonetani, T.; Kobayashi, A.; Fukusaki, E. Prediction of Japanese green tea ranking by fourier transform near-infrared reflectance spectroscopy. J. Agric. Food Chem. 2007, 55, 9908–9912. [Google Scholar] [CrossRef]
- Erban, A.; Fehrle, I.; Martinez-Seidel, F.; Brigante, F.; Más, A.L.; Baroni, V.; Wunderlin, D.; Kopka, J. Discovery of food identity markers by metabolomics and machine learning technology. Sci. Rep. 2019, 9, 9697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rešetar, D.; Pavelić, S.K.; Josić, D. Foodomics for investigations of food toxins. Curr. Opin. Food Sci. 2015, 4, 86–91. [Google Scholar] [CrossRef]
- Giacometti, J.; Josic, D. Foodomics in microbial safety. Trends Anal. Chem. 2013, 52, 16–22. [Google Scholar] [CrossRef]
- Ulaszewska, M.M.; Weinert, C.H.; Trimigno, A.; Portmann, R.; Andres Lacueva, C.; Badertscher, R.; Brennan, L.; Brunius, C.; Bub, A.; Capozzi, F.; et al. Nutrimetabolomics: An Integrative Action for Metabolomic Analyses in Human Nutritional Studies. Mol. Nutr. Food Res. 2019, 63, e1800384. [Google Scholar] [CrossRef]
- Wishart, D.S. Metabolomics: Applications to food science and nutrition research. Trends Food Sci. Technol. 2008, 19, 482–493. [Google Scholar] [CrossRef]
- Gibney, M.J.; Walsh, M.; Brennan, L.; Roche, H.M.; German, B.; Van Ommen, B. Metabolomics in human nutrition: Opportunities and challenges. Am. J. Clin. Nutr. 2005, 82, 497–503. [Google Scholar] [CrossRef]
- Alfaro, A.C.; Young, T. Showcasing metabolomic applications in aquaculture: A review. Rev. Aquac. 2018, 10, 135–152. [Google Scholar] [CrossRef]
- Utpott, M.; Rodrigues, E.; de Rios, A.O.; Mercali, G.D.; Flôres, S.H. Metabolomics: An analytical technique for food processing evaluation. Food Chem. 2022, 366, 130685. [Google Scholar] [CrossRef] [PubMed]
- Lucini, L.; Rocchetti, G.; Kane, D.; Trevisan, M. Phenolic fingerprint allows discriminating processed tomato products and tracing different processing sites. Food Control 2017, 73, 696–703. [Google Scholar] [CrossRef]
- Rubert, J.; Zachariasova, M.; Hajslova, J. Advances in high-resolution mass spectrometry based on metabolomics studies for food—A review. Food Addit. Contam. Part A Chem. Anal. Control. Expo Risk Assess. 2015, 32, 1685–1708. [Google Scholar] [CrossRef]
- Lytou, A.E.; Panagou, E.Z.; Nychas, G.J.E. Effect of different marinating conditions on the evolution of spoilage microbiota and metabolomic profile of chicken breast fillets. Food Microbiol. 2017, 66, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ye, Y.; Pan, D.; Sun, Y.; Wang, Y.; Cao, J. Metabonomics profiling of marinated meat in soy sauce during processing. J. Sci. Food Agric. 2018, 98, 1325–1331. [Google Scholar] [CrossRef]
- Garcia, C.J.; García-Villalba, R.; Garrido, Y.; Gil, M.I.; Tomás-Barberán, F.A. Untargeted metabolomics approach using UPLC-ESI-QTOF-MS to explore the metabolome of fresh-cut iceberg lettuce. Metabolomics 2016, 12, 138. [Google Scholar] [CrossRef]
- Hennig, K.; De Vos, R.C.H.; Maliepaard, C.; Dekker, M.; Verkerk, R.; Bonnema, G. A metabolomics approach to identify factors influencing glucosinolate thermal degradation rates in Brassica vegetables. Food Chem. 2014, 155, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Arapitsas, P.; Della Corte, A.; Gika, H.; Narduzzi, L.; Mattivi, F.; Theodoridis, G. Studying the effect of storage conditions on the metabolite content of red wine using HILIC LC-MS based metabolomics. Food Chem. 2016, 197, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Teegarden, M.D.; Schwartz, S.J.; Cooperstone, J.L. Profiling the impact of thermal processing on black raspberry phytochemicals using untargeted metabolomics. Food Chem. 2019, 274, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Tu, Y.; Huang, D.; Dong, Z.; Huang, M.; Cheng, J.; Tan, J.; Chen, W.; Sun, L.; Chen, W. Selection of appropriate post-harvest processing methods based on the metabolomics analysis of Salvia miltiorrhiza Bunge. Food Res. Int. 2021, 144, 110366. [Google Scholar] [CrossRef]
- Chen, Q.; Shi, J.; Mu, B.; Chen, Z.; Dai, W.; Lin, Z. Metabolomics combined with proteomics provides a novel interpretation of the changes in nonvolatile compounds during white tea processing. Food Chem. 2020, 332, 127412. [Google Scholar] [CrossRef]



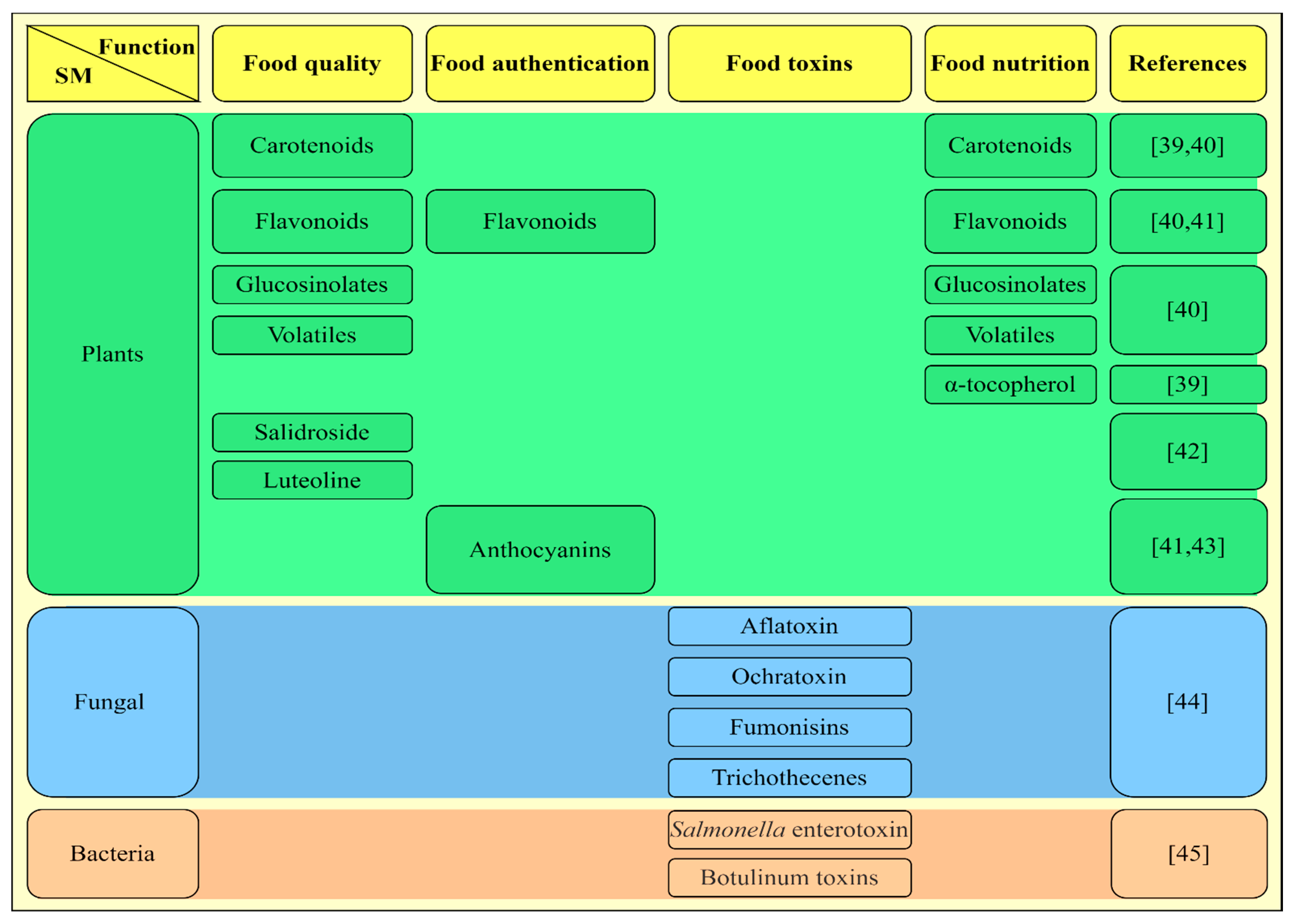{kind=link}
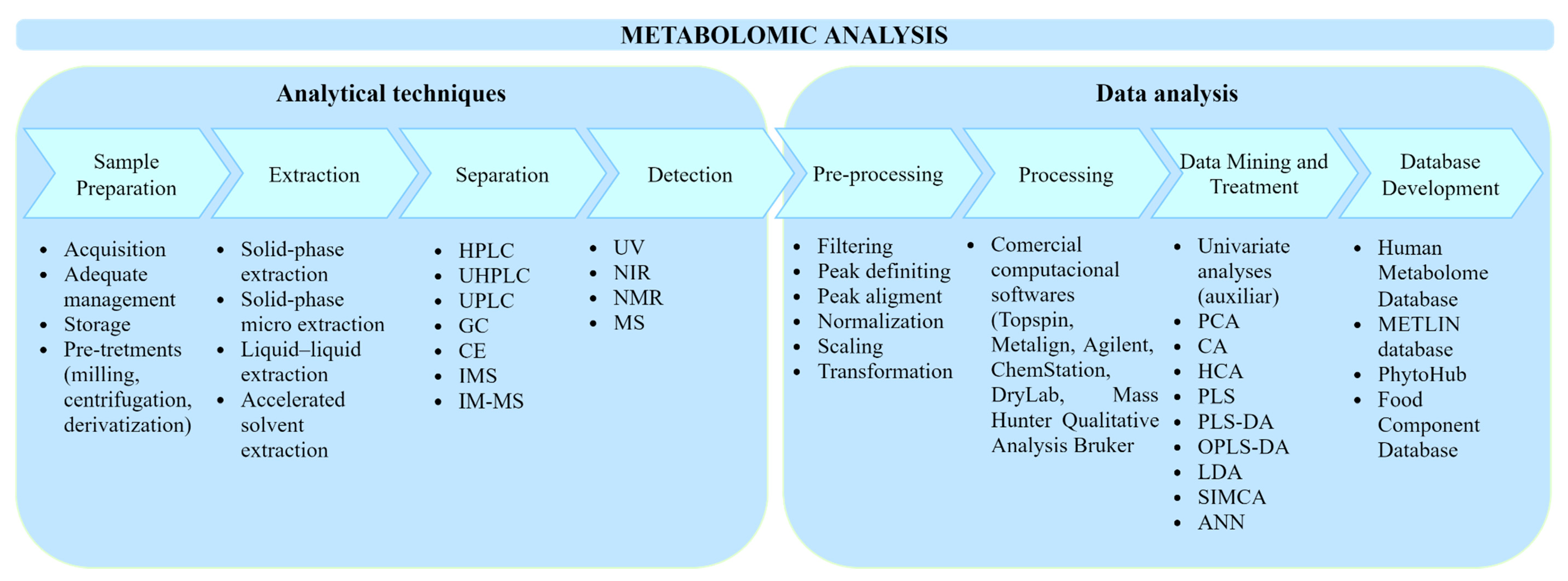{kind=link}
| Food Purpose of Analysis | Detection Technology | Data Treatment | References | |
|---|---|---|---|---|
| Authentication | Milk compounds (sugars, vitamins, nucleotides, and aromatic compounds) to distinguish milk from pasture and indoor total mixed ration-based diets | 1H NMR | PLS-DA | [12] |
| Authentication | Variation of coffee components by region | LC–MS and GC-FID; targeted and non-targeted analysis | PCA | [58] |
| Authentication | Identification of anthocyanin content of red wines to detect possible adulteration with black rice anthocyanins | 1H NMR and Fourier transform near-infrared | [43] | |
| Authentication | Identification of anthocyanins in Pinot Noir, Cabernet Sauvignon, and Merlot red wines | NMR and LC–MS | ||
| Authentication | Anthocyanins profile of grape berry skins belonging to different grape varieties | HPLC–MS and HPLC–NMR | ||
| Authentication | Analysis of the aromatic composition of wine phenolic extracts | LC–NMR/MS | ||
| Authentication | Differentiation of fiano di Avellino white wines obtained by fermentation with either a commercial or a selected autochthonous Saccharomyces cerevisiae yeast starter | 1H NMR | PCA | |
| Authentication | Metabolite profiling to study the fermentative behavior of lactic acid bacteria in grape wines | 1H NMR and GC | ||
| Authentication | Negroamaro red wines obtained through different wine-making technologies (traditional, ultrasounds, and cryomaceration with dry ice) and soil management practices (soil tillage and cover crop) | 1H NMR and GC | PCA (unsupervised) and OPLS-DA (supervised) | |
| Authentication | Prediction of the origin of the agricultural system through metabolite profile of carrots (Daucus carota L.) | LC–MS; untargeted | OPLS-DA | [78] |
| Authentication | Determination of Schisandra chinensis herb origin through GC–MS and LC–MS, and primary or secondary metabolites | GC–MS and LC–MS | PCA | [95] |
| Quality | Identification of metabolomic marker compounds to predict lettuce (Lactuca sativa L.) browning | UHPLC–HRMS; untargeted analysis | PCA; SIMCA 13 | [52] |
| Quality | Identification of metabolites in mangosteen (Garcinia mangostana Linn.) that contribute to ripening characteristics | GC–MS and LC–MS | PCA and PLS-DA | [61] |
| Quality | Influence of grapevine red blotch disease on the primary and secondary metabolites in skin, pulp, and seed tissues of Cabernet sauvignon grapes at harvest | 1H NMR and RP-HPLC–DAD | PCA, analysis of variance and the two means of each variable, t-test | [53] |
| Quality | Identification of biomarkers compounds responsible for freshness and non-freshness of egg products and validation of molecules | (UHPLC–HRMS); untargeted | SIMCA, PCA, and OPLS-DA | [106] |
| Quality/Nutritional | Investigation of metabolite profile variations during industrial pasta processing for five different commercial products | GC–MS and LC–MS | ANOVA, PCA, and factor analysis | [74] |
| Nutritional | Metabolite profile and discrimination among the different germinated rice (black, red, and white) | 1H NMR | PCA and PLS-DA | [100] |
| Nutritional | Investigation of metabolic changes following dietary intervention with soy isoflavones in healthy of premenopausal women | 1H NMR and RP-HPLC–DAD | PCA and SIMCA-P | [107] |
| Application in Food Processing | Detection Technology | Data Treatment | References |
|---|---|---|---|
| Variation in phytochemical profiles (carotenoids, flavonoids, glucosinolates, volatiles) of tomato, broccoli, and carrot purees modifying the processing order (between blending and heat treatment) | HPLC-PDA, GC–MS, 1H NMR, RP-LC-PDA-QTOF MS, and GC–MS for volatiles components | PCA and Student’s t-tests | [40] |
| Investigation of metabolomic profiling of chia, linseed, and sesame as processing-dependent biomarkers in cookies production | GC–MS | PCA and RF | [110] |
| Investigation of metabolite profile variations (phytos-terols, hydroxy fatty acids, tocopherols, and carotenoids) during industrial semolina pasta processing for five different commercial products | GC–MS and LC–MS | ANOVA, PCA | [74] |
| Investigation of different marinades in chicken breast fillets. Combination between pomegranate and lemon juice, probably due to the synergistic effect of organic acids (lemon juice) and polyphenols (pomegranate juice), provided the high decrease in Pseudomonas spp. bacteria | HPLC system | PCA | [120] |
| Investigation of changes in metabolite composition of marinated meat in soy sauce during processing as taste quality is directly related to primary and secondary metabolites | 1H NMR | PCA, OPLS-DA, and ANOVA | [121] |
| Study on metabolomics of lettuce and the changes after storage of two cultivars with different susceptibility to browning. Tendency showed high amounts of phenolic compounds, fatty acids, and lysophospholipid with the storage time (day 5) and with the browning process | UPLC–ESI-QTOF-MS (untargeted) | PCA and HCA (unsupervised methods) | [122] |
| Investigation of the relationship between specific metabolites and the plant matrix with glucosinolate thermal degradation during food processing of Brassica vegetables. The interest is to minimize losses of glucosinolate during vegetable processing | HPLC-PDA-QTOF MS (untargeted) | PCA, HCA and RF | [123] |
| Optimization by applying metabolic profiling method to study the effect of typical domestic storage conditions for five red wines for a period of 24 months. Storage conditions had a major impact on the polar metabolite fingerprint, and the markers revealed included phenolic compounds, vitamins, and 4-amino-heptanedioic acid and its ethyl ester | UPLC–QTOF-MS (untargeted) | PCA, OPLS-DA, t-test, U-test, and S-plot | [124] |
| Investigation of distinctions in the phenolic profile of tomato products (crushed pulp, puree, and paste) and in tomato paste under three different treatments (cold, warm, and hot). Distinctions were possible to identify, especially in relation to flavonoids, phenylpropanoids, and lignans, as well as distinctions between the production location | UHPLC/Q-TOF | ANOVA, HCA, (unsupervised) and PLS-DA | [118] |
| Investigation of chemical profile changes resulting from thermal processing of black raspberries powder into a nectar beverage with a metabolomics approach. Degradation products of anthocyanins were identified along with other proposed phenolic degradants, while quercetin, phenolic acids, and ellagic acid were relatively stable to processing | UHPLC–QTOF-MS (Untargeted) | HCA | [125] |
| Investigation of metabolic changes during post-harvest of Salvia miltiorrhiza Bunge. The processing demonstrated great impacts on phenolic acids than on tanshinones, and enzymatic browning was the major influencing factor during post-harvest processing. The data showed that the reduction of the enzymatic browning could be achieved by controlling the moisture and steaming process | UHPLC–QTOF-MS | PCA (unsupervised), PLS-DA, and OPLS-DA | [126] |
| Investigation of metabolomics and proteomics to study the change mechanism of nonvolatile compounds during white tea processing. Decreased content of catechins, proanthocyanidins, thasins, and phenolic acids and increased content of free amino acids, theaflavins, and nucleotides are responsible for the sweet taste of tea. The drying process was found to promote the formation of white tea taste | UPLC–LTQ-Orbitrap-MS (untargeted) | PCA and ANOVA | [127] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pedrosa, M.C.; Lima, L.; Heleno, S.; Carocho, M.; Ferreira, I.C.F.R.; Barros, L. Food Metabolites as Tools for Authentication, Processing, and Nutritive Value Assessment. Foods 2021, 10, 2213. https://doi.org/10.3390/foods10092213
Pedrosa MC, Lima L, Heleno S, Carocho M, Ferreira ICFR, Barros L. Food Metabolites as Tools for Authentication, Processing, and Nutritive Value Assessment. Foods. 2021; 10(9):2213. https://doi.org/10.3390/foods10092213
Chicago/Turabian StylePedrosa, Mariana C., Laíres Lima, Sandrina Heleno, Márcio Carocho, Isabel C. F. R. Ferreira, and Lillian Barros. 2021. "Food Metabolites as Tools for Authentication, Processing, and Nutritive Value Assessment" Foods 10, no. 9: 2213. https://doi.org/10.3390/foods10092213